Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
ΚΙΝΗΤΙΚΗ ΑΝΑΠΤΥΞΗΣ ΜΙΚΡΟΒΙΩΝ ΚΑΙ ΠΑΡΑΓΩΓΗΣ ΜΕΤΑΒΟΛΙΚΩΝ ΠΡΟΪΟΝΤΩΝ

2
Μικροβιακή ανάπτυξη

3
Μέτρηση βιομάζας με φασματοφωτομετρία

4
Τριβλεία Petri

5
Ανάπτυξη καλλιέργειας

6
ΓΕΝΙΚΕΣ ΕΝΝΟΙΕΣ ΚΙΝΗΤΙΚΗΣ
Η αντίδραση μπορεί να αντιπροσωπεύει μία συνολική αντίδραση στην οποία περίπτωση, όπως είδαμε, οι στοιχειομετρικοί συντελεστές δεν είναι σταθεροί. Όταν δεν μεταβάλλονται αισθητά, μπορούμε να τους θεωρήσουμε σταθερούς και να χειριστούμε την αντίδραση σαν να ήταν απλή.

7
Ρυθμός αντίδρασης V ο όγκος του χώρου που λαμβάνει χώρα η αντίδραση
ξ η έκταση της αντίδρασης ξ*=ξ/V η ειδική έκταση Η τελευταία ισότητα ισχύει μόνο για συστήματα σταθερού όγκου.

8
Για κάθε ουσία Ni Ο ρυθμός εν γένει εξαρτάται από: την θερμοκρασία,
την πίεση την σύσταση Εκτός από τις ουσίες που συμπεριλαμβάνονται στην αντίδραση, υπάρχουν και άλλες ουσίες οι οποίες αν και παρούσες ως ιχνοστοιχεία μπορούν να επηρεάζουν σημαντικά τον ρυθμό αντίδρασης.

9
Μακριά από την ισορροπία:
φ(ci) συνάρτηση των συγκεντρώσεων ci και k μία "σταθερά" που ονομάζεται κινητική σταθερά. Η k είναι σταθερά μόνο όσο αφορά τις συγκεντρώσεις των αντιδρώντων και προϊόντων αλλά εξαρτάται από την θερμοκρασία με την σχέση Arrhenius: R η σταθερά των αερίων Τ η απόλυτη θερμοκρασία Α μία σταθερά που ονομάζεται προεκθετικός συντελεστής
 συνάρτηση των συγκεντρώσεων ci. και k μία σταθερά που ονομάζεται κινητική σταθερά. Η k είναι σταθερά μόνο όσο αφορά τις συγκεντρώσεις των αντιδρώντων και προϊόντων αλλά εξαρτάται από την θερμοκρασία με την σχέση Arrhenius: R η σταθερά των αερίων. Τ η απόλυτη θερμοκρασία. Α μία σταθερά που ονομάζεται προεκθετικός συντελεστής.")
10
Αν ο ρυθμός μπορεί να γραφεί στην μορφή:
τα βi είναι οι τάξεις της αντίδρασης ως προς τις ουσίες i Τέτοιου είδους εκφράσεις παίρνουμε για τις απλές αντιδράσεις κινητική δρώντων μαζών Τότε η τάξη ως προς μία ουσία συμπίπτει με τον αριθμό των μορίων που παίρνουν μέρος στην αντίδραση (μοριακότητα). Δηλαδή για κινητική δρώντων μαζών: βΑ=a, βB=b…
. Δηλαδή για κινητική δρώντων μαζών: βΑ=a, βB=b…")
11
Όταν δεν παρατηρείται κινητική δρώντων μαζών, τότε μπορούμε να συμπεράνουμε ότι πρόκειται για συνολική αντίδραση Το σύνολο των απλών αντιδράσεων που οδηγούν σε κάποια συνολική αντίδραση ονομάζεται μηχανισμός της συνολικής αντίδρασης. Η ύπαρξη πολλαπλών μηχανισμών συμβατών με την συνολική αντίδραση δυσχεραίνει το πρόβλημα της διαλεύκανσης του σωστού μηχανισμού. Δεν υπάρχει γενική μέθοδος μέτρησης του ρυθμού αντίδρασης. Αν η στοιχειομετρία είναι γνωστή (και σταθερή) αρκεί η παρακολούθηση μίας μόνο ουσίας ,μια και για κάθε i:
 αρκεί η παρακολούθηση μίας μόνο ουσίας ,μια και για κάθε i:")
12
Μέθοδοι επεξεργασίας κινητικών δεδομένων
Διακρίνουμε τρεις γενικές κατηγορίες: διαφορικές μεθόδους ολοκληρωτικές μεθόδους μεθόδους που βασίζονται στην απλοποίηση της παράστασης του ρυθμού (χρησιμοποιώντας π.χ. περίσσεια κάποιων αντιδρώντων)
")
13
Δύο Διαφορικές μέθοδοι
Μέθοδος Α: παραγώγιση των δεδομένων από μία και μοναδική πειραματική καμπύλη. Μέθοδος B: Με βάση πολλαπλές μετρήσεις του αρχικού ρυθμού αντίδρασης αλλάζοντας κάθε φορά την αρχική συγκέντρωση κάποιων αντιδρώντων.

14
Μέθοδος Α με δεδομένα τα ci συναρτήσει του t
παραγωγίζοντας γραφικά το διάγραμμα της συγκέντρωσης συναρτήσει του χρόνου γίνεται προσδιορισμός του ρυθμού συναρτήσει του χρόνου. υπολογίζοντας την παράσταση φ(ci) συναρτήσει του χρόνου και κατασκευάζοντας το διάγραμμα του ρυθμού r ως προς φ(ci) βλέπουμε αν τα σημεία κείνται σε ευθεία. η υπόθεση για την μορφή εξάρτησης ήταν σωστή και η κλίση της ευθείας μας δίνει την κινητική σταθερά. υποθέτουμε διαφορετική μορφή της φ(ci) και ξαναδοκιμάζουμε μέχρι να φέρουμε τα δεδομένα σε ευθεία γραμμή.
 συναρτήσει του χρόνου και κατασκευάζοντας το διάγραμμα του ρυθμού r ως προς φ(ci) βλέπουμε αν τα σημεία κείνται σε ευθεία. η υπόθεση για την μορφή εξάρτησης ήταν σωστή και η κλίση της ευθείας μας δίνει την κινητική σταθερά. υποθέτουμε διαφορετική μορφή της φ(ci) και ξαναδοκιμάζουμε μέχρι να φέρουμε τα δεδομένα σε ευθεία γραμμή.")
15
Μέθοδος πολλαπλών αρχικών συγκεντρώσεων

16
Ολοκληρωτικές μέθοδοι
Ολοκληρώνουμε μία έκφραση για τον ρυθμό που έχουμε αρχικά υποθέσει για να προβλέψουμε τις συγκεντρώσεις συναρτήσει του χρόνου Εάν το διάγραμμα του ψ(ci) συναρτήσει του t δώσει ευθεία γραμμή, η έκφραση που υποτέθηκε είναι σωστή, και η κλίση μάς δίνει την κινητική σταθερά. Εάν όχι τότε υποθέτουμε νέα έκφραση και να επαναλαμβάνουμε τη διαδικασία.
 συναρτήσει του t δώσει ευθεία γραμμή, η έκφραση που υποτέθηκε είναι σωστή, και η κλίση μάς δίνει την κινητική σταθερά. Εάν όχι τότε υποθέτουμε νέα έκφραση και να επαναλαμβάνουμε τη διαδικασία.")
17
Οι ρυθμοί

18
Ειδικοί ρυθμοί (ανά g κυτταρικής μάζας)
")
19
Ταξινόμηση μοντέλων μικροβιακής ανάπτυξης

20
Εξισορροπημένη ανάπτυξη
«Μέσο» κύτταρο

21
Καμπύλη ανάπτυξης φάση καθυστέρησης φάση επιτάχυνσης
φάση εκθετικής ανάπτυξης φάση επιβράδυνσης στάσιμη φάση φάση θανάτου ή αποδόμησης

22
ΜΗ ΔΟΜΗΜΕΝΑ - ΜΗ ΚΑΤΑΝΕΜΗΜΕΝΑ ΜΟΝΤΕΛΑ

23
ΜΟΝΤΕΛΑ ΑΝΑΠΤΥΞΗΣ ΜΙΑΣ ΜΕΤΑΒΛΗΤΗΣ

24
Malthus Ανεξέλεγκτη ανάπτυξη!
ο ειδικός ρυθμός ανάπτυξης μ είναι σταθερός. Ολοκληρώνοντας την εξίσωση με αρχική συνθήκη x(0)=xo: Ανεξέλεγκτη ανάπτυξη!
=xo: Ανεξέλεγκτη ανάπτυξη!")
25
Ενώ μπορεί να περιγράψει την ανάπτυξη στην εκθετική φάση, δεν μπορεί να προβλέψει άλλες φάσεις.
το μοντέλο αυτό είναι ανίκανο να προβλέψει την επήρεια στον ρυθμό ανάπτυξης οιασδήποτε παραμέτρου όπως συγκεντρώσεις θρεπτικών συστατικών κ.λ.π. Από την εξίσωση μπορούμε να προβλέψουμε τον χρόνο διπλασιασμού της βιομάζας:

26
Παράδειγμα: αποδόμηση σουλφαμεθοξαζόλης

27
Malthus model μ= d-1

28
Λογιστικό ή σιγμοειδές Verhulst 1844, Pearl και Reed το 1920
Προβλέπει εκθετική αλλά και στάσιμη

29
Λογιστικό k=0,328 d-1 β=0,032 l/mg

30
Πρόβλεψη λογιστικού μοντέλου
Ανάπτυξη μέχρι 315 mg/l σε 22 περίπου d Όμως στάσιμη φάση στις 10 d

31
Volterra περιλαμβάνει έναν επί πλέον όρο "μνήμης" αναγνωρίζοντας ότι οι παρελθούσες συνθήκες επηρεάζουν τον τρέχοντα ρυθμό ανάπτυξης είτε θετικά (ko>0) είτε αρνητικά (ko<0)
 είτε αρνητικά (ko<0)")
32
Τα μοντέλα μιας μεταβλητής:
είναι όλα εμπειρικά στην φύση τους δεν προβλέπουν φάση καθυστέρησης δεν περιγράφουν τις επιδράσεις των διαφόρων παραγόντων στον ρυθμό ανάπτυξης δεν κάνουν καμία χρήση των γνώσεων της βιοχημείας και μικροβιολογίας.

33
ΜΟΝΤΕΛΑ ΑΝΑΠΤΥΞΗΣ ΜΕ ΠΕΡΙΟΡΙΣΜΟ ΥΠΟΣΤΡΩΜΑΤΟΣ
Διάφορα μοντέλα ανάλογα με τη συνάρτηση

34
Το μοντέλο Monod όπου: S: περιοριστικό υπόστρωμα (mg/L)
Ks: σταθερά κορεσμού (ίση με την συγκέντρωση του υποστρώματος στην οποία ο ρυθμός ανάπτυξης είναι ίσος με το ήμισυ του μέγιστου ειδικού ρυθμού ανάπτυξης) μmax: μέγιστος ειδικός ρυθμός ανάπτυξης
 μmax: μέγιστος ειδικός ρυθμός ανάπτυξης.")
35
Παράδειγμα σουλφομεθοξαζόλης
Kinetic model Kinetic constants μmax = 0.21 d-1 Ks = mg/L Yx/s = 0.95

36
Εναλλακτικές εκφράσεις

37
Εναλλακτικές εκφράσεις

38
Konak Το μοντέλο αυτό είναι υπό μορφή κανονικής διαφορικής εξίσωσης

39
Γενικευμένη εξίσωση Kargi και Shuler
μrel=μ/μmax

40
Παρεμπόδιση υποστρώματος
μmax=0,5 d-1 Κs=30 mg/l KΙ=100 mg/l Andrews

41
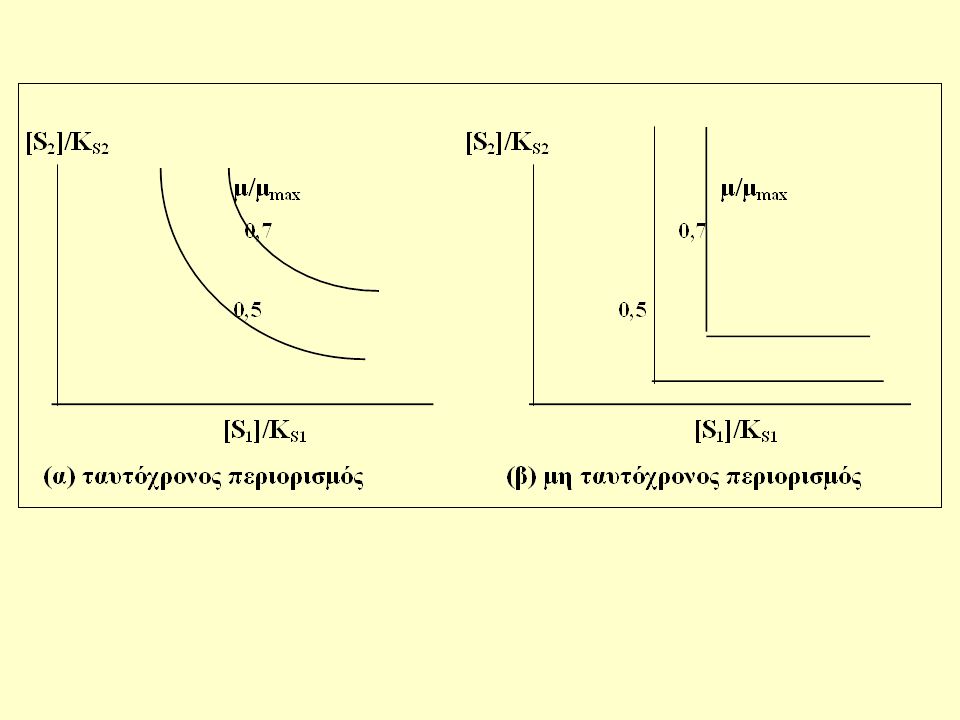
Πολλαπλά υποστρώματα Διαδοχική χρήση (διαυξική ανάπτυξη εναλλακτικών υποστρωμάτων) Ταυτόχρονη χρήση (συμπληρωματικών υποστρωμάτων) Οι περιπτώσεις της ταυτόχρονης χρήσης διακρίνονται περαιτέρω επί τη βάσει του αν ο περιορισμός είναι ταυτόχρονος ή όχι.

42
Ταυτόχρονος περιορισμός

43
Μη ταυτόχρονος περιορισμός

45
Διαδοχική χρήση

46
Επαύξηση Στην περίπτωση επαύξησης του ρυθμού από την παρουσία κάποιας ένωσης SE ακολουθώντας τους Tsao και Hanson έχουμε την ακόλουθη τροποποίηση του μέγιστου ειδικού ρυθμού ανάπτυξης:

48
Μοντέλα καθυστέρησης O’Neil and Lyberatos Wang and Stephanopoulos

49
ΣΥΝΤΗΡΗΣΗ ΚΑΙ ΕΝΔΟΓΕΝΗΣ ΜΕΤΑΒΟΛΙΣΜΟΣ
κατανάλωση υποστρώματος όχι μόνο για ανάπτυξη αλλά και για συντήρηση με ρυθμό ανάλογο της συγκέντρωσης της ζώσας βιομάζας: όπου xv η συγκέντρωση της ζώσας βιομάζας. Η τιμή της σταθεράς ms εξαρτάται από τις συνθήκες. όσο πιο αντίξοες οι συνθήκες, τόσο μεγαλύτερη και η σταθερά

50
Ενδογενής μεταβολισμός ή ενδογενής αναπνοή
Η ενέργεια που απαιτείται για την διατήρηση ζώντων οργανισμών και γίνεται εις βάρος της βιομάζας δεν επηρεάζεται το ισοζύγιο του υποστρώματος αλλά από τον ρυθμό ανάπτυξης βιομάζας αφαιρούμε ένα ρυθμό ενδογενούς μεταβολισμού:

51
Ταξινόμηση μεταβολικών προϊόντων με βάση την κινητική

52
ΜΟΝΤΕΛΑ ΓΙΑ ΤΗΝ ΧΡΗΣΗ ΥΠΟΣΤΡΩΜΑΤΟΣ
όπου P1,...,Pn είναι τα εξωκυτταρικά προϊόντα, και YPi/S είναι οι συντελεστές απόδοσης που συσχετίζουν το υπόστρωμα με το προϊόν i. Υποθέτοντας αμελητέα χρήση για συντήρηση και ότι όλα τα προϊόντα είναι συσχετιζόμενα με την ανάπτυξη:

53
Πότε ισχύει το κάτωθι; Όταν: 1. όλα τα προϊόντα είναι σχετιζόμενα με την ανάπτυξη 2. η συντήρηση και ο ενδογενής μεταβολισμός μπορούν να αγνοηθούν

54
Φάση καθυστέρησης Dean και Hinshelwood
ο χρόνος καθυστέρησης tL είναι ο χρόνος μετά την μεταφορά της μαγιάς στο νέο θρεπτικό μέσο στο οποίο κάποιο κρίσιμο ένζυμο φτάνει σε κάποια κρίσιμη συγκέντρωση c* V: ο λόγος των όγκων του προηγούμενου μέσου προς τον όγκο του νέου Νο : ο αριθμός των παλαιών κυττάρων ανά μονάδα όγκου στο νέο μέσο a: η συγκέντρωση του ενζύμου στο προηγούμενο μέσο a’: ο ρυθμός παραγωγής του ενζύμου από τα "παλαιά" κύτταρα ανά κύτταρο a" : ο ρυθμός παραγωγής του ενζύμου από νέα κύτταρα Προκειμένου να ελαχιστοποιήσουμε τον χρόνο καθυστέρησης χρειάζεται μεγάλη μαγιά (No) και μεγάλος λόγος όγκων (V). Στην πράξη προσπαθούμε να χρησιμοποιούμε όγκο μαγιάς περίπου το ένα δέκατο του όγκου του νέου μέσου.
 και μεγάλος λόγος όγκων (V). Στην πράξη προσπαθούμε να χρησιμοποιούμε όγκο μαγιάς περίπου το ένα δέκατο του όγκου του νέου μέσου.")
55
Στάσιμη φάση Για την στάσιμη φάση κατ' αρχήν διακρίνουμε δύο πιθανούς λόγους διακοπής της ανάπτυξης. ο πρώτος είναι η εξάντληση κάποιου θρεπτικού συστατικού ο δεύτερος η συσσώρευση κάποιου τοξικού προϊόντος. Στην πρώτη περίπτωση η τελική συγκέντρωση της βιομάζας θα είναι απλώς:

56
Στάσιμη φάση Στη δεύτερη περίπτωση πρέπει να βρούμε την εξάρτηση του ειδικού ρυθμού ανάπτυξης από το τοξικό προϊόν και να εκφράσουμε τον ρυθμό παραγωγής του τοξικού προϊόντος συναρτήσει του υποστρώματος και της βιομάζας. Η ανάπτυξη θα διακοπεί όταν ο ειδικός ρυθμός ανάπτυξης μηδενιστεί, δηλαδή όταν η συγκέντρωση φτάσει κάποιο συγκεκριμένο επίπεδο. Επιλύνοντας τα ισοζύγια από κοινού βρίσκουμε τις τελικές συγκεντρώσεις του υποστρώματος και της βιομάζας.

57
Στάσιμη φάση Για να μπορέσουμε να διακρίνουμε πειραματικά τον λόγο της διακοπής της ανάπτυξης, δεν έχουμε παρά να αραιώσουμε ένα δείγμα με νερό. Αν παρατηρηθεί περαιτέρω ανάπτυξη ο λόγος είναι η συσσώρευση τοξίνης, ενώ αν δεν παρατηρηθεί πρόκειται σίγουρα για εξάντληση υποστρώματος.

58
Φάση θανάτου Η σταθερά kd είναι συνάρτηση της θερμοκρασίας και προσδιορίζεται εύκολα από την κλίση διαγράμματος του λογαρίθμου της βιομάζας συναρτήσει του χρόνου.

59
ΔΟΜΗΜΕΝΑ ΜΗ ΚΑΤΑΝΕΜΗΜΕΝΑ ΜΟΝΤΕΛΑ
Όταν ενδιαφέρει η εξέλιξη κάποιου συγκεκριμένου κυτταρικού συστατικού, ή απαιτείται η αναγνώριση των κύριων συστατικών του κυττάρου για την επαρκή περιγραφή της συνολικής κινητικής ανάπτυξης (όταν δηλαδή η παραδοχή της εξισορροπημένης ανάπτυξης δεν ευσταθεί) πρέπει να επιστρατεύσουμε τις γνώσεις μας για την βιοχημεία και φυσιολογία του κυττάρου για την ανάπτυξη κατάλληλων δομημένων μοντέλων.
 πρέπει να επιστρατεύσουμε τις γνώσεις μας για την βιοχημεία και φυσιολογία του κυττάρου για την ανάπτυξη κατάλληλων δομημένων μοντέλων.")
60
Πως συνδέεται η συγκέντρωση κάποιου συγκεκριμένου ενδοκυτταρικού παράγοντα με τον ρυθμό ανάπτυξης του κυττάρου που αναπτύσσεται σε κάποιο μέσο όγκου VR? Ας υποθέσουμε ότι το σύστημα είναι κλειστό. Έστω cj η συγκέντρωση του συστατικού j μέσα στο κύτταρο, σε moles j ανά μονάδα κυτταρικού όγκου. Αν η συγκέντρωση της βιομάζας είναι x (σε g/l) τότε η συνολική βιομάζα είναι VRx Αν η πυκνότητα ρc των κυττάρων θεωρείται σταθερή, τότε ο ολικός όγκος της βιολογικής φάσεως (του συνόλου των κυττάρων) είναι (VRx)/ρc. Αρα ο συνολικός αριθμός των moles του j μέσα στο μέσο είναι (VRxcj)/ρc.
 τότε η συνολική βιομάζα είναι VRx. Αν η πυκνότητα ρc των κυττάρων θεωρείται σταθερή, τότε ο ολικός όγκος της βιολογικής φάσεως (του συνόλου των κυττάρων) είναι (VRx)/ρc. Αρα ο συνολικός αριθμός των moles του j μέσα στο μέσο είναι (VRxcj)/ρc.")
62
Ο πρώτος όρος είναι ο καθαρός ρυθμός μεταβολής λόγω αντίδρασης
Ο δεύτερος όρος αντιπροσωπεύει την μείωση που παρατηρείται εξ αιτίας της αραίωσης που προκύπτει από την αύξηση του κυτταρικού όγκου λόγω ανάπτυξης. Η σχέση αυτή είναι ιδιαίτερα σημαντική μια και χρησιμοποιείται για την κατασκευή οιουδήποτε δομημένου μοντέλου.

63
ΔΙΑΜΕΡΙΣΜΑΤΙΚΑ ΔΟΜΗΜΕΝΑ ΜΟΝΤΕΛΑ
Τα διαμερισματικά μοντέλα χωρίζουν την βιομάζα σε επί μέρους τμήματα. Έτσι π.χ. μπορούμε να διακρίνουμε ένα συνθετικό μέρος βιομάζας (βασικά RNA) και ένα δομικό μέρος (DNA και πρωτείνες). Εναλλακτικά μπορούμε να διακρίνουμε την βιομάζα σε ένα αφομοιωτικό τμήμα και ένα συνθετικό τμήμα, κ.ο.κ. Ο διαχωρισμός σε τμήματα έχει κάποια βιοχημική και δομική βάση, αλλά δεν αντιστοιχεί επακριβώς σε κάποιο πραγματικό μηχανισμό.
 και ένα δομικό μέρος (DNA και πρωτείνες). Εναλλακτικά μπορούμε να διακρίνουμε την βιομάζα σε ένα αφομοιωτικό τμήμα και ένα συνθετικό τμήμα, κ.ο.κ. Ο διαχωρισμός σε τμήματα έχει κάποια βιοχημική και δομική βάση, αλλά δεν αντιστοιχεί επακριβώς σε κάποιο πραγματικό μηχανισμό.")
64
Williams Η βιομάζα αποτελείται από δύο τμήματα 1 και 2. Το πρώτο είναι συνθετικό και το δεύτερο δομικό. Το συνθετικό μέρος παράγεται με την ταυτόχρονη κατανάλωση υποστρώματος και διέπεται από συντελεστή απόδοσης Υ που δίνεται σε g μέρους 1 / g υποστρώματος Το δεύτερο δομικό μέρος βιομάζας παράγεται με ρυθμό ανάλογο του γινομένου ρ1ρ2 όπου ρi η συγκέντρωση του μέρους i μέσα στο κύτταρο Αναγκαία και ικανή συνθήκη για διχοτόμηση αποτελεί ο διπλασιασμός του μέρους 2

66
το μοντέλο υποθέτει ότι ο ειδικός ρυθμός ανάπτυξης είναι ανάλογος της συγκέντρωσης υποστρώματος (μ=k1[S]) και ότι η πυκνότητα της βιομάζας ρκ=ρ1+ρ2 είναι σταθερή. Ουσιαστικά οι δύο πρώτες εξισώσεις είναι ισοδύναμες με το μοντέλο δύο φάσεων μια και μπορούν να επιλυθούν ανεξάρτητα από τις δύο τελευταίες. Επίσης πολλαπλασιάζοντας την πρώτη με Υ και προσθέτοντας με την δεύτερη βλέπουμε ότι το άθροισμα Υ[S]+x είναι σταθερό και ίσο με Υ[Sο]+xο. Αντικαθιστώντας το [S] συναρτήσει του x καταλήγουμε ουσιαστικά στην λογιστική εξίσωση των Verhulst και Pearl. Ορίζοντας f2=ρ2/ρκ (το κλάσμα του τμήματος 2) παίρνουμε:
![το μοντέλο υποθέτει ότι ο ειδικός ρυθμός ανάπτυξης είναι ανάλογος της συγκέντρωσης υποστρώματος (μ=k1[S]) και ότι η πυκνότητα της βιομάζας ρκ=ρ1+ρ2 είναι σταθερή.](http://slideplayer.gr/slide/2632963/9/images/66/%CF%84%CE%BF+%CE%BC%CE%BF%CE%BD%CF%84%CE%AD%CE%BB%CE%BF+%CF%85%CF%80%CE%BF%CE%B8%CE%AD%CF%84%CE%B5%CE%B9+%CF%8C%CF%84%CE%B9+%CE%BF+%CE%B5%CE%B9%CE%B4%CE%B9%CE%BA%CF%8C%CF%82+%CF%81%CF%85%CE%B8%CE%BC%CF%8C%CF%82+%CE%B1%CE%BD%CE%AC%CF%80%CF%84%CF%85%CE%BE%CE%B7%CF%82+%CE%B5%CE%AF%CE%BD%CE%B1%CE%B9+%CE%B1%CE%BD%CE%AC%CE%BB%CE%BF%CE%B3%CE%BF%CF%82+%CF%84%CE%B7%CF%82+%CF%83%CF%85%CE%B3%CE%BA%CE%AD%CE%BD%CF%84%CF%81%CF%89%CF%83%CE%B7%CF%82+%CF%85%CF%80%CE%BF%CF%83%CF%84%CF%81%CF%8E%CE%BC%CE%B1%CF%84%CE%BF%CF%82+%28%CE%BC%3Dk1%5BS%5D%29+%CE%BA%CE%B1%CE%B9+%CF%8C%CF%84%CE%B9+%CE%B7+%CF%80%CF%85%CE%BA%CE%BD%CF%8C%CF%84%CE%B7%CF%84%CE%B1+%CF%84%CE%B7%CF%82+%CE%B2%CE%B9%CE%BF%CE%BC%CE%AC%CE%B6%CE%B1%CF%82+%CF%81%CE%BA%3D%CF%811%2B%CF%812+%CE%B5%CE%AF%CE%BD%CE%B1%CE%B9+%CF%83%CF%84%CE%B1%CE%B8%CE%B5%CF%81%CE%AE..jpg "Ουσιαστικά οι δύο πρώτες εξισώσεις είναι ισοδύναμες με το μοντέλο δύο φάσεων μια και μπορούν να επιλυθούν ανεξάρτητα από τις δύο τελευταίες. Επίσης πολλαπλασιάζοντας την πρώτη με Υ και προσθέτοντας με την δεύτερη βλέπουμε ότι το άθροισμα Υ[S]+x είναι σταθερό και ίσο με Υ[Sο]+xο. Αντικαθιστώντας το [S] συναρτήσει του x καταλήγουμε ουσιαστικά στην λογιστική εξίσωση των Verhulst και Pearl. Ορίζοντας f2=ρ2/ρκ (το κλάσμα του τμήματος 2) παίρνουμε:")
67
Από την παραδοχή για διχοτόμηση φαίνεται ότι ο αριθμός των κυττάρων είναι ανάλογος του f2x
επομένως το μέγεθος των κυττάρων είναι ανάλογο του x/(f2x)=1/f2
=1/f2.")
68
Καλλιέργεια σε αντιδραστήρα διαλείποντος έργου με μαγιά που προέρχεται από στάσιμη φάση (ρ1=0)
K1=0,005 K2=0,1 Y=0,5 So=270 Xo=5 Σχετικό μέγεθος 1/f2-1 Βιομάζα x/(Y.So) Υπόστρωμα S/So Αριθμός f2.x/(Yso)
 Υπόστρωμα S/So. Αριθμός f2.x/(Yso)")
69
Προβλέπει ορισμένα ποιοτικά χαρακτηριστικά που συνήθως παρατηρούνται σε πραγματικές καλλιέργειες:
(α) Ύπαρξη φάσης καθυστέρησης κατά την οποία αυξάνεται το μέγεθος των κυττάρων χωρίς αύξηση του αριθμού τους. (β) Ύπαρξη φάσης εκθετικής ανάπτυξης κατά την οποία και το μέσο κυτταρικό μέγεθος παίρνει την μεγαλύτερη του τιμή. (γ) Μεταβολή της σύστασης των κυττάρων κατά τον κύκλο ανάπτυξης. (δ) Ύπαρξη στάσιμης φάσης με μικρά σε μέγεθος κύτταρα. (ε) Ανομοιόμορφη μεταβολή του αριθμού και της μάζας των κυττάρων.
 Ύπαρξη φάσης καθυστέρησης κατά την οποία αυξάνεται το μέγεθος των κυττάρων χωρίς αύξηση του αριθμού τους. (β) Ύπαρξη φάσης εκθετικής ανάπτυξης κατά την οποία και το μέσο κυτταρικό μέγεθος παίρνει την μεγαλύτερη του τιμή. (γ) Μεταβολή της σύστασης των κυττάρων κατά τον κύκλο ανάπτυξης. (δ) Ύπαρξη στάσιμης φάσης με μικρά σε μέγεθος κύτταρα. (ε) Ανομοιόμορφη μεταβολή του αριθμού και της μάζας των κυττάρων.")
70
ΜΗΧΑΝΙΣΤΙΚΑ ΔΟΜΗΜΕΝΑ ΜΟΝΤΕΛΑ
Τα μηχανιστικά μοντέλα κάνουν άμεση χρήση των γνώσεων που έχουμε για τον μεταβολισμό ή/και την γενετική του κυττάρου. τα μοντέλα αυτά περιλαμβάνουν μόνο στοιχεία που: κρίνονται ως απαραίτητα για την περιγραφή της κινητικής (μια και παίζουν καθοριστικό ρόλο στην παρατηρούμενη κινητική και στοιχειομετρία) ή συνδέονται άμεσα με την κινητική επί μέρους συστατικών που μας ενδιαφέρουν (π.χ. προϊόντων). καθοριστικό ρόλο παίζει η δυνατότητα μέτρησης των μεταβλητών που περιλαμβάνονται στο μοντέλο.
 ή. συνδέονται άμεσα με την κινητική επί μέρους συστατικών που μας ενδιαφέρουν (π.χ. προϊόντων). καθοριστικό ρόλο παίζει η δυνατότητα μέτρησης των μεταβλητών που περιλαμβάνονται στο μοντέλο.")
71
Παραγωγή κάποιας πρωτείνης P που λαμβάνει υπόψη του τις διεργασίες της μεταγραφής και μετάφρασης:

72
Εδώ ξ είναι ο συντελεστής αποδοτικότητας της μετάφρασης.

73
Αν υποθέσουμε εξισορροπημένη ανάπτυξη,
οι χρονικές παράγωγοι στο αριστερό μέρος των εξισώσεων μηδενίζονται, και οι αλγεβρικές εξισώσεις που προκύπτουν δίνουν: Η σχέση αυτή δίνει την συγκέντρωση του πρωτεϊνικού προϊόντος συναρτήσει των ρυθμών ανάπτυξης, μεταγραφής και μετάφρασης

74
ΚΙΝΗΤΙΚΗ ΑΠΟΣΤΕΙΡΩΣΗΣ
Αποστείρωση ενός μέσου ονομάζουμε την εξόντωση ή απομάκρυνση όλων των μικροοργανισμών με ταυτόχρονη απενεργοποίηση όλων των ιών. Η αποστείρωση είναι μία ιδιαίτερα βασική διεργασία που συνήθως προηγείται της κύριας βιοτεχνολογικής διεργασίας που περιλαμβάνει ανάπτυξη μικροοργανισμών (αποστείρωση θρεπτικού μέσου).
.")
75
Στις φαρμακοβιομηχανίες και στις βιομηχανίες τροφίμων χρησιμοποιούνται μία ή περισσότερες από τις ακόλουθες βασικές μεθόδους αποστείρωσης: (α) υγρή θέρμανση (ατμοί) (β) ξερή θέρμανση (κλίβανοι) (γ) μικροβιοκτόνοι παράγοντες όπως χλώριο, οξείδιο του αιθυλενίου και φορμαλδεύδη (δ) ακτινοβολία (ε) διήθηση Μία κατάλληλη επεξεργασία φέρνει την επιθυμητή απομάκρυνση μικροοργανισμών χωρίς να προκαλέσει βλάβες στα προϊόντα. Η πιο συνηθισμένη διεργασία είναι η χρήση υπέρθερμων ατμών.
 υγρή θέρμανση (ατμοί) (β) ξερή θέρμανση (κλίβανοι) (γ) μικροβιοκτόνοι παράγοντες όπως χλώριο, οξείδιο του αιθυλενίου και φορμαλδεύδη. (δ) ακτινοβολία. (ε) διήθηση. Μία κατάλληλη επεξεργασία φέρνει την επιθυμητή απομάκρυνση μικροοργανισμών χωρίς να προκαλέσει βλάβες στα προϊόντα. Η πιο συνηθισμένη διεργασία είναι η χρήση υπέρθερμων ατμών.")
77
Κινητική αποστείρωσης

79
Μοντέλο δύο φάσεων Πολλοί οργανισμοί δεν εξουδετερώνονται άμεσα, αλλά μετατρέπονται πρώτα σε σπόρια, τα οποία εξουδετερώνονται στη συνέχεια με άλλη κινητική. Σε αυτή την περίπτωση έχουμε: ΝR NS ND με σταθερές KR και KS αντίστοιχα

80
Η επίλυση με ΝR(0)=No και ΝS(0)=0 δίνει:
όπου Ν=ΝR +NS

82
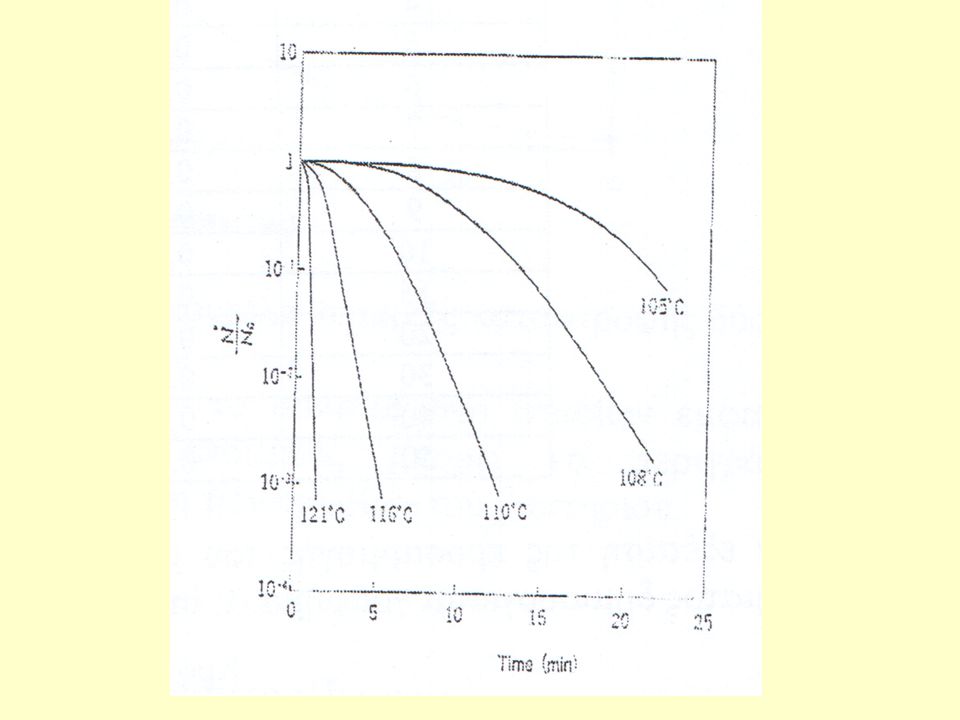
Αριθμός ζωντανών σπορίων (Ν)
ΠΡΟΒΛΗΜΑ Κατά τη θερμική απενεργοποίηση σπορίων βακτηρίου στους 121ºC προέκυψαν τα δεδομένα του Πίνακα που παρατίθεται. Ποιο πρότυπο ακολουθεί η θερμική καταστροφή του μικροοργανισμού; Εκτιμείστε την προσαρμογή των δεδομένων στο προτεινόμενο μοντέλο και υπολογίστε τις σχετικές σταθερές. Χρόνος (sec) Αριθμός ζωντανών σπορίων (Ν) 100 8 75 1 99 9 70 2 98 10 66 3 95 15 46 4 91 20 31 5 88 30 13 6 83 40 7 79 50
 Αριθμός ζωντανών σπορίων (Ν)")
83
Επίλυση Χρόνος (sec) Ν/Νο ln(Ν/No) 1,000 0,000 1 0,990 -0,010 2 0,980
1,000 0,000 1 0,990 -0,010 2 0,980 -0,020 3 0,950 -0,051 4 0,910 -0,094 5 0,880 -0,128 6 0,830 -0,186 7 0,790 -0,236 8 0,750 -0,288 9 0,700 -0,357 10 0,660 -0,416 15 0,460 -0,777 20 0,310 -1,171 30 0,130 -2,040 40 0,050 -2,996 50 0,020 -3,912

84
Η προσαρμογή ευθείας στα σημεία του γραφήματος δίνει συντελεστή συσχέτισης R2=0,9875
ενώ από την κλίση της ευθείας υπολογίζεται ο ειδικός ρυθμός θερμικής καταστροφής k=0,080 sec-1.

85
Στη συνέχεια εξετάζεται η προσαρμογή του μοντέλου θερμικής καταστροφής δύο σταδίων Επιλύοντας την εξίσωση χρησιμοποιώντας αλγόριθμο μη γραμμικής βελτιστοποίησης προκύπτει συντελεστής συσχέτισης R2=0.9999, ενώ οι τιμές των σταθερών υπολογίζονται ίσες με: KS = σπόρια s KR = σπόρια s-1 Από τα δύο πρότυπα που εξετάστηκαν, το μοντέλο δύο σταδίων εμφανίζει τον υψηλότερο συντελεστή συσχέτισης, επομένως μπορεί να περιγράψει πιο ικανοποιητικά τη θερμική απενεργοποίηση των σπορίων του βακτηρίου.

86
ΑΠΟΛΥΜΑΝΣΗ Απολύμανση ονομάζουμε την επιλεκτική μείωση οργανισμών που μπορεί να μη συνοδεύεται από πλήρη εξόντωση σαπροφυτικών οργανισμών και ενδόσπορων. Η απολύμανση απαντάται στην πράξη σε ιατρικές εφαρμογές καθώς και στην επεξεργασία νερού και λυμάτων. Για τον καθαρισμό του νερού το πιο συνηθισμένο απολυμαντικό μέσο είναι το χλώριο. Η κινητική της απολύμανσης είναι ανάλογη της κινητικής θερμικής αποστείρωσης (κινητική πρώτης τάξης ως προς την συγκέντρωση οργανισμών).
.")
87
Κινητική απολύμανσης Η εξάρτηση της κινητικής από την συγκέντρωση του απολυμαντικού παράγοντα δίνεται συνήθως εμπειρικά από την σχέση: Cnt=σταθερά όπου: t ο χρόνος που απαιτείται για να επιτευχθεί μία δεδομένη ποσοστιαία μείωση του πληθυσμού C η συγκέντρωση του απολυμαντικού παράγοντα. π.χ. αν t είναι ο χρόνος για 99% μείωση του πληθυσμού E.coli σε διάλυμα υποχλωριώδους οξέος, η σταθερά είναι 0,24 ενώ n=0,86.

Παρόμοιες παρουσιάσεις