Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Μεταλλάξεις και αναδιατάξεις του γονιδιωματικού DNA
Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

16
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.17 Αυθόρμητες βλάβες του DNA. Υπάρχουν δύο βασικοί τύποι αυθόρμητων βλαβών του DNA: (Α) η απαμίνωση της αδενίνης, της κυτοσίνης και της γουανίνης και (Β) η αποπουρίνωση (απομάκρυνση πουρινών) λόγω της διάσπασης του δεσμού μεταξύ μιας πουρίνης και της δεοξυριβόζης, οπότε προκύπτει στο DNA μια απουρινική θέση (AP, Apurinic). dGMP = μονοφωσφορική δεοξυγουανοσίνη (deoxyguanosine Μonophosphate). Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
 η απαμίνωση της αδενίνης, της κυτοσίνης και της γουανίνης και (Β) η αποπουρίνωση (απομάκρυνση πουρινών) λόγω της διάσπασης του δεσμού μεταξύ μιας πουρίνης και της δεοξυριβόζης, οπότε προκύπτει στο DNA μια απουρινική θέση (AP, Apurinic). dGMP = μονοφωσφορική δεοξυγουανοσίνη (deoxyguanosine Μonophosphate). Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
17
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.18 Παραδείγματα βλάβης του DNA που επάγεται από ακτινοβολία και χημικές ουσίες. (Α) Το υπεριώδες φως επάγει τον σχηματισμό διμερών πυριμιδίνης στα οποία δύο παρακείμενες πυριμιδίνες (π.χ. θυμίνες) συνδέονται με έναν κυκλοβουτανικό δακτύλιο. (Β) Αλκυλίωση είναι η προσθήκη μεθυλομάδων ή αιθυλομάδων σε ποικίλες θέσεις των βάσεων του DNA. Στο παράδειγμα που παρουσιάζεται εδώ, η αλκυλίωση της O6 θέσης της γουανίνης οδηγεί στον σχηματισμό O6-μεθυλογουανίνης. (Γ) Πολλά καρκινογόνα [π.χ. το βενζο(α)πυρένιο] αντιδρούν με βάσεις του DNA, προκαλώντας την προσθήκη μεγάλων χημικών ομάδων. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
 Το υπεριώδες φως επάγει τον σχηματισμό διμερών πυριμιδίνης στα οποία δύο παρακείμενες πυριμιδίνες (π.χ. θυμίνες) συνδέονται με έναν κυκλοβουτανικό δακτύλιο. (Β) Αλκυλίωση είναι η προσθήκη μεθυλομάδων ή αιθυλομάδων σε ποικίλες θέσεις των βάσεων του DNA. Στο παράδειγμα που παρουσιάζεται εδώ, η αλκυλίωση της O6 θέσης της γουανίνης οδηγεί στον σχηματισμό O6-μεθυλογουανίνης. (Γ) Πολλά καρκινογόνα [π.χ. το βενζο(α)πυρένιο] αντιδρούν με βάσεις του DNA, προκαλώντας την προσθήκη μεγάλων χημικών ομάδων. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
19
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.19 Άμεση επιδιόρθωση διμερών θυμίνης. Τα διμερή θυμίνης που έχουν προκληθεί από υπεριώδη ακτινοβολία είναι δυνατόν να επιδιορθωθούν με φωτοεπανενεργοποίηση, κατά την οποία χρησιμοποιείται ενέργεια από το ορατό φως για να διασπαστούν οι δεσμοί που σχηματίζουν τον κυκλοβουτανικό δακτύλιο. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

20
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.20 Επιδιόρθωση της O6-μεθυλογουανίνης. Το ένζυμο μεθυλοτρανσφεράση της O6-μεθυλογουανίνης μεταφέρει τη μεθυλομάδα από την O6-μεθυλογουανίνη σε μια κυστεΐνη του ενεργού του κέντρου. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

22
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.21 Επιδιόρθωση με εκτομή βάσης. Σε αυτό το παράδειγμα έχει σχηματιστεί ουρακίλη (U) από την απαμίνωση κυτοσίνης (C), με αποτέλεσμα να υπάρχει μια ουρακίλη απέναντι από μια γουανίνη (G) τηςσυμπληρωματικής αλυσίδας του DNA. Ο δεσμός μεταξύ της ουρακίλης και της δεοξυριβόζης κόβεται από μια DNA γλυκοζυλάση, αφήνοντας στο DNA ένα σάκχαρο που δε συνδέεται με βάση (μια θέση AP). Η θέση αυτή εντοπίζεται από την ενδονουκλεάση AP, η οποία κόβει την αλυσίδα του DNA. Η δεοξυριβόζη απομακρύνεται από τη φωσφοδιεστεράση της δεοξυριβόζης (deoxyribose phosphodiesterase). Το κενό που δημιουργείται αποκαθίσταται αρχικά με τη δράση της DNA πολυμεράσης που προσθέτει τη σωστή βάση (C) και κατόπιν με τη δράση της λιγάσης. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
 από την απαμίνωση κυτοσίνης (C), με αποτέλεσμα να υπάρχει μια ουρακίλη απέναντι από μια γουανίνη (G) τηςσυμπληρωματικής αλυσίδας του DNA. Ο δεσμός μεταξύ της ουρακίλης και της δεοξυριβόζης κόβεται από μια DNA γλυκοζυλάση, αφήνοντας στο DNA ένα σάκχαρο που δε συνδέεται με βάση (μια θέση AP). Η θέση αυτή εντοπίζεται από την ενδονουκλεάση AP, η οποία κόβει την αλυσίδα του DNA. Η δεοξυριβόζη απομακρύνεται από τη φωσφοδιεστεράση της δεοξυριβόζης (deoxyribose phosphodiesterase). Το κενό που δημιουργείται αποκαθίσταται αρχικά με τη δράση της DNA πολυμεράσης που προσθέτει τη σωστή βάση (C) και κατόπιν με τη δράση της λιγάσης. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
24
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
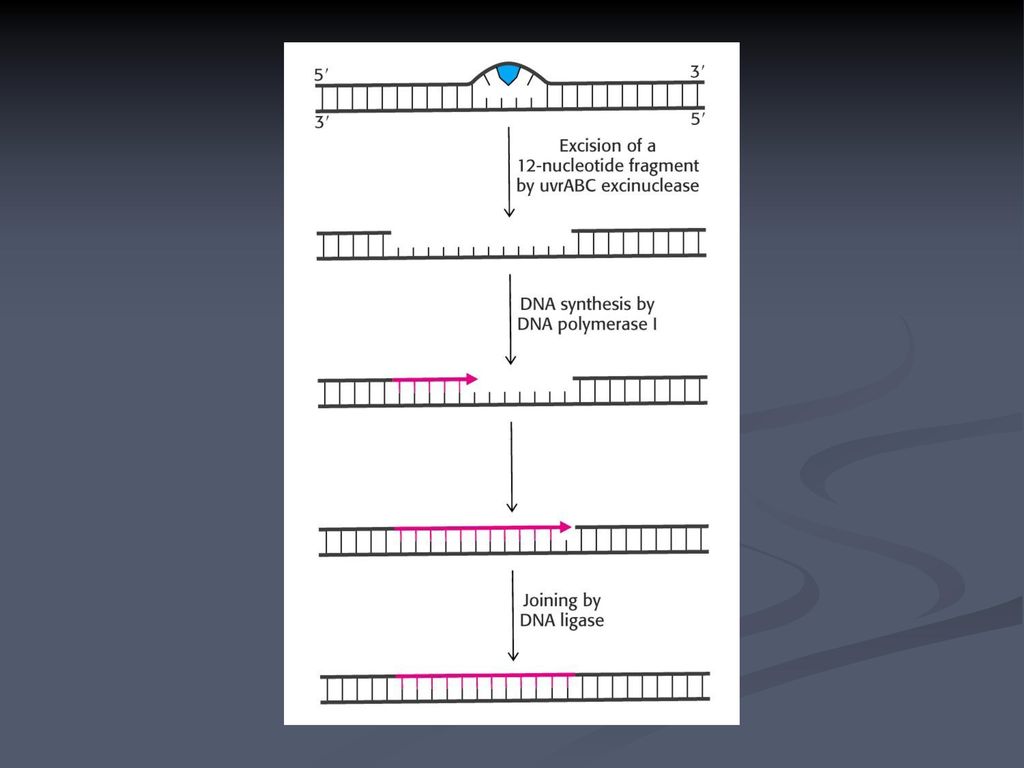
ΕΙΚΟΝΑ 6.22 Επιδιόρθωση διμερών θυμίνης με εκτομή νουκλεοτιδίου. Το DNA που έχει υποστεί τη βλάβη εντοπίζεται και κόβεται και στις δύο πλευρές του διμερούς θυμίνης από νουκλεάσες. Ακολουθεί η αποπεριέλιξή του από μια ελικάση, ώστε να επιτευχθεί η εκτομή ενός ολιγονουκλεοτιδίου που περιέχει τις βάσεις που έχουν υποστεί βλάβη. Το χάσμα που δημιουργείται αποκαθίσταται από την DNA πολυμεράση και τη λιγάση. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

25
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.23 Επιδιόρθωση με εκτομή νουκλεοτιδίου σε κύτταρα θηλαστικών. Η βλάβη στο DNA (π.χ. ένα διμερές θυμίνης) εντοπίζεται από την XPC και ακολουθεί η πρόσδεση της XPA, του RPA και του TFIIH, του οποίου υπομονάδες είναι οι ελικάσες XPB και XPD. Μετά την αποπεριέλιξη του DNA από τις XPB και XPD, στρατολογούνται οι ενδονουκλεάσες XPG και XPF/ERCC1, οι οποίες κόβουν το DNA και αφαιρούν το ολιγονουκλεοτίδιο που φέρει τη βλάβη. Το χάσμα που δημιουργείται αποκαθίσταται από την DNA πολυμεράση και τη λιγάση. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
 εντοπίζεται από την XPC και ακολουθεί η πρόσδεση της XPA, του RPA και του TFIIH, του οποίου υπομονάδες είναι οι ελικάσες XPB και XPD. Μετά την αποπεριέλιξη του DNA από τις XPB και XPD, στρατολογούνται οι ενδονουκλεάσες XPG και XPF/ERCC1, οι οποίες κόβουν το DNA και αφαιρούν το ολιγονουκλεοτίδιο που φέρει τη βλάβη. Το χάσμα που δημιουργείται αποκαθίσταται από την DNA πολυμεράση και τη λιγάση. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
26
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.24 Επιδιόρθωση συνδεδεμένη με τη μεταγραφή σε κύτταρα θηλαστικών. Η RNA πολυμεράση λόγω της βλάβης (π.χ. ενός διμερούς θυμίνης) ακινητοποιείται στη μεταγραφόμενη αλυσίδα του DNA. Η σταματημένη πολυμεράση στρατολογεί τις CSA και CSB στη θέση της βλάβης. Στη συνέχεια, οι CSA και CSB στρατολογούν την XPA, την RPA και τον TFIIH. Κατόπιν, πραγματοποιείται η επιδιόρθωση με εκτομή, όπως περιγράφεται στην Εικόνα 6.23. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
 ακινητοποιείται στη μεταγραφόμενη αλυσίδα του DNA. Η σταματημένη πολυμεράση στρατολογεί τις CSA και CSB στη θέση της βλάβης. Στη συνέχεια, οι CSA και CSB στρατολογούν την XPA, την RPA και τον TFIIH. Κατόπιν, πραγματοποιείται η επιδιόρθωση με εκτομή, όπως περιγράφεται στην Εικόνα Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
28
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.25 Επιδιόρθωση αταίριαστου ζεύγους στην E. coli. Το σύστημα επιδιόρθωσης αταίριαστου ζεύγους ανιχνεύει και αποκόπτει αταίριαστες βάσεις στο νεοσυντιθέμενο DNA, το οποίο διακρίνεται από τη μητρική αλυσίδα επειδή δεν έχει ακόμα μεθυλιωθεί. Η MutS προσδένεται στην αταίριαστη βάση και ακολουθεί η πρόσδεση των MutL και MutH. Η MutH ενεργοποιείται από τη MutL και κόβει τη μη μεθυλιωμένη αλυσίδα απέναντι από μια θέση μεθυλίωσης. Στη συνέχεια, οι MutS και MutL, μαζί με μια ελικάση και μια εξωνουκλεάση, αποκόπτουν το τμήμα της μη μεθυλιωμένης αλυσίδας που περιέχει την αταίριαστη βάση. Το χάσμα που δημιουργείται αποκαθίσταται από την DNA πολυμεράση και τη λιγάση. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

29
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.26 Επιδιόρθωση αταίριαστου ζεύγους σε κύτταρα θηλαστικών. Οι ευκαρυωτικές πρωτεΐνες MSH και MLH, που είναι ομόλογες των βακτηριακών MutS και MutL αντίστοιχα, προσδένονται σε μια αταίριαστη βάση και κατευθύνουν την εκτομή του DNA μεταξύ αυτής και μιας μονόκλωνης ρήξης. Οι μονόκλωνες ρήξεις αντιστοιχούν στις εγκοπές στα άκρα των τμημάτων Okazaki του καθυστερημένου κλώνου και στο αναπτυσσόμενο 3΄ άκρο του προπορευόμενου κλώνου. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

30
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.27 Διαβλαβική σύνθεση DNA. Ένα διμερές θυμίνης παρεμποδίζει την αντιγραφή. Ωστόσο, εξειδικευμένες DNA πολυμεράσες, όπως η πολυμεράση V (pol V), εντοπίζουν τη βλάβη και διεκπεραιώνουν τη σύνθεση του DNA απέναντι από την περιοχή της βλάβης. Στη συνέχεια, η σύνθεση του DNA συνεχίζεται από την κύρια πολυμεράση της αντιγραφής, ενώ το διμερές θυμίνης απομακρύνεται με επιδιόρθωση με εκτομή νουκλεοτιδίου. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
, εντοπίζουν τη βλάβη και διεκπεραιώνουν τη σύνθεση του DNA απέναντι από την περιοχή της βλάβης. Στη συνέχεια, η σύνθεση του DNA συνεχίζεται από την κύρια πολυμεράση της αντιγραφής, ενώ το διμερές θυμίνης απομακρύνεται με επιδιόρθωση με εκτομή νουκλεοτιδίου. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
34
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.28 Επιδιόρθωση δίκλωνων ρήξεων. Η ιονίζουσα ακτινοβολία και ορισμένες χημικές ενώσεις προκαλούν δίκλωνες ρήξεις στο DNA. Αυτές οι βλάβες είναι δυνατόν να επιδιορθωθούν μέσω ομόλογου ανασυνδυασμού με ένα φυσιολογικό χρωμόσωμα. Εναλλακτικά, τα κομμένα άκρα του μορίου είναι δυνατόν απλώς να επανασυνδεθούν μεταξύ τους. Η διαδικασία όμως αυτή, που είναι γνωστή ως σύνδεση μη ομόλογων άκρων, οδηγεί συχνά στην απώλεια βάσεων γύρω από την περιοχή της βλάβης. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

36
Working model for joining of various types of DNA DSBs
Working model for joining of various types of DNA DSBs. First, DSB recognition by Ku70/80 is fast and reversible. Subsequently, XRCC4/ligase IV can be attracted, forming a reversible complex. Alternatively, DNA-PKCS and XRCC4/ligase IV can both be recruited and form a more stable complex. We hypothesize that the joining reaction of simple DSBs can be accomplished via either of these two intermediates, whereas complex DSBs can be joined only via the right branch.

41
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.29 Ομόλογος ανασυνδυασμός με ζευγάρωμα συμπληρωματικών βάσεων. Αρχικά, δημιουργούνται ταυτόσημες ασυμπτωτικές εγκοπές στα δύο μητρικά μόρια DNA. Κατόπιν, η μονόκλωνη περιοχή κάθε χρωμοσώματος ζευγαρώνει με τη συμπληρωματική της μονόκλωνη περιοχή του άλλου χρωμοσώματος. Έτσι δημιουργείται μια ετεροδίκλωνη περιοχή (δηλαδή μια περιοχή στην οποία οι δύο αλυσίδες του DNA προέρχονται από διαφορετικά μητρικά μόρια), ενώ παράλληλα ανταλλάσσονται τμήματα των εμπλεκόμενων μορίων. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
, ενώ παράλληλα ανταλλάσσονται τμήματα των εμπλεκόμενων μορίων. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
42
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.30 Το μοντέλο του Holliday για τον ομόλογο ανασυνδυασμό. Αρχικά, δημιουργείται μια μονόκλωνη εγκοπή στην ίδια θέση σε καθένα από τα μητρικά μόρια. Ακολούθως, τα δύο μόρια ανταλλάσσουν τις αλυσίδες με την εγκοπή. Στη συνέχεια, οι δύο εγκοπές κλείνουν με τη δράση της DNA λιγάσης και σχηματίζεται ένα ενδιάμεσο μόριο, ο κόμβος του Holliday, στο οποίο διασταυρώνονται οι δύο αλυσίδες που ανταλλάχθηκαν. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

43
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.31 Ταυτοποίηση των κόμβων του Holliday με ηλεκτρονική μικροσκοπία. Φωτογραφία ηλεκτρονικού μικροσκοπίου που δείχνει έναν κόμβο του Holliday κατά τον ανασυνδυασμό πλασμιδιακών μορίων DNA στην E. coli. Από κάτω παρουσιάζεται σχηματικά η δομή που φαίνεται στη φωτογραφία. Η διαμόρφωση του κόμβου που εμφανίζεται εδώ προκύπτει από το μόριο με τις διασταυρωμένες αλυσίδες μετά από περιστροφή του, όπως φαίνεται διαγραμματικά στην Εικόνα (Ευγενική προσφορά των Huntington Potter, University of South Florida, και David Dressler, University of Oxford.) Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
 Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
44
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.32 Iσομερισμός και λύση των κόμβων του Holliday. Οι κόμβοι του Holliday λύνονται με κοπή και επανασύνδεση των διασταυρωμένων αλυσίδων. Όταν διασταυρώνονται μεταξύ τους οι αλυσίδες που είχαν υποστεί τις εγκοπές στην αρχή της διαδικασίας του ανασυνδυασμού, προκύπτουν ετεροδίκλωνα μόρια τα οποία δεν εμφανίζουν ανασυνδυασμό των γενετικών δεικτών που βρίσκονται εκατέρωθεν της ετεροδίκλωνης περιοχής. Ωστόσο, με δύο περιστροφές παράγεται ένα ισομερές στο οποίο διασταυρώνονται οι μητρικές αλυσίδες που δεν υπέστησαν αρχικά εγκοπές. Στην περίπτωση αυτή, από την κοπή και επανασύνδεση των διασταυρωμένων αλυσίδων προκύπτουν ανασυνδυασμένα ετεροδίκλωνα μόρια. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

45
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.33 Έναρξη του ανασυνδυασμού από δίκλωνες ρήξεις. Οι δύο αλυσίδες του DNA στην περιοχή της δίκλωνης ρήξης αποικοδομούνται μερικώς με κατεύθυνση 5΄προς 3΄ από νουκλεάσες. Στη συνέχεια, τα μονόκλωνα άκρα εισβάλλουν στο ομόλογο μόριο και ζευγαρώνουν μεταξύ τους οι αλυσίδες των δύο μητρικών μορίων. Τα χάσματα αποκαθίστανται με τη δράση της DNA πολυμεράσης και οι εγκοπές κλείνουν με τη δράση της DNA λιγάσης. Έτσι σχηματίζεται ένα μόριο με δύο κόμβους του Holliday. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

46
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.34 Λειτουργία της πρωτεΐνης RecA. Η RecA αρχικά προσδένεται σε μονόκλωνο DNA και σχηματίζεται ένα ινίδιο πρωτεΐνης-DNA. Στη συνέχεια, προσδένεται και σε ένα δίκλωνο μόριο DNA, οπότε σχηματίζεται ένα σύμπλοκο των δύο μορίων DNA χωρίς αρχικά να ζευγαρώνουν οι αλυσίδες τους. Κατόπιν, οι συμπληρωματικές βάσεις των ομόλογων περιοχών των δύο μορίων ζευγαρώνουν και πραγματοποιείται η ανταλλαγή αλυσίδων, που οδηγεί στον σχηματισμό μιας ετεροδίκλωνης περιοχής. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

47
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.35 Ινίδια που σχηματίζονται από την πρόσδεση των πρωτεϊνών RecA και Rad51 στο DNA. Φωτογραφίες ηλεκτρονικού μικροσκοπίου που δείχνουν ινίδια τα οποία σχηματίζονται από την πρόσδεση της πρωτεΐνης RecA της E. coli και της πρωτεΐνης Rad51 του ανθρώπου στο DNA. (Από τη δημοσίευση του S. C. West, Nature Rev. Mol. Cell Biol. 4: 1. Ευγενική προσφορά των A. Stasiak και S. West.) Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
 Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
48
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.36 Μετατόπιση του σημείου διασταύρωσης των αλυσίδων και λύση των κόμβων του Holliday. Η RuvA εντοπίζει τον κόμβο του Holliday και στρατολογεί τη RuvB, μαζί με την οποία καταλύουν τη μετατόπιση της θέσης στην οποία διασταυρώνονται οι δύο αλυσίδες. Στη συνέχεια, η RuvC λύνει τους κόμβους του Holliday κόβοντας τις διασταυρωμένες αλυσίδες, τις οποίες κατόπιν συνδέει η λιγάση. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

49
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.37 Δομή μιας ανοσοσφαιρίνης. Οι ανοσοσφαιρίνες αποτελούνται από δύο βαριές και δύο ελαφριές αλυσίδες, οι οποίες συνδέονται με δισουλφιδικούς δεσμούς. Τόσο οι βαριές όσο και οι ελαφριές αλυσίδες αποτελούνται από μεταβλητές και σταθερές περιοχές. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

50
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.38 Αναδιάταξη των γονιδίων των ελαφριών αλυσίδων των ανοσοσφαιρινών. Τα γονίδια των ελαφριών αλυσίδων (εδώ απεικονίζονται τα γονίδια των ελαφριών αλυσίδων κ του ποντικιού) αποτελούνται από μια σταθερή περιοχή (C), μια συνδετική περιοχή (J) και μια μεταβλητή περιοχή (V). Στο DNA της γαμετικής σειράς υπάρχουν 150 περίπου διαφορετικές περιοχές V, οι οποίες βρίσκονται σε απόσταση περίπου 20 kb από τέσσερις εναλλακτικές περιοχές J και από μία μοναδική περιοχή C. Κατά την ανάπτυξη των λεμφοκυττάρων B, με ανασυνδυασμό ειδικής θέσης συνδέεται μία από τις περιοχές V με μία από τις περιοχές J. Αυτή η αναδιάταξη ενεργοποιεί τη μεταγραφή, προκαλώντας τον σχηματισμό ενός πρωτογενούς μεταγράφου το οποίο περιέχει την αναδιαταγμένη περιοχή VJ μαζί με τις υπόλοιπες περιοχές J και την περιοχή C. Στη συνέχεια, οι υπόλοιπες περιοχές J απομακρύνονται με μάτισμα, ώστε να παραχθεί ένα λειτουργικό mRNA με δομή VJC. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
 αποτελούνται από μια σταθερή περιοχή (C), μια συνδετική περιοχή (J) και μια μεταβλητή περιοχή (V). Στο DNA της γαμετικής σειράς υπάρχουν 150 περίπου διαφορετικές περιοχές V, οι οποίες βρίσκονται σε απόσταση περίπου 20 kb από τέσσερις εναλλακτικές περιοχές J και από μία μοναδική περιοχή C. Κατά την ανάπτυξη των λεμφοκυττάρων B, με ανασυνδυασμό ειδικής θέσης συνδέεται μία από τις περιοχές V με μία από τις περιοχές J. Αυτή η αναδιάταξη ενεργοποιεί τη μεταγραφή, προκαλώντας τον σχηματισμό ενός πρωτογενούς μεταγράφου το οποίο περιέχει την αναδιαταγμένη περιοχή VJ μαζί με τις υπόλοιπες περιοχές J και την περιοχή C. Στη συνέχεια, οι υπόλοιπες περιοχές J απομακρύνονται με μάτισμα, ώστε να παραχθεί ένα λειτουργικό mRNA με δομή VJC. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
51
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.39 Αναδιάταξη των γονιδίων των βαριών αλυσίδων των ανοσοσφαιρινών. Τα γονίδια των βαριών αλυσίδων εκτός από τις περιοχές V, J και C διαθέτουν και περιοχές D. Αρχικά, μια περιοχή D συνδέεται με μια περιοχή J και κατόπιν μια περιοχή V συνδέεται με την αναδιαταγμένη περιοχή DJ. Οι υπόλοιπες περιοχές J απομακρύνονται με μάτισμα, για να παραχθεί ένα λειτουργικό mRNA βαριάς αλυσίδας με δομή VDJC. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

52
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.40 Δομή ενός υποδοχέα κυττάρων T. Οι υποδοχείς των κυττάρων T αποτελούνται από τις πολυπεπτιδικές αλυσίδες α και β, που διαθέτουν μια μεταβλητή και μια σταθερή περιοχή. Οι δύο αυτές αλυσίδες διασχίζουν την κυτταροπλασματική μεμβράνη και συνδέονται μεταξύ τους με δισουλφιδικούς δεσμούς. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση

53
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.41 Ανασυνδυασμός V(D)J. Δίπλα στις κωδικές περιοχές των γονιδίων των ανοσοσφαιρινών και των υποδοχέων των κυττάρων T (για παράδειγμα, στις περιοχές V και D) βρίσκονται μικρές αλληλουχίες που ονομάζονται σήματα ανασυνδυασμού (RS, Recombination Signals). Τα δύο RS, τα οποία βρίσκονται εκατέρωθεν των τμημάτων DNA που πρόκειται να απομακρυνθούν κατά τον ανασυνδυασμό, έχουν αντίθετο προσανατολισμό το ένα σε σχέση με το άλλο (ο προσανατολισμός καθενός από τα δύο RS της εικόνας υποδεικνύεται με βέλος). Τα RS αναγνωρίζονται από ένα σύμπλοκο των πρωτεϊνών RAG1 και RAG2, οι οποίες εκφράζονται ειδικά στα λεμφοκύτταρα και κόβουν το DNA ανάμεσα στην κωδική αλληλουχία και το RS. Στη συνέχεια, οι δύο κωδικές περιοχές συνδέονται μεταξύ τους, προκειμένου να σχηματιστεί ένα αναδιαταγμένο γονιδιακό τμήμα. Εξαιτίας της απώλειας βάσεων κατά τη σύνδεση των μη ομόλογων άκρων και λόγω της προσθήκης βάσεων από την επάκρια δεοξυνουκλεοτιδυλική τρανσφεράση, προκαλούνται μεταλλαγές οι οποίες συνεισφέρουν στην αύξηση της ποικιλότητας των λειτουργικών γονιδίων που σχηματίζονται από την αναδιάταξη του γενετικού υλικού. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
J. Δίπλα στις κωδικές περιοχές των γονιδίων των ανοσοσφαιρινών και των υποδοχέων των κυττάρων T (για παράδειγμα, στις περιοχές V και D) βρίσκονται μικρές αλληλουχίες που ονομάζονται σήματα ανασυνδυασμού (RS, Recombination Signals). Τα δύο RS, τα οποία βρίσκονται εκατέρωθεν των τμημάτων DNA που πρόκειται να απομακρυνθούν κατά τον ανασυνδυασμό, έχουν αντίθετο προσανατολισμό το ένα σε σχέση με το άλλο (ο προσανατολισμός καθενός από τα δύο RS της εικόνας υποδεικνύεται με βέλος). Τα RS αναγνωρίζονται από ένα σύμπλοκο των πρωτεϊνών RAG1 και RAG2, οι οποίες εκφράζονται ειδικά στα λεμφοκύτταρα και κόβουν το DNA ανάμεσα στην κωδική αλληλουχία και το RS. Στη συνέχεια, οι δύο κωδικές περιοχές συνδέονται μεταξύ τους, προκειμένου να σχηματιστεί ένα αναδιαταγμένο γονιδιακό τμήμα. Εξαιτίας της απώλειας βάσεων κατά τη σύνδεση των μη ομόλογων άκρων και λόγω της προσθήκης βάσεων από την επάκρια δεοξυνουκλεοτιδυλική τρανσφεράση, προκαλούνται μεταλλαγές οι οποίες συνεισφέρουν στην αύξηση της ποικιλότητας των λειτουργικών γονιδίων που σχηματίζονται από την αναδιάταξη του γενετικού υλικού. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
54
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.42 Ανασυνδυασμός αλλαγής τάξης. Η αλλαγή τάξης συμβαίνει με ανασυνδυασμό ανάμεσα σε περιοχές-διακόπτες (S) οι οποίες χαρακτηρίζονται από την παρουσία υψηλά επαναλαμβανόμενων αλληλουχιών DNA και βρίσκονται ανοδικά κάθε περιοχής C στον γενετικό τόπο των βαριών αλυσίδων (εδώ απεικονίζεται ο γενετικός τόπος του ποντικιού). Στο παράδειγμα της εικόνας, ένα αναδιαταγμένο τμήμα V(D)J μετατοπίζεται από τη Cμ στη Cγ1 με ανασυνδυασμό μεταξύ των περιοχών-διακοπτών Sμ και Sγ1. Το παρεμβαλλόμενο DNA απομακρύνεται στη μορφή ενός κυκλικού μορίου. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
 οι οποίες χαρακτηρίζονται από την παρουσία υψηλά επαναλαμβανόμενων αλληλουχιών DNA και βρίσκονται ανοδικά κάθε περιοχής C στον γενετικό τόπο των βαριών αλυσίδων (εδώ απεικονίζεται ο γενετικός τόπος του ποντικιού). Στο παράδειγμα της εικόνας, ένα αναδιαταγμένο τμήμα V(D)J μετατοπίζεται από τη Cμ στη Cγ1 με ανασυνδυασμό μεταξύ των περιοχών-διακοπτών Sμ και Sγ1. Το παρεμβαλλόμενο DNA απομακρύνεται στη μορφή ενός κυκλικού μορίου. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
55
Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση
ΕΙΚΟΝΑ 6.43 Μοντέλο για τον ρόλο της επαγόμενης από ενεργοποίηση απαμινάσης (AID) στη σωματική υπερμεταλλαγή και στον ανασυνδυασμό αλλαγής τάξης. Η AID προκαλεί την απαμίνωση της κυτοσίνης του DNA σε ουρακίλη. Κατά την επακόλουθη απομάκρυνση της U με επιδιόρθωση με εκτομή βάσης (βλ. Εικόνα 6.21) δημιουργείται ένα κενό στη μία αλυσίδα του DNA. Η κάλυψη του κενού αυτού στις μεταβλητές (V) περιοχές, η οποία ενδεχομένως πραγματοποιείται από εξειδικευμένες DNA πολυμεράσες με μειωμένη πιστότητα (βλ. Εικόνα 6.27), μπορεί να οδηγήσει σε σωματική υπερμεταλλαγή. Στις περιοχές-διακόπτες (S), λόγω της εκτομής βάσεων σε αντικριστές περιοχές των δύο αλυσίδων, δημιουργούνται δίκλωνες ρήξεις που ενεργοποιούν τον μηχανισμό της σύνδεσης μη ομόλογων άκρων (βλ. Εικόνα 6.28), ο οποίος μπορεί να οδηγήσει σε ανασυνδυασμό αλλαγής τάξης. Ακαδημαϊκές Εκδόσεις Το κύτταρο-Μια Μοριακή Προσέγγιση
 στη σωματική υπερμεταλλαγή και στον ανασυνδυασμό αλλαγής τάξης. Η AID προκαλεί την απαμίνωση της κυτοσίνης του DNA σε ουρακίλη. Κατά την επακόλουθη απομάκρυνση της U με επιδιόρθωση με εκτομή βάσης (βλ. Εικόνα 6.21) δημιουργείται ένα κενό στη μία αλυσίδα του DNA. Η κάλυψη του κενού αυτού στις μεταβλητές (V) περιοχές, η οποία ενδεχομένως πραγματοποιείται από εξειδικευμένες DNA πολυμεράσες με μειωμένη πιστότητα (βλ. Εικόνα 6.27), μπορεί να οδηγήσει σε σωματική υπερμεταλλαγή. Στις περιοχές-διακόπτες (S), λόγω της εκτομής βάσεων σε αντικριστές περιοχές των δύο αλυσίδων, δημιουργούνται δίκλωνες ρήξεις που ενεργοποιούν τον μηχανισμό της σύνδεσης μη ομόλογων άκρων (βλ. Εικόνα 6.28), ο οποίος μπορεί να οδηγήσει σε ανασυνδυασμό αλλαγής τάξης. Ακαδημαϊκές Εκδόσεις 2011 Το κύτταρο-Μια Μοριακή Προσέγγιση.")
Παρόμοιες παρουσιάσεις