Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Galvenais audu saderības komplekss
Major Histocompatibility Complex (MHC)
")
2
Lekcijas plāns Galvenais audu saderības komplekss (MHC)
MHC kompleksa atklāšana un tā loma imūnatbildē MHC organizācija genomā MHC molekulu struktūra Peptīdu saistīšanās pie MHC molekulām MHC molekulu producēšana Antigēna apstrāde un prezentēšana T limfocītiem To antigēnu īpašības, kurus atpazīst T limfocīti Antigēnprezentējošās šūnas Antigēna apstrāde Lipīdu antigēnu prezentēšana Ilona Mandrika

3
Kā tika atklāts Galvenais audu saderības komplekss?
MHC molekulas tika atklātas pētot audu atgrūšanu peļu līnijās un trušos. 1916.g. Pārstādot audzējus starp dažādām peļu līnijām novēroja, ka audzēji tiek pieņemti vienas līnijas ietvaros, bet atgrūsti dažādu peļu līniju ietvaros. 1927.g. Secināts, ka pārstādot audus, tie netiek atgrūsti, ja donors un recipients ir identiski dvīņi. Audu saderība ir atkarīga no ģēnētikas

4
Kā tika atklāts Galvenais audu saderības komplekss?
George Snell un viņa kolēģi ( ) Transplanti vienas peļu līnijas Ietvaros tiek pieņemti. Transplanti starp dažādām peļu līnijām tiek atgrūsti. Transplanta atpazīšana par savu vai svešu ir iedzimta īpašība. Sistemātiski krustoja peļu līnijas, kuras bija ģenētiski identiskas, izņemot vienu ģenētisko rajonu (congenic strains).
 Transplanti vienas peļu līnijas Ietvaros. tiek pieņemti. Transplanti starp dažādām. peļu līnijām tiek atgrūsti. Transplanta atpazīšana. par savu vai svešu ir. iedzimta īpašība. Sistemātiski krustoja peļu līnijas, kuras bija ģenētiski identiskas, izņemot vienu ģenētisko rajonu (congenic strains).")
5
Peļu audu saderības lokuss tika sasaistīts ar gēnu
17.hromosomā, kas kodē asins grupas antigēnu (antigēnu II). Šo gēnu rajonu nosauca par Histocompatibility- 2 vai vienkārši H-2. Ģenētisko reģiona daļu, kas kontrolēja audu atgrūšanu un sastāvēja no vairākiem saistītiem gēniem nosauca par Galveno audu saderības kompleksu - Major Histocompatibility Complex (MHC).
. Šo gēnu rajonu nosauca par Histocompatibility- 2. vai vienkārši H-2. Ģenētisko reģiona daļu, kas kontrolēja audu atgrūšanu. un sastāvēja no vairākiem saistītiem gēniem nosauca par. Galveno audu saderības kompleksu - Major Histocompatibility Complex (MHC).")
6
Galvenais Audu Saderības Komplekss
(Major Histocompatibility Complex) Genoma rajons, kas kodē vairākus gēnus (tāpēc Komplekss), kuri spēlē svarīgu lomu (tāpēc Galvenais) audu translantācijā (tāpēc Audu Saderības).
 Genoma rajons, kas kodē vairākus gēnus (tāpēc Komplekss), kuri spēlē svarīgu lomu (tāpēc Galvenais) audu translantācijā (tāpēc Audu Saderības).")
7
Turpmākajos 20 gados vienīgā zināmā loma MHC kompleksam
bija audu atgrūšana. Transplantācija nav gluži dabīgs fenomens. Kāpēc tad MHC gēni tiek saglabāti evolūcijas gaitā? gados tika atklāts, ka MHC gēniem ir fundamentāla loma visās imūnajās atbildēs pret proteīnu antigēniem.

8
Pirmie trīs gēni tika nosaukti
Cilvēka MHC molekulas = Cilvēka Leikocītu Antigēni Human Leukocyte Antigens HLA = H-2 pelēs Jean Dausset pirmais parādīja, ka indivīdiem, kuri atgrūda nieri, parādījās cirkulējošās antivielas, reaģējošas ar donora limfocītu antigēniem. Šie antigēni (alloantigēni) ir polimorfo gēnu produkti, kuri spēj atšķirt savu no sveša. Pirmie trīs gēni tika nosaukti HLA-A, HLA-B, HLA-C. Baruj Benacerraf atklāja, ka MHC gēni kontrolē organisma spēju reaģēt uz antigēniem.
 ir polimorfo gēnu produkti, kuri. spēj atšķirt savu no sveša. Pirmie trīs gēni tika nosaukti. HLA-A, HLA-B, HLA-C. Baruj Benacerraf atklāja, ka MHC gēni kontrolē. organisma spēju reaģēt uz antigēniem.")
9
Nobela prēmija Fizioloģijā vai Medicīnā 1980. gadā
“par viņu atklājumiem attiecībā uz ģenētiski noteiktām struktūrām uz šūnu virsmas, kas regulē imunoloģiskās reakcijas" George D. Snell Jean Dausset Baruj Benacerraf ASV Francija ASV

10
MHC organizācija genomā

11
MHC organizācija genomā
Cilvēkam MHC atrodas 6.hromosomas īsajā plecā un aptver kb (0,1% no cilvēka genoma). Class II Class III Class I - 20 gēni - 30 gēni - 15 gēni Antigēna (peptīda) prezentācija Komplementa sistēmas proteīni Antigēna (peptīda) prezentācija b2-mikroglobulīnu kodē gēns, kas atrodas 15.hromosomā
. Class II. Class III. Class I gēni gēni gēni. Antigēna (peptīda) prezentācija. Komplementa. sistēmas proteīni. Antigēna (peptīda) prezentācija. b2-mikroglobulīnu kodē gēns, kas atrodas 15.hromosomā.")
12
Kodē gēnus, kas kontrolē visas specifiskās imūnatbildes.
Class II Class III Class I Klasiskie MHC gēni Class I Class II Kodē gēnus, kas kontrolē visas specifiskās imūnatbildes. Galvenā funkcija ir antigēna peptīdu prezentēšana T limfocītiem. Nepiedalās antigēna peptīdu prezentēšanā T limfocītiem. Spēlē lomu imūnatbildēs. MHC rajons Kodē arī gēnus, kas ietekmē augšanu, attīstību, vairošanos, smaržas, ožu.

13
HLA-A HLA-C HLA-B TAP 1 un TAP 2 – transporta proteīnu gēni (nozīmīgi antigēnu pārstrādē) TAP - transporter associated with antigen processing
 TAP - transporter associated with antigen processing.")
14
MHC ir poligēns, t.i. satur vairākus dažādus MHC I un II klases gēnus. MHC ir ar gēniem blīvākais rajons cilvēka genomā.

15
MHC ir polimorfs, t.i. populācijā gēnam ir vairāki varianti.
MHC gēni ir vispolimorfiskākie gēni visā genomā. HLA alēļu skaits (2010.gads) HLA alēļu skaits (2014.g.) HLA I.klases alēles 8976 HLA II. klases alēles 2870 HLA kopējais alēļu skaits 11846
 HLA alēļu skaits (2014.g.) HLA I.klases alēles HLA II. klases alēles HLA kopējais alēļu skaits")
16
MHC polimorfisms veidojas gēnu konversijas un
rekombinācijas procesu rezultātā Gēnu konversija Hromosomu nobīde mejozes laikā MHC gēni Pēc hromosomu atdalīšanas mejozē DNS ir samainīta starp haplotipiem. Rekombinācija starp haplotipiem Abos variantos MHC tips saglabājās, bet var izveidoties jauns alēļu variants.

17
MHC monomorfisms Ne visām zīdītāju sugām ir augsts MHC polimorfisms.

18
MHC molekulu polimorfisms
MHC molekulu dažādība nevar pasargāt katru indivīdu no jebkura patogēna. Bet MHC polimorfisms pasargā populāciju no iznīcības.

19
MHC molekulu polimorfisms
MHC molekulu dažādība pastāv populācijas līmenī. MHC molekulas tips un variants cilvēka dzīves laikā nemainās. Tas ir pretstatā ar T un B šūnu antigēna receptoriem, kuri mainās cilvēka dzīves laikā.

20
MHC gēni ir kodominanti ekspresēti katrā indivīdā.
Katrā indivīdā vienādā pakāpē tiek ekspresētas mātes un tēva alēles. Tas ļauj palielināt MHC molekulu skaitu nepieciešamo peptīdu piesaistei un T-šūnu prezentācijai. Katrs indivīds ekspresē 6 veidu MHC I klases molekulas uz katras šūnas un 6 vai 8 MHC II klases molekulas. DR α un β-ķēdes var savienoties cis (abas no viena vecāka) vai trans (katra no sava vecāka) kombinācijās. Turklāt DR β-ķēdi var kodēt vairāk nekā viens gēns.
 vai. trans (katra no sava vecāka) kombinācijās. Turklāt DR β-ķēdi. var kodēt vairāk nekā viens gēns.")
21
Šobrīd MHC gēnu lokuss ir viena no visvairāk pētītām
ģenētiskām sistēmām. Ietekmē Infekcijas slimību rezistenci Autoimunitāti Audu saderību transplantācijā Spontānos abortus

22
Izmaiņas MHC lokusā ir saistītas ar
dažāda veida slimībām (>100). ► Monogēnās slimības (hemohromatoze saistīta ar MHCI) ► Infekcijas (HIV-AIDS) ► Autoimūnās slimības (psoriāze, astma, reimatoīdais artrīts) ► Vēzis (Hodžkina limfoma) ► Jūtība pret zālēm
. ► Monogēnās slimības (hemohromatoze saistīta ar MHCI) ► Infekcijas (HIV-AIDS) ► Autoimūnās slimības (psoriāze, astma, reimatoīdais artrīts) ► Vēzis (Hodžkina limfoma) ► Jūtība pret zālēm.")
23
Vēzis Jūtība pret zālēm Monogēnās slimības Infekcijas
Autoimūnās slimības Vēzis Jūtība pret zālēm Monogēnās slimības Infekcijas

24
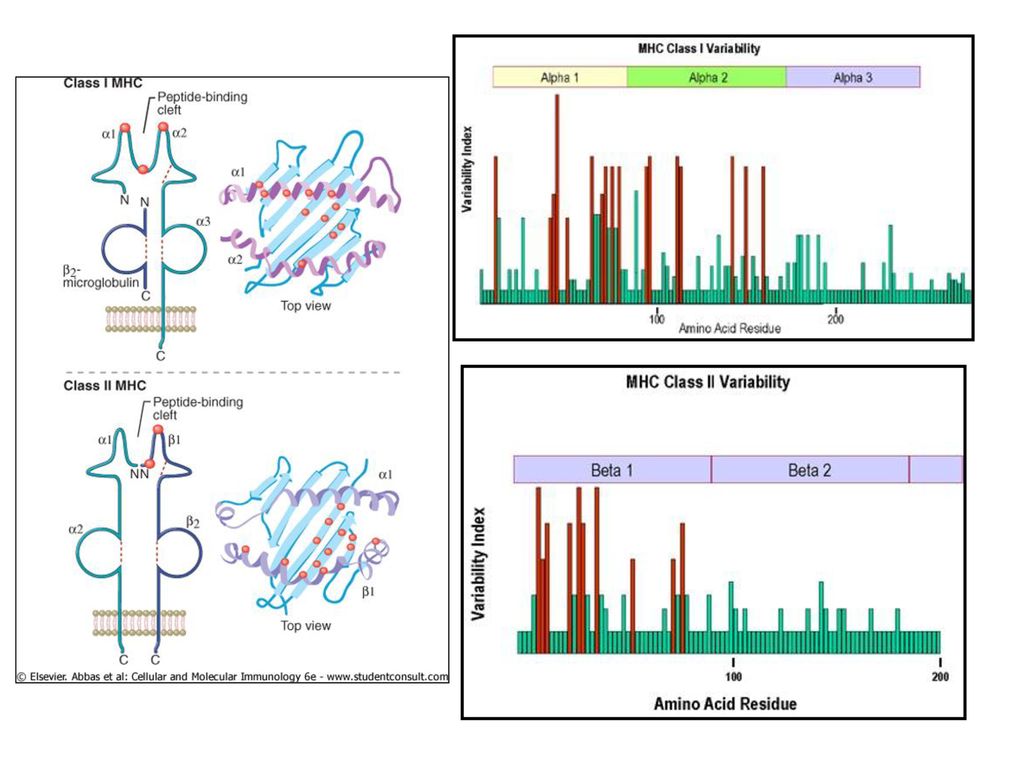
MHC I un II klases molekulu struktūra

25
Kopīgas īpašības visām I un II klases MHC molekulām
Transmembrānas glikoproteīni Ekspresētas uz šūnu virsmas Ļoti polimorfas Iesaistītas imunoregulācijā Katram proteīnam izšķir 4 rajonus - Variablais peptīda-saistīšanas rajons - Konstantais rajons - Transmembrānas rajons - Citoplazmiskais rajons

26
MHC I molekulas struktūra
2 polipeptīdu ķēdes: ▪ Garā a-ķēde (45kDa) ▪ Īsā ķēde (12kDa) – b2-mikroglobulīns Nekovalenti saistītais b2-mikroglobulīns palīdz stabilizēt MHC molekulas konformāciju. ► Citoplazmiskais rajons sastāv no 30 bāziskām aminoskābēm un satur fosforilēšanas saitus.
 ▪ Īsā ķēde (12kDa) – b2-mikroglobulīns. Nekovalenti saistītais. b2-mikroglobulīns palīdz. stabilizēt MHC molekulas. konformāciju. ► Citoplazmiskais rajons sastāv no. 30 bāziskām aminoskābēm un. satur fosforilēšanas saitus.")
27
MHC I molekulas struktūra
► Transmembrānas rajons – 25 hidrofobās aminoskābes. ► Ļoti konservatīvs a3 domēns pie kura saistās T-šūnu CD8 molekula ► Ļoti polimorfs peptīda saistīšanas rajons, kuru veido abi a1 un a2 domēni

28
MHC II molekulas struktūra
Divas aptuveni vienāda garuma polipetīdu ķēdes a un b. -ķēde ir kD b- ķēde ir kD ► Īss citoplazmiskais rajons, kas satur fosforilēšanas saitus. ►Transmembrānas rajons – 25 hidrofobās aminoskābes katrā a un b ķēdē.

29
MHC II molekulas struktūra
► Ļoti konservatīvs b2 domēns pie kura saistās T-šūnu CD4 molekula ► Ļoti polimorfs peptīda saistīšanas rajons, kuru veido abi a1 un b1 domēni

31
Kodē proteīnus, kas nav membrānu proteīni un
MHC I a ķēdi kodē HLA A HLA B HLA C HLA E HLA F HLA G MHC II a ķēdi kodē HLA DPA1 HLA DQA1 HLA DRA b ķēdi kodē HLA DPB1 HLA DQB1 HLA DRB1 HLA DRB3 HLA DRB4 HLA DRB5 Kodē proteīnus, kas nav membrānu proteīni un nav strukturāli līdzīgi MHC I un MHC II molekulām (TNF-α, komplementa sistēmas C2, C4 molekulas).
.")
32
Peptīda piesaiste pie MHC molekulas
ir fundamentāli atšķirīga no antigēna piesaistes pie B un T limfocītu antigēnu receptoriem MHC molekulas uzrāda plašu specifiskumu peptīdu piesaistē. Katra MHC molekula var piesaistīt daudz dažādus peptīdus. MHC molekulas peptīda saistīšanas vietā vienlaicīgi var piesaistīt tikai VIENU peptīdu. Peptīdiem, kurus saista MHC molekulas ir kopīgas strukturālās īpašības, kuras veicina šo piesaisti

33
☞ MHC I saista 8-11 aminoskābju garus peptīdus
☞ MHC II saista aminoskābju garus peptīdus (optimāli 12-16) ☞ Peptīdi saistās izstieptā konformācijā
 ☞ Peptīdi saistās izstieptā konformācijā.")
34
Peptīdi saistās pie MHC molekulām caur strukturāli
līdzīgām enkura aminoskābēm Katrs peptīds satur 1 vai 2 enkura aminoskābes. Enkura aminoskābes (zaļas) nav identiskas, bet ir līdzīgas: F un Y ir aromātiskas aminoskābes V, L un I ir lielas hidrofobas aminoskābes Ir attēloti peptīdi eluēti no divām dažādām I klases MHC molekulām.
 nav identiskas, bet ir līdzīgas: F un Y ir aromātiskas aminoskābes. V, L un I ir lielas hidrofobas aminoskābes. Ir attēloti peptīdi eluēti no divām dažādām. I klases MHC molekulām.")
35
Peptīdu piesaiste pie MHC molekulām
Pie MHC molekulas piesaistītais antigēna peptīds disociē no MHC molekulas ļoti lēni. Kinētikas disociācijas konstantes variē no stundām līdz pat vairākām dienām. Cilvēka MHC molekulas neatšķir svešus peptīdus no saviem peptīdiem. Kāpēc tad mēs parasti neattīstam imūnatbildi pret saviem proteīniem?

36
Pilnībā nokomplektētas MHC molekulas ir heterotrimēri
MHC I MHC II a- ķēde a- ķēde b- ķēde b2-mikroglobulīns piesaistīts antigēna peptīds piesaistīts antigēna peptīds MHC molekulas bez peptīda ir nestabilas Peptīda antigēna piesaiste stabilizē a-ķēdes un b2-mikroglobulīna mijiedarbību a-ķēdes un b2-mikroglobulīna mijiedarbība pastiprina peptīda antigēna piesaisti

37
T-šūnu antigēna receptors atpazīst
aminoskābes abos Antigēna peptīdā MHC molekulā Antigēna atpazīšanas specifiskums MHC restrikcija

38
Self MHC restriction Katra indivīda T-šūnas atpazīst svešus antigēnus TIKAI TAD, ja šie peptīdi ir piesaistīti un izstādīti uz paša indivīda MHC molekulām. Peter Doherty Rolf Zinkernagel g. atklājumi Nobeļa prēmija gadā

39
MHC molekulu funkcijas
▶ Svarīgas molekulas antigēna atpazīšanā un apstrādē. Inducē un stimulē imūnatbildi. I klases MHC molekulas ▶ Piesaista citoplazmā degradētos antigēnus (vīrusu proteīnus) ▶ Prezentē peptīdus CD8+ citotoksiskajiem T- limfocītiem II klases MHC molekulas ▶ Piesaista endocītiskajos pūslīšos degradētus peptīdus ▶ Prezentē peptīdus CD4+ T limfocītiem palīgiem (T helper)
 ▶ Prezentē peptīdus CD8+ citotoksiskajiem T- limfocītiem. II klases MHC molekulas. ▶ Piesaista endocītiskajos pūslīšos degradētus peptīdus. ▶ Prezentē peptīdus CD4+ T limfocītiem palīgiem (T helper)")
40
MHC molekulu ekspresija
MHC izplatība ir saistīta ar CD4+ un CD8+ T-šūnu funkcijām. ▶ CD8+ T-limfocīti nogalina šūnas, kuras ir inficētas ar intracelulāriem mikrobiem vai vīrusiem. Vīrusi var inficēt jebkuru kodolsaturošu šūnu. ▶ CD4+ T-šūnas palīdz makrofāgiem iznīcināt ekstracelulāros mikrobus un aktivēt B limfocītus antivielu producēšanai.

41
MHC molekulu ekspresija
MHC I klases molekulas: Ekspresētas uz visām kodolsaturošām šūnām. MHC II klases molekulas: Ekspresētas uz antigēnprezentējošo šūnu virsmas. Tiek regulētas ar IFN-g.

42
Katra kodolu saturoša šūna ir pārklāta ar vismaz
MHC I molekulām.

43
Secinājumi: MHC molekulu struktūra
▶ MHC I un II klases molekulām ir ▪ dažāda struktūra, ▪ dažāda izplatība šūnu veidos ▪ dažādas funkcijas ▶ Peptīdi, kurus saista MHC I un II klases molekulas ir atvasināti no dažādiem šūnu nodalījumiem un ir dažāda garuma ▶ Interferons-g regulē MHC molekulu ekspresiju

44
prezentēšana T limfocītiem
Antigēna apstrāde un prezentēšana T limfocītiem

45
Antigēnprezentējošās šūnas
T- šūnu funkcionēšana ir atkarīga no mijiedarbības ar citām šūnām.

46
Antigēnprezentējošās šūnas (APŠ)
Pārvērš proteīnu antigēnus par peptīdiem = antigēnu apstrāde. Eksponē peptīda-MHC kompleksu T-šūnu atpazīšanai. Ekspresē ko-stimulātorus, kuru ir svarīgi T šūnu aktivēšanai. Apoptoze

47
Dažādu šūnu tipi darbojās kā APŠ lai aktivētu
naivās un iepriekš diferencētās T-šūnas Naivās CD4+ Naivās CD8+ Diferenciētas CD4+ T-šūnas T-limfocītus (helper)
")
48
Profesionālās APŠ Vienīgās šūnas, kuru galvenās funkcijas ir antigēnu saistīšana un prezentēšana. Vienīgās APŠ, kuras ir spējīgas ierosināt T-šūnu atbildi.

49
Dendrītiskās šūnas ierosina T-šūnu atbildi
< 1% no šūnu populācijas 25% no epidermas (virsādas) virsmas
 virsmas.")
50
APŠ antigēn-prezentējošās funkcijas tiek pastiprinātas mikrobu produktu ietekmē
MHC un ko-stimulātoru ekspresija Uzlabojās antigēna prezentācijas efektivitāte Citokīnu produkcija APŠ Stimulē T šūnu atbildi Hemokīna receptora CCR7 ekspresija Stimulē APŠ migrāciju uz infekcijas vietām

51
Naivās T šūnas ekspresē CCR7 un migrē
hemokīni

52
Dendrītiskās un T šūnas in vitro

53
Kāpēc dendrītiskās šūnas ir visefektīvākās T-limfocīta primārās atbildes ierosināšanā?
DŠ stratēģiski izvietotas vietās, kur parasti mikrobi iekļūst mūsu organismā. DŠ ekspresē receptorus, kuri spēj piesaistīt mikrobus (mannozes un Toll-like receptori). DŠ pārsvarā migrē uz T-šūnu zonām limfmezglos, caur kurām cirkulē naivās T šūnas. Nobriedūšās DŠ ekspresē kostimulātorus, kuri ir nepieciešami naivo T-šūnu aktivēšanai.
. DŠ pārsvarā migrē uz T-šūnu zonām limfmezglos, caur. kurām cirkulē naivās T šūnas. Nobriedūšās DŠ ekspresē kostimulātorus, kuri ir nepieciešami naivo T-šūnu aktivēšanai.")
54
APŠ funkcijas Antigēna apstrāde Antigēna prezentēšana T-šūnām

55
fizikoķīmiskās īpašības
Antigēna īpašības, kuras atpazīst T-šūnas Antigēns fizikoķīmiskās īpašības T šūnas B šūnas un antivielas

56
Antigēna īpašības, kuras atpazīst T-šūnas
B šūnas Pārsvarā tikai peptīdus Peptīdus, proteīnus, nukleīnskābes, lipīdus, polisaharīdus, nelielas ķīmiskās molekulas Specifiski atpazīst peptīda aminoskābes sekvences Atpazīst lineāras aminoskābju sekvences un konformacionālos epitopus Atpazīst tikai tos antigēnus, kuri atrodas uz APŠ virsmas Piesaista kā šķīstošos tā arī uz šūnu virsmas eksponētos antigēnus

57
▶CD4+ T-šūnas atpazīst peptīda- MHC II klases molekulu kompleksus
Peptīdi atvasināti no ekstracelulāriem proteīniem, kas APŠ tiek internalizēti pūslīšos. ▶ CD8+ T-šūnas atpazīst peptīda- MHC I klases molekulas Peptīdi ir atvasināti no citozolā esošajiem, parasti endogēni sintezētiem proteīniem ▶Īpaša T-šūnu populācija atpazīst lipīdus ne MHC saistītā ceļā.

58
Antigēna apstrāde

59
Citozolā esošo antigēnu apstrāde
MHC I klases saistītai prezentācijai

60
Citozolā esošo antigēnu avots
Vīrusu un intracelulāro mikrobu produkti Kļūmīgi sintezēti vai aplami salocīti proteīni (brāķi) 30- 70% no sintezētiem proteīniem uzreiz tiek degradēti pēc to sintēzes. Tie ir defektīvie ribosomālie produkti – DRIPs.
 % no sintezētiem proteīniem uzreiz. tiek degradēti pēc to sintēzes. Tie ir defektīvie ribosomālie produkti – DRIPs.")
61
Citozolā esošo antigēnu avots
Vēžu šūnas var producēt proteīnu antigēnus no mutētiem vai pārekspresētiem gēniem Peptīdi, kuri nāk no fagosomās internalizētiem mikrobiem Producē listeriolizīnu, poras veidojošo toksīna proteīnu, kurš ļauj baktērijai izsprukt no pūslīša uz citozolu. Listeria monocytogenesis

62
Citozolā esošo proteīnu proteolītiskā degradācija
cytosolic proteins Proteolīze proteosomās Proteīnu degradācija sākās tikai pēc tam, kad pie proteīna ar kovalentām saitēm tiek pievienotas vairākas kopijas neliela polipeptīda- ubikvitīna. Poli-ubikvitīns kalpo kā specifisks signāls proteosomas atkarīgai degradācijai Proteasomas

63
Proteasoma (11S cap) 1500 kD 700 kD
Notver un novirza proteīnus uz kameru Proteīna degradācija 1500 kD IFN-g inducē jaunas proteasomas b-subvienības: LMP2, LMP7 un MECL-1, kuras aizvieto atbilstošas katalītiskās subvienības b1, b2 un b5. LMP- Low Molecular Wheight Protease complex

64
izmainīta katalītiskā aktivitāte substrāta specifiskums Producētie peptīdi ir no 6 līdz 30 aminoskābēm gari un parasti satur bāziskās vai hidrofobās aminoskābes C- terminālajā galā.

65
Peptīdu transportēšana no citozola uz endoplazmatisko tīklojumu
TAP – Transporter Associated with antigen Processing Peptīdu transports no citozola uz ET ir ATP atkarīgs. Optimāli tiek transportēti no 6 līdz 30 aminoskābēm gari peptīdi ar bāzisko vai hidrofobo C-galu.

66
TAP – Transporter Associated with antigen Processing
funkcijas TAP virza peptīdus no citozola uz ET lumenu. TAP ir platforma, kur savācās MHC I molekula piesaistot šāperonu tapazīnu.

67
Peptīda- MHC I klases kompleksa savākšanās ER
ER AminoPeptidase kalneksīns kalretikulīns

68
2 tapazīna un 2 MHC I klases molekulas saistās ar katru
Skats no ER lūmena N-terminālie palīgdomēni TAP1 un TAP2 transmembranālie domēni veido poru caur kuru tiek virzīts peptīds MHCI 2 tapazīna un 2 MHC I klases molekulas saistās ar katru N-terminālo palīgdomēnu MHCI MHCI Tapazīns ir transmembranālais glikoproteīns. Tapazīns piesaistās pie TAP un b2-mikroglobulīna. Kalpo kā tilts starp MHC I molekulu un TAP1/2 kompleksu. TMD -transmembrane domain; NBD-nucleotide binding domain

69
Peptīda piesaiste pie MHC molekulām notiek
PIRMS ekspresijas uz šūnas virsmas un ir būtisks komponents MHC molekulas biosintēzē un savākšanās. Peptīda piesaiste ir nepieciešama abām I un II klases MHC molekulām lai notiktu kompleksa stabila savākšanās un ekspresija uz šūnu virsmas.

70
saistītā antigēna apstrāde
MHC II klases saistītā antigēna apstrāde un prezentācija

71
Prezentē proteīnus, kuri ir ielauzušies šūnā no ārienes.
APŠ piesaista ekstracelulāros proteīna antigēnus un tos internalizē. C-tipa lektīnus Makrofāgi ekspresē receptorus antivielu Fc daļai un receptorus C3b komplementa proteīnam B šūnas ekspresē virsmas imunoglobulīnu APŠ atpazīst struktūras, kuras ir raksturīgas mikrobu patogēniem (lipīdi un ogļhidrāti) un nav atrodamas zīdītāju šūnās.
 un nav atrodamas. zīdītāju šūnās.")
72
MHC II klases saistītā antigēnu peptīdu
prezentācija

73
MHC II molekulas biosintēze un transports
uz endosomām MHC II klases a un b ķēdes sintezējās un saistās viena ar otru ET. Viņu foldings un savākšanās notiek ar ET šaperonu proteīnu (kalneksīns) palīdzību. a un b ķēdes komplekss ir nestabils un tāpēc tiek stabilizēts ar trešo proteīnu – invarianto (nemainīgo) ķēdi (Ii). Ii ir trimērs, kas sastāv no trim 30kD subvienībām.
 palīdzību. a un b ķēdes komplekss ir nestabils un tāpēc tiek. stabilizēts ar trešo proteīnu – invarianto (nemainīgo) ķēdi (Ii). Ii ir trimērs, kas sastāv no trim 30kD subvienībām.")
74
Invariantās ķēdes proteīns
► Katra Ii subvienība saista vienu ab heterodimēru un nobloķē MHC II molekulas peptīda saistīšanas vietu. ► Ii veicina MHC II molekulas foldingu un savākšanos un virza jaunizveidoto MHC II molekulu uz vēlajām endosomām un lizosomām.

75
Makrofāgos un B šūnās daļu no vēlajām endosomām, kuras ir
Early endosome with antigen HLA-DM Makrofāgos un B šūnās daļu no vēlajām endosomām, kuras ir bagātas ar MHC II klases molekulām sauc par MHC II klases molekulu nodalījumu vai MIIC (compartment) Enzīmi antigēna degradācijai Ii MIIC MHC II molekulas Cilvēka leikocītu antigēns DM (HLA-DM)
 Enzīmi antigēna. degradācijai. Ii. MIIC. MHC II. molekulas. Cilvēka leikocītu. antigēns DM (HLA-DM)")
76
CLIP aizvākšana ir nepieciešama pirms peptīda antigēna saistīšanas.
MIIC iekšienē Catepsin S 24 aa CLIP Class II-associated Invariant Peptide CLIP aizvākšana ir nepieciešama pirms peptīda antigēna saistīšanas. HLA-DM palīdz aizvākt CLIP.

77
HLA-DM Gēns ir kodēts MHC II lokusā
Struktūra ir līdzīga MHC II molekulai Kolokalizēta ar MHC II molekulām endosomās Nav polimorfa Nesaistās ar Ii peptīdu Netiek ekspresēta uz šūnas virsmas DM

78
HLA-DM molekulas funkcijas
▶ Izraisa CLIP disociāciju no MHC II peptīda saistīšanas gropes ▶ Stabilizē un pasargā no degradācijas tukšas MHC II ▶ Veicina antigēna peptīda piesaisti pie MHC II

79
Peptīda piesaistīšana pie MHC II molekulas to stabilizē.
Stabils peptīda-MHC II komplekss tiek nogādāts uz APŠ virsmas. Tiek izstādīts atpazīšanai CD4+ T-šūnām.

80
MHC - peptīda kompleksa veidošanās procesā NAV
mehānisma, kas ļautu atšķirt savus proteīnus no svešiem proteīniem. Viena šūna parasti ir apkārusies ar MHC molekulām. Lielāka daļa no MHC molekulām ir MHC I klases molekulas, kas saistītas ar saviem proteīniem. Kā T-šūna spēj atpazīt un būt aktivēta ar jebkuru svešu antigēnu?

81
T-šūnas ir ārkārtīgi jūtīgas.
< 0.1% no kopēja skaita II klases MHC molekulu spēj iniciēt specifisko T-šūnu atbildi. Kāpēc parasti mēs neveidojām imūnatbildi pret saviem proteīniem? T šūnas specifiskas pret self peptīda-MHC kompleksiem tiek nogalinātas vai inaktivētas.

82
Lipīdu antigēnu prezentācija
Lipīdu antigēni tiek prezentēti T-šūnām caur šūnu virsmas molekulām -CD1 (cluster of differentiation) CD1 tiek ekspresētas uz dažādām APŠ un epitēlijā. APŠ ekspresē uz savas virsmas dažādas formas CD1 molekulas. Katra CD1 molekula piesaista noteikta veida lipīda antigēnu (lipopeptīdu vai glikolipīdu). CD1 molekulas veido antigēna saistīšanas gropi līdzīgi kā MHC molekulas. Aminoskābes antigēna saistīšanas gropē ir hidrofobākas nekā tādas MHC molekulā. Līdzīgi kā proteīna antigēni, arī lipīdu antigēni tiek sadalīti un prezentēti kā fragmenti.
 CD1 tiek ekspresētas uz dažādām APŠ un epitēlijā. APŠ ekspresē uz savas virsmas dažādas formas CD1 molekulas. Katra CD1 molekula piesaista noteikta veida lipīda antigēnu (lipopeptīdu vai glikolipīdu). CD1 molekulas veido antigēna saistīšanas gropi līdzīgi kā MHC molekulas. Aminoskābes antigēna saistīšanas gropē ir hidrofobākas nekā. tādas MHC molekulā. Līdzīgi kā proteīna antigēni, arī lipīdu antigēni tiek sadalīti un prezentēti kā fragmenti.")
83
CD1 prezentētus lipīdu antigēnus atpazīst vairākas šūnas
CD4+ T-šūnas CD8+ T-šūnas CD4-CD8- T- šūnas NK-T-šūnas (T-šūnas, kuras ekspresē NK šūnu marķierus)
")
84
APŠ antigēns T-limfocīts

Παρόμοιες παρουσιάσεις



















 4.2.1 Nguyên tắc hoạt động 4.1.2 Thời gian làm.>")