Κατέβασμα παρουσίασης
2
Μια μεταγραφική μονάδα μεταγράφεται σε ένα ενιαίο RNA.

3
Η μεταγραφή αποτελεί το πρώτο βήμα στη γονιδιακή έκφραση και το κυριότερο στάδιο ελέγχου της. Πώς η RNA πολυμεράση εντοπίζει τους υποκινητές πάνω στο DNA; Πώς οι ρυθμιστικές πρωτεΐνες αλληλεπιδρούν με την RNA πολυμεράση (και μεταξύ τους) για να ενεργοποιήσουν ή να καταστείλουν συγκεκριμένα βήματα της μεταγραφής;
 για να ενεργοποιήσουν ή να καταστείλουν συγκεκριμένα βήματα της μεταγραφής;.")
4
Εισαγωγικά περί πολυμερασών… Οι RNA πολυμεράσες των βακτηρίων, των ευκαρυωτών και των αρχαίων είναι μεγάλα ένζυμα με πολλές υπομονάδες που χρειάζονται τη βοήθεια επιπρόσθετων παραγόντων για να αναγνωρίσουν συγκεκριμένα γονίδια Παρόλο που οι διάφορες RNA πολυμεράσες διαφέρουν από πολλές απόψεις, οι καταλυτικός πυρήνας των ενζύμων εμφανίζει αξιοσημείωτη ομοιότητα και στις τρεις ενότητες ζωής Επομένως, τα συμπεράσματα από τη μελέτη μια πολυμεράσης μας είναι χρήσιμα και για τις υπόλοιπες

5
Η κρυσταλλική δομή της RNA πολυμεράσης στην E. coli δεν έχει προσδιοριστεί Πληροφορίες για την κρυσταλλική δομή της πολυμεράσης έχουμε από Thermus aquaticus και T. thermophilus Λόγω υψηλού βαθμού ομολογίας μπορούμε και βγάζουμε συμπεράσματα για άλλες πολυμεράσες

6
Συμβάσεις αποτύπωσης της πορείας

7
Οι RNA πολυμεράσες των βακτηρίων είναι μεγάλες πρωτεΐνες με πολλές υπομονάδες 459 kDa

8
Η δοκιμή ενεργότητας της RNA πολυμεράσης DNA Mg 2+ Τρία μη ραδιενεργά ΝΤΡ Ένα ραδιενεργό ΝΤΡ (σημασμένο με 3 Η ή 14 C) Τερματισμός της αντίδρασης με τριχλωροοξικό οξύ (TCA) που καθιστά αδιάλυτο το νεοσχηματισμένο RNA Η ποσότητα της ραδιενέργειας στο ίζημα είναι ανάλογη της ποσότητας RNA που έχει σχηματιστεί
 Τερματισμός της αντίδρασης με τριχλωροοξικό οξύ (TCA) που καθιστά αδιάλυτο το νεοσχηματισμένο RNA Η ποσότητα της ραδιενέργειας στο ίζημα είναι ανάλογη της ποσότητας RNA που έχει σχηματιστεί")
9
H T7 RNA πολυμέραση Μία πολυπεπτιδική υπομονάδα 99 kDa Τριτοταγή δομή που μοιάζει με την Klenow της βακτηριακής RNA pol Συνθέτει RNA με μεγάλη αποτελεσματικότητα

10
Γιατί η RNA pol της E. coli είναι τόσο μεγάλη; Στο βακτήριο η RNA pol πρέπει να αναγνωρίσει ~4300 γονίδια με ~1000 διαφορετικές θέσεις πρόσδεσης Η βακτηριακή RNA pol αποκρίνεται σε μεγάλο αριθμό ρυθμιστικών πρωτεϊνών που τροποποιούν την ικανότητα της πολυμεράσης να αναγνωρίζει μια θέση πρόσδεσης

11
Η αντίδραση της μεταγραφής συντελείται σε τρία στάδια, τα εξής τέσσερα: Αναγνώριση της μήτρας Κλειστό σύμπλοκο Ανοικτό σύμπλοκο Έναρξη Ολοκληρώνεται εάν και εφόσον το ένζυμο καταφέρει να μετακινηθεί από την αρχική του θέση. Επιμήκυνση Τερματισμός

12
Το ολοένζυμο της RNA pol των βακτηρίων αποτελείται από ένα κεντρικό ένζυμο και τον παράγοντα σ Η κεντρική πολυμεράση μπορεί να διαχωριστεί από τον παράγοντα σ με χρωματογραφία ανταλλαγής ιόντων σε στήλη φωσφοκυτταρίνης

13
Η κεντρική πολυμέραση α2ββ’ είναι σε θέση να συνθέσει RNA χρησιμοποιώντας ως μήτρα μονόκλωνο DNA ή δίκλωνο DNA που φέρει εγκοπή Δεν μπορεί να χρησιμοποιήσει ως μήτρα ανέπαφο δίκλωνο μόριο DNA Ούτε ο παράγοντας σ ούτε η υπομονάδα ω είναι απαραίτητα για το σχηματισμό φωσφοδιεστερικού δεσμού Καμία από τις υπομονάδες α ή β ή β’ δεν έχει την ικανότητα να καταλύσει από μόνη της τη σύνθεση του RNA

14
Η σύνδεση της RNA πολυμεράσης στον υποκινητή

15
Ο κύκλος του παράγοντα σ

16
Ο σχηματισμός του κλειστού δυαδικού συμπλόκου είναι αντιστρεπτός και χαρακτηρίζεται από μια σταθερά ισορροπίας Κ Β. Οι τιμές της Κ Β ποικίλλουν ευρέως ανάλογα με τον υποκινητή. Η μετατροπή του κλειστού συμπλόκου σε ανοικτό περιγράφεται από μια κινητική σταθερά k 2 και είναι γρήγορη. Η ενσωμάτωση των δύο πρώτων νουκλεοτιδίων και η δημιουργία ενός φωσφοδιεστερικού δεσμού (δημιουργία τριαδικού συμπλόκου) περιγράφεται από μια ακόμη πιο γρήγορη κινητική σταθερά k i. Το ένζυμο δεν μετακινείται αν δεν δημιουργηθεί μια αλυσίδα RNA με μήκος 9 βάσεων. Μετά την έναρξη ο παράγοντας σ μπορεί να «εκκαθαριστεί»: αυτή η διαδικασία διαρκεί 1-2 sec. Η RNA πολυμεράση διέρχεται από αρκετά στάδια πριν τη φάση επιμήκυνσης. Ένα κλειστό δυαδικό σύμπλοκο μετατρέπεται σε μια ανοικτή μορφή και μετά σε ένα τριαδικό σύμπλοκο
 περιγράφεται από μια ακόμη πιο γρήγορη κινητική σταθερά k i. Το ένζυμο δεν μετακινείται αν δεν δημιουργηθεί μια αλυσίδα RNA με μήκος 9 βάσεων. Μετά την έναρξη ο παράγοντας σ μπορεί να «εκκαθαριστεί»: αυτή η διαδικασία διαρκεί 1-2 sec. Η RNA πολυμεράση διέρχεται από αρκετά στάδια πριν τη φάση επιμήκυνσης. Ένα κλειστό δυαδικό σύμπλοκο μετατρέπεται σε μια ανοικτή μορφή και μετά σε ένα τριαδικό σύμπλοκο.")
17
Ο παράγοντας σίγμα και το κεντρικό ένζυμο ανακυκλώνονται σε διαφορετικές φάσεις κατά τη μεταγραφή.

18
Η αναγνώριση του υποκινητή εξαρτάται από πρότυπες αλληλουχίες Ο David Prinbow χρησιμοποίησε τη μέθοδο της προστασίας του DNA για την ανάλυση υποκινητών σ 70 διαφόρων γονιδίων

19
Μετά από ανάλυση >300 υποκινητών προέκυψαν συναινετικές (ή πρότυπες) αλληλουχίες για τον σ 70
 αλληλουχίες για τον σ 70")
20
T 82 T 84 G 78 A 65 C 54 A 45 T 80 A 95 T 45 A 60 A 50 T 96 Ένας τυπικός υποκινητής αποτελείται από τρία στοιχεία, τα εξής τέσσερα: Την πρότυπη αλληλουχία στη θέση -10 Την πρότυπη αλληλουχία στη θέση -35 Την απόσταση ανάμεσα στις θέσεις -35 και -10 Το σημείο έναρξης

21
Η αλληλουχία -35 χρησιμοποιείται για την αρχική αναγνώριση και η αλληλουχία -10 για την αντίδραση τήξης, που μετατρέπει ένα κλειστό σύμπλοκο σε ανοικτό. Οι μειορυθμικές μεταλλάξεις στην –35 μειώνουν το ρυθμό σχηματισμού του κλειστού συμπλόκου. Οι μειορυθμικές μεταλλάξεις στην –10 δεν επηρεάζουν τον αρχικό σχηματισμό του συμπλόκου, όμως επιβραδύνουν τη μετατροπή του σε ανοικτό. Άρα: Η -35 παρέχει το σήμα αναγνώρισης για την RNApol Η -10 επιτρέπει τη μετατροπή από κλειστή σε ανοικτή μορφή (Τ/Α)
.")
22
Οι υποκινητές σ 70 των ριβοσωμικών RNA έχουν μια περιοχή που ονομάζεται ανοδικό στοιχείο (up element) Το στοιχείο αυτό αλληλεπιδρά με την υπομονάδα α του ολοενζύμου Αυτή η αλληλεπίδραση ευθύνεται για την αύξηση της ισχύος του υποκινητή που εξηγεί τον υψηλό ρυθμό σύνθεσης του rRNA
 Το στοιχείο αυτό αλληλεπιδρά με την υπομονάδα α του ολοενζύμου Αυτή η αλληλεπίδραση ευθύνεται για την αύξηση της ισχύος του υποκινητή που εξηγεί τον υψηλό ρυθμό σύνθεσης του rRNA")
23
Η ιχνηλάτηση DNA αποκαλύπτει τη μετάβαση από το κλειστό στο ανοικτό σύμπλοκο

24
Όταν το σύμπλοκο RNA πολυμέρασης σ 70 με τον υποκινητή σ 70 γίνει σε χαμηλή θερμοκρασία ( 0 °C), η οποία αποτρέπει τις αλλαγές διαμόρφωσης του συμπλόκου, η προστατευόμενη περιοχή του DNA εκτείνεται μεταξύ των θέσεων -55 και -10: ΚΛΕΙΣΤΟ ΣΥΜΠΛΟΚΟ Όταν το σύμπλοκο σχηματίζεται στους 37 °C, η προστατευόμενη περιοχή είναι μεγαλύτερη και εκτείνεται μεταξύ των θέσεων -55 και +20: ΑΝΟΙΚΤΟ ΣΥΜΠΛΟΚΟ
, η οποία αποτρέπει τις αλλαγές διαμόρφωσης του συμπλόκου, η προστατευόμενη περιοχή του DNA εκτείνεται μεταξύ των θέσεων -55 και -10: ΚΛΕΙΣΤΟ ΣΥΜΠΛΟΚΟ Όταν το σύμπλοκο σχηματίζεται στους 37 °C, η προστατευόμενη περιοχή είναι μεγαλύτερη και εκτείνεται μεταξύ των θέσεων -55 και +20: ΑΝΟΙΚΤΟ ΣΥΜΠΛΟΚΟ")
25
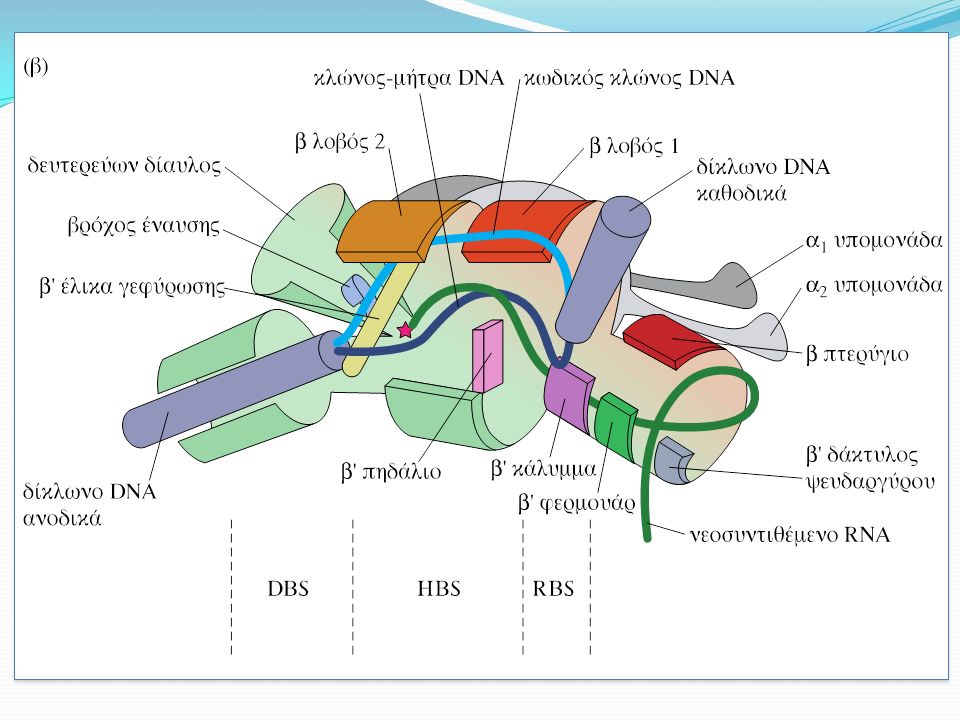
Οι κρυσταλλικές δομές της RNA pol υποδεικνύουν το μηχανισμό λειτουργίας του ενζύμου Το 1999 ο Seth Darst προσδιόρισε την κρυσταλλική δομή της κεντρικής RNA pol του Thermus aquaticus Η πρωτεΐνη αυτή έχει πέντε υπομονάδες: α2ββ’ω Η υπομονάδα ω συμβάλλει στη συναρμολόγηση του ενζύμου 1. Σχήμα δαγκάνας 2. Οι υπομονάδες α σχηματίζουν διμερές. Η μία α με τη β και η άλλη α με τη β’. Καμία α δεν έχει πρόσβαση στο χώρο κατάλυσης 3. Οι β και β’ συνιστούν το 60% της πολυμεράσης. Η β’ παρέχει το συντηρημένο μοτίβο NADFDGD που είναι απαραίτητο για την κατάλυση

26
Το 2002 ο Shigeyuki Yokoyama προσδιόρισαν την κρυσταλλική δομή του Thermus thermophilus

27
Οι κρύσταλλοι ολοενζύμου πολυμεράσης με συνθετικό DNA παρέχουν πληροφορίες για τις αλληλεπιδράσεις μεταξύ κεντρικού ενζύμου, παράγοντα σ και DNA

28
Όλες οι επαφές της αλληλουχίας των πλαισίων -10 και -35 με την RNA πολυμεράση λαμβάνουν χώρα μέσω αλληλεπιδράσεων με την υπομονάδα σ

29
Οι παράγοντες σ δημιουργούν άμεσες επαφές με το DNA Η υπομονάδα σ 70 έχει 4 συντηρημένες δομικές επικράτειες, καθεμία των οποίων μπορεί να υποδιαιρεθεί σε μικρότερες ιδιαίτερα συντηρημένες

30
Άμεση απόδειξη της αλληλεπίδρασης ανάμεσα στις επικράτειες του παράγονται σ και τις περιοχές του υποκινητή προέρχεται από ορισμένες μεταλλάξεις του σ που αντισταθμίζουν μειορυθμικές μεταλλάξεις στις περιοχές –35 και –10 Τα αμινοξέα στην α-έλικα 2.4 του σ 70 δημιουργούν ειδικές επαφές με βάσεις στην περιοχή -10 της κωδικής αλυσίδας του υποκινητή

31
Το Ν-τελικό άκρο του σίγμα εμποδίζει τις επικράτειες πρόσδεσης στο DNA να προσδεθούν σε αυτό. Όταν σχηματίζεται ένα ανοικτό σύμπλοκο, το Ν-τελικό άκρο μετακινείται 20 Ǻ μακριά και οι δύο επικράτειες πρόσδεσης στο DNA απομακρύνονται η μία από την άλλη κατά 15 Ǻ Αν το Ν-τελικό άκρο του παράγοντα σ απαλειφθεί, τότε μπορεί να συνδεθεί με το DNA. Έξτρα μεταλλάξεις μπορούν να προσδιορίσουν τα ακριβή σημεία επαφής του σ με τις περιοχές -35 και -10.

32
Το ολοένζυμο της RNApol-σ 70 διέρχεται αρκετούς κύκλους διακοπτόμενης έναρξης πριν διαφύγει από τον υποκινητή

33
Μετακινείται ή όχι η πολυμεράση κατά τη διακοπτόμενη έναρξη; Τρία μοντέλα που προσπαθούν να συμβιβάσουν αντιφατικά πειραματικά δεδομένα

35
Το σύμπλοκο επιμήκυνσης της μεταγραφής Κεντρική RNA pol, DNA μήτρα, νέο RNA Μόρια ΝΤΡ διέρχονται από τον δευτερεύοντα δίαυλο για να φτάσουν το καταλυτικό κέντρο. Περιορισμός ταχύτητας επιμήκυνσης: 30 νουκλεοτίδια / sec Η 3’-υδροξυλομάδα πραγματοποιεί πυρηνόφιλη προσβολή στην α-φωσφορυλομάδα του σωστού ΝΤΡ για το σχηματισμό του 5’-3’ φωσφοδιεστερικού δεσμού Τα εισερχόμενα ΝΤΡ παρέχουν αρκετή ενέργεια για τη σύνθεση του φωσφοδιεστερικού δεσμού

36
Το τριαδικό σύμπλοκο επιμήκυνσης είναι ~35 bp μικρότερο από το σύμπλοκο έναρξης (DNase protection experiments) Περίπου 14 bp εντός της προστατευόμενης περιοχής αποδιατάσσονται σχηματίζοντας της μεταγραφική θηλιά, η οποία μένει σταθερή κατά την επιμήκυνση Τα πρώτα 8 νουκλεοτίδια της θηλιάς είναι ζευγαρωμένα με την αλυσίδα RNA
 Περίπου 14 bp εντός της προστατευόμενης περιοχής αποδιατάσσονται σχηματίζοντας της μεταγραφική θηλιά, η οποία μένει σταθερή κατά την επιμήκυνση Τα πρώτα 8 νουκλεοτίδια της θηλιάς είναι ζευγαρωμένα με την αλυσίδα RNA")
37
Η κρυσταλλική δομή του συμπλόκου επιμήκυνσης του Thermus thermophilus

39
Απομάκρυνση λαθών Όταν υπάρχει αταίριαστο ζευγάρι DNA- RNA η πολυμεράση σταματά και μετά οπισθοδρομεί μια θέση Η αστάθεια του υβριδίου DNA-RNA επάγει την οπισθοδρόμηση Ένα οπισθοδρομημένο σύμπλοκο διασώζεται με τη σχάση και απελευθέρωση του 3’-άκρου Ο παράγοντας GreA επάγει την απομάκρυση 2-3 νουκλεοτιδίων Ο παράγοντας GreB απομακρύνει τμήματα έως 18 νουκεοτίδια

41
Η βακτηριακή RNA pol τερματίζει τη μεταγραφή σε διακριτές θέσεις

42
Οι αλληλουχίες DNA που απαιτούνται για τον τερματισμό εντοπίζονται πριν την αλληλουχία τερματισμού. Ίσως είναι αναγκαίος ο σχηματισμός μιας φουρκέτας στο RNA. Το 3’ άκρο του μορίου μπορεί να έχει δημιουργηθεί με αποκοπή του πρωτογενούς μεταγράφου και επομένως να μην αντιστοιχεί στην πραγματική θέση τερματισμού της μεταγραφής. Ο αντιτερματισμός έχει σαν αποτέλεσμα την αναγνωστική διέλευση. Άρα, ο τερματισμός μπορεί να αποτελέσει μια ευκαιρία ελέγχου γονιδιακής έκφρασης

43
Υπάρχουν δύο τύποι αλληλουχιών τερματισμού στην E. coli

44
Οι ενδογενείς αλληλουχίες τερματισμού περιλαμβάνουν παλινδρομικές περιοχές οι οποίες σχηματίζουν φουρκέτες (δομές στελέχους-βρόχου) με ποικίλο μήκος, από 7-20 bp. Η δομή στελέχους-βρόχου περιλαμβάνει μια περιοχή πλούσια σε G- C και ακολουθείται από μια σειρά καταλοίπων U. Υπάρχουν ~1100 τέτοιες αλληλουχίες στο γονιδίωμα της E. coli => τα μισά γονίδια φέρουν ενδογενείς τερματιστές. Ποια είναι η επίδραση της φουρκέτας; Πιθανώς μια παύση (~60sec) που αποσταθεροποιεί ακόμα περισσότερο τα ζευγάρια Α=U. Η αποτελεσματικότητα τερματισμού in vitro είναι από 2-90%. Συνεπώς η φουρκέτα και τα U είναι αναγκαία αλλά όχι επαρκή.
 που αποσταθεροποιεί ακόμα περισσότερο τα ζευγάρια Α=U. Η αποτελεσματικότητα τερματισμού in vitro είναι από 2-90%. Συνεπώς η φουρκέτα και τα U είναι αναγκαία αλλά όχι επαρκή..")
45
Πώς λειτουργεί ο παράγοντας ρ (Rho) Ο παράγοντας ρ είναι μια απαραίτητη πρωτεΐνη στην E. coli που λειτουργεί αποκλειστικά στο στάδιο του τερματισμού Αποτελείται από έξι όμοιες υπομονάδες και έχει μοριακό βάρος ~275 kD. Κάθε υπομονάδα έχει μια επικράτεια πρόσδεσης στο RNA και μια επικράτεια υδρόλυσης του ΑΤΡ.

46
Ένας Rho-εξαρτώμενος τερματιστής έχει αλληλουχία πλούσια σε C και φτωχή σε G, και εντοπίζεται πριν από την ακριβή θέση (ή θέσεις) τερματισμού. Η αλληλουχία παρουσιάζεται με τη μορφή του RNA. Αντιπροσωπεύει το 3΄ άκρο του RNA.

47
Ο παράγοντας Rho «καταδιώκει» την RNA πολυμεράση κατά μήκος του RNA και μπορεί να προκαλέσει τερματισμό όταν προλάβει το στάσιμο ένζυμο σε μια Rho-εξαρτώμενη αλληλουχία τερματισμού.

48
Πολικότητα (Polarity): σε μερικές περιπτώσεις, μια ανερμηνεύσιμη μετάλλαξη σε ένα γονίδιο μιας μεταγραφικής μονάδας εμποδίζει τη μεταγραφή των επόμενων γονιδίων της ίδιας μονάδας. Πώς εξηγείται;;

49
Η δράση του παράγοντα Rho μπορεί να συνδέσει τη μεταγραφή με τη μετάφραση όταν η Rho-εξαρτώμενη αλληλουχία τερματισμού βρίσκεται λίγο μετά από μια ανερμηνεύσιμη μετάλλαξη.

50
Tropp, 12.1 έως 12.4


















 Κύτταρο Β ) Δομές DNA - RNA Παρουσίαση Βιολογίας.>")





