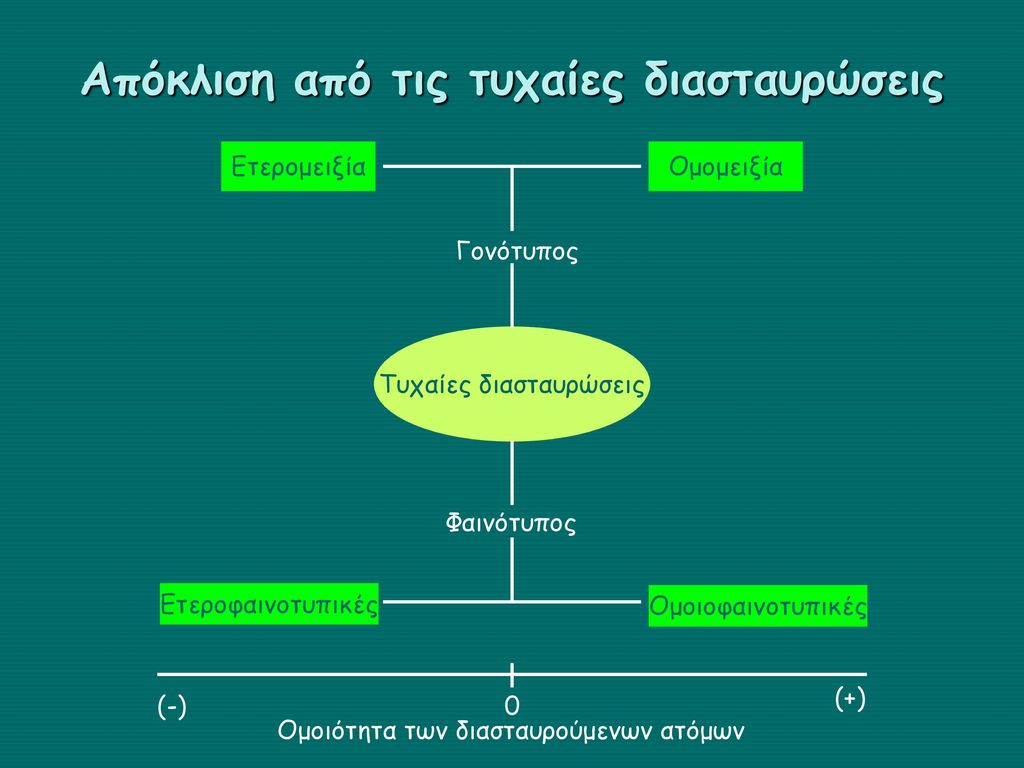
Απόκλιση από τις τυχαίες διασταυρώσεις Γονότυπος Φαινότυπος Ομομειξία Ομοιοφαινοτυπικές Ετερομειξία Ετεροφαινοτυπικές Τυχαίες διασταυρώσεις (-) (+) Ομοιότητα των διασταυρούμενων ατόμων
Ετερομειξία ≠ Ομομειξία Συζεύξεις μεταξύ μη συγγενών Ανάλογα με τον βαθμό γενετικής διαφοροποίησης των διασταυρούμενων ατόμων διακρίνεται σε: Αμιγή αναπαραγωγή και Διασταύρωση
Αμιγής αναπαραγωγή Ήπιο σύστημα ετερομειξίας Σύζευξη μεταξύ ατόμων τα οποία είναι μεταξύ τους λιγότερο συγγενή από ότι κατά μέσο όρο δύο τυχαία άτομα του πληθυσμού Για περιορισμένο μέγεθος πληθυσμού, δεν αυξάνει την ετεροζυγωτία του πληθυσμού αλλά επιβραδύνει τον ρυθμό μείωσης της ετεροζυγωτίας Σε συνδυασμό με την επιλογή γεννητόρων αποτελεί τη σπουδαιότερη μέθοδο βελτίωσης των ζώων
Διασταύρωση δύο πληθυσμών ή σειρών Ακραίο σύστημα ετερομειξίας Είναι η σύζευξη μεταξύ ατόμων τα οποία ανήκουν σε διαφορετικές αναπαραγωγικές κοινότητες, όπως φυλές, πληθυσμοί, υποπληθυσμοί, αμιγείς σειρές ή ακόμα και είδη Έχει τα ακριβώς αντίθετα αποτελέσματα από την ομομειξία Δηλαδή, αυξάνει το βαθμό ετεροζυγωτίας των απογόνων και αναβαθμίζει τους χαρακτήρες αυτούς που υποβαθμίζει η ομομειξία (γονιμότητα, βιωσιμότητα, ανθεκτικότητα)
Διασταύρωση Αύξηση της ετεροζυγωτίας p, q P, Q Π1 Π2 H1 = 2pq H2 = 2PQ H = ½ (H1+H2) = pq + PQ p + q = P + Q p - P = Q – q = y P = p – y , Q = q + y H = 2pq + (p-q)y – y2 Έστω ο γενετικός τόπος Α(Α,α) και δύο πληθυσμοί Π1 και Π2. Αν οι συχνότητες των αλληλομόρφων Α και α, διαφέρουν στους δύο πληθυσμούς και είναι p, q στον Π1 και P, Q στον Π2, τότε η συχνότητα των ετεροζυγωτών ατόμων είναι 2pq στον Π1 και 2PQ στον Π2. Δηλαδή η μέση ετεροζυγωτία των δύο πληθυσμών είναι ο μέσος όρος της ετεροζυγωτίας των δύο πληθυσμών, ήτοι H = pq + PQ. Επειδή δε, p+q = 1 όπως και P+Q=1, έπεται ότι p+q = P+Q p-P = Q-q. Αν ορίσω y την ποσότητα αυτή, τότε p-P=y P=p-y και Q-q=y Q=q+y. Αντικαθιστώντας τα P και Q, προκύπτει ότι H = pq+(p-y)(q+y) = pq+pq+py-qy-y2 = 2pq + y(p-q) – y2 Αν ωστόσο διασταυρωθούν τα άτομα του πληθυσμού Π1 με αυτά του Π2 τότε η ετεροζυγωτία της F1 γενιάς δίνεται από το άθροισμα pQ+Pq. Με αντικατάσταση πάλι προκύπτει ότι η ετεροζυγωτία της F1 είναι μεγαλύτερη από την μέση ετεροζυγωτία των δύο πληθυσμών κατά την θετική ποσότητα y2 . Επομένως η διασταύρωση προκαλεί αύξηση της ετεροζυγωτίας στην γενιά F1. Οι συχνότητες των αλληλομόρφων Α και α στην γενιά F1 είναι οι μέσες τιμές των αλληλομόρφων στους δύο αρχικούς πληθυσμούς. Δηλαδή, η νέα συχνότητα του αλληλομόρφου Α είναι ίση με ½(p+P) = ½ (p+p-y) = ½ (2p-y) = p – ½ y. Αντίστοιχα προκύπτει ότι η συχνότητα του αλληλομόρφου α είναι q + ½ y. Στην F2, ο πληθυσμός βρίσκεται σε ισορροπία H-W και άρα η ετεροζυγωτία της γενιάς F2 παραμένει σταθερή και στις επόμενες γενιές, υπό την προϋπόθεση ότι δεν ασκούνται επιλεκτικές πιέσεις. Η ετεροζυγωτία της γενιάς F2 είναι ίση με 2(p- ½ y)(q + ½ y) = (2p-y)(q+ ½ y) = 2pq + py –qy – ½ y2 = 2pq + y(p-q)- ½ y2. Η ετεροζυγωτία της γενιάς F2 είναι μειωμένη κατά ½ y2, σε σχέση με την ετεροζυγωτία της F1. Άρα η μέγιστη ετεροζυγωτία επιτυγχάνεται στην F1. HF1 = pQ + Pq = … = 2pq + (p-q)y HF1 – H = y2 HF2 = 2p’q’ = 2 x ½(p+P) x ½(q+Q) =… = 2pq + (p-q)y –½y2
Επιλογή Έχουμε επιλογή όταν άτομα με διαφορετικούς γονοτύπους που ζουν κάτω από ορισμένες συνθήκες αφήνουν συστηματικά διαφορετικό αριθμό απογόνων Ο συντελεστής προσαρμογής w ενός γονοτύπου εκφράζει την ποσοτική συνεισφορά του γονοτύπου σε απογόνους στην επόμενη γενιά Συντελεστής επιλογής s (w= 1-s) Ο συντελεστής προσαρμογής συνήθως δίνεται σε σχετικές τιμές. Εκφράζεται ως το πηλίκο του συντελεστή προσαρμογής προς τον συντελεστή προσαρμογής του γονοτύπου με την υψηλότερη τιμή. Δηλαδή ο γονότυπος με το μέγιστο συντελεστή προσαρμογής παίρνει την τιμή 1 και οι άλλοι γονότυποι εκφράζονται ως αποκλίσεις από την τιμή αυτή Ένα άλλο μέτρο της ισχύος της επιλογής είναι ο συντελεστής επιλογής
Μετά την επιλογή Δq= pq[p(w12-w11) + q(w22-w12)]/w A : a p’ = (p2w11 + pqw12)/w και q’ = (q2w22 + pqw12)/w Δq = q’ – q Δq = (q2w22 + pqw12)/w – qw/w = = [q2w22 + pqw12 – q(p2w11+2pqw12+q2w22)]/w = = [q2w22 + pqw12 – qp2w11-2pq2w12-q3w22] /w = = [q2w22(1-q)+pqw12(1-2q)-p2qw11]/w = [pq2w22 +pqw12(p-q)- p2qw11 ]/w = pq[qw22 +pw12 –qw12-pw11]/w Δq= pq[p(w12-w11) + q(w22-w12)]/w Δp= pq [p(w11-w12) + q(w12-w22)]/w
Επιλογή εναντίον υπολειπόμενων αλληλομόρφων ΕΕ Ee ee Σύνολο p2 2pq q2 1 1 1 1-s p2 2pq q2(1-s) 1-sq2 Δq = pq[p(1-1) + q(1-s-1)] / (1-sq2) = -spq2/ (1-sq2) Αν s = 1 Δq = - q2/(1+q) Δq = q’- q q’ = q + Δq = q/(1+q) q’’= q/(1+2q) …. qt = q/(1+tq) t = (1/qt) – (1/q) Δηλαδή, η επιλογή εναντίον ενός υπολειπόμενου αλληλομόρφου μειώνει τη συχνότητά του στον πληθυσμό. Η μείωση αυτή εξαρτάται από τον συντελεστή επιλογής και την συχνότητα του αλληλομόρφου στον πληθυσμό. Στην περίπτωση που το αλληλόμορφο είναι αβιώσιμο ή οι ομοζυγώτες για το αλληλόμορφο αυτό δεν αναπαράγονται σημαίνει ότι ο γονότυπος αυτός έχει συντελεστή προσαρμογής μηδέν και η επιλογή εναντίον του μέγιστη, δηλαδή ίση με τη μονάδα. Στην περίπτωση αυτή η αλλαγή Δq εξαρτάται μόνο από τη συχνότητα q. Έχει αναφερθεί ότι η συχνότητα του αλληλομόρφου HALn στους χοίρους της φυλής Pietrain ανέρχεται σε 0,92. Τα ομοζυγωτά άτομα αντιδρούν θετικά στη δοκιμασία αλοθάνης (αναισθητικό). Αν θέλουμε να τη μειώσουμε στο 0,01 θα χρειαστούν περίπου 99 γενιές ακόμα κι αν ο συντελεστής επιλογής είναι 1.
Επιλογή εναντίον κυρίαρχων αλληλομόρφων EE Ee ee Σύνολο p2 2pq q2 1 1-s 1-s 1 p2(1-s) 2pq(1-s) q2 1-sp(2-p) Δp = pq[p(1-s-1+s)+q(1-s-1)] / [1-sp(2-p)] =-spq2 / [1-sp(2-p)] =-sp(1-p)2 / [1-sp(2-p)] Αν s=1 1-sp(2-p) 1- p(2-p) = 1-2p+p2 = 1-p –p + p2 = q – p(1-p) =q- pq = q(1-p) = q2 Δp = - pq2 / q2 = -p Δηλαδή, η συχνότητα του Ε βαίνει μειούμενη έως ότου ενδεχομένως εξαφανιστεί από τον πληθυσμό. Σε περίπτωση που το Ε είναι κυρίαρχο αβιώσιμο ή τα άτομα Ε_ αποκλείονται από την αναπαραγωγή τότε απομακρύνεται από τον πληθυσμό σε μα γενιά.
Επιλογή υπέρ των ετεροζυγωτών (Υπερκυριαρχία) ΑΑ Αα αα Σύνολο p2 2pq q2 1 1-s 1 1-t p2(1-s) 2pq q2(1-t) 1-sp2-tq2 Δq = pq[p(1-1+s)+q(1-t-1)] /(1-sp2-tq2) Δq = pq (sp-tq)/ (1-sp2-tq2) Αν Δq=0 sp-tq =0 ↔ s(1-q)=tq ↔ qe=s/(s+t) Οι προηγούμενες περιπτώσεις είδαμε ότι οδηγούν στην καθήλωση του ενός ή του άλλου αλληλομόρφου στον πληθυσμό, με τελικό βέβαια αποτέλεσμα την μείωση της γενετικής παραλλακτικότητας του πληθυσμού. Εδώ, με την επιλογή υπέρ των ετεροζυγωτών συμβαίνει ακριβώς το αντίθετο. Δηλαδή επιτυγχάνεται μια σταθερή ισορροπία όπου διατηρούνται και τα δύο αλληλόμορφα στον πληθυσμό, σε συχνότητες που εξαρτώνται από τους συντελεστές επιλογής εναντίον των δύο ομοζυγωτών γονοτύπων.
Επιλογή εναντίον ετεροζυγωτών (Υποκυριαρχία) ΑΑ Αα αα Σύνολο p2 2pq q2 1 1 1-s 1 p2 2pq(1-s) q2 1-2spq Δp= pq[p(1-1+s)+q(1-s-1)] /(1-2spq) Δp= s(p-q)pq/(1-2spq) Το πρόσημο εξαρτάται από τη διαφορά (p - q) Σε αυτή την περίπτωση, όπου οι συντελεστές προσαρμογής των δύο ομοζυγωτών γονοτύπων είναι ίσοι μεταξύ τους, αυτό που συμβαίνει είναι η μείωση της συχνότητας του αλληλομόρφου με την μικρότερη συχνότητα. Χοίρους: S_ ευαίσθητα στο E.coli, ss ανθεκτικά (διάρροια των νεογέννητων) Αν ss άτομα έχουν πρόσφατα εισαχθεί στον πληθυσμό. Οι S_ χοιρομητέρες αναπτύσσουν αντισώματα στο Ε.coli τα οποία και περνούν στα έμβρυα τους, τα οποία προστατεύουν από τη διάρροια. Η επιλογή εναντίον του S μειώνεται. Παράλληλα οι Ss απόγονοι των ss χοιρομητέρων δεν έχουν λάβει αντισώματα από τις ss μητέρες και είναι ευαίσθητα στο e.coli. Έχουμε δηλαδή επιλογή εναντίον των ετεροζυγωτών, ενώ οι δύο ομοζυγώτες μπορεί να θεωρηθούν ως έχοντες τον ίδιο συντελεστή προσαρμογής. Όταν το βακτήριο εξαφανιστεί, παύει η επιλογή εναντίον των ετεροζυγωτών. Μέχρι την επόμενη έξαρση του βακτηρίου στον πληθυσμό, οπότε και θα ξαναρχίσει ο κυκλος της επιλογής εναντίον των ετεροζυγωτών, αν το αλληλόμορφο s έχει συχνότητα μεγαλύτερη του 0,5 τότε η επιλογή εναντίον των ετεροζυγωτών θα αυξήσει περαιτέρω την συχνότητά του. Αντίθετα, αν είναι μικρότερη του 0,5 θα την μειώσει περισσότερο.
Φαινότυπος = Γονότυπος + Περιβάλλον
Ποιοτικοί και Ποσοτικοί Χαρακτήρες ΑΣΥΝΕΧΗΣ aa Aa AA ΣΥΝΕΧΗΣ
Μεριστικοί χαρακτήρες
Χαρακτήρες ουδού
Υποθετικό παράδειγμα γενετικών και περιβαλλοντικών επιδράσεων στο ύψος φυτών P = G P = E P = G + E P = G+ E+ GxE
Ερωτήματα της ποσοτικής γενετικής Σε ποιο βαθμό η παρατηρούμενη φαινοτυπική ποικιλομορφία απορρέει από διαφορές στο γονότυπο και σε ποιο βαθμό αντανακλά την επίδραση που έχει το εκάστοτε περιβάλλον; Πόσα γονίδια καθορίζουν τον φαινότυπο; Έχουν όλα τα γονίδια ίση συνεισφορά στο φαινότυπο; Ποιες οι σχέσεις μεταξύ των αλληλομόρφων τους; Όταν υπάρχει επιλογή ενός φαινοτύπου πόσο γρήγορα μεταβάλλεται η σύσταση του πληθυσμού για τον χαρακτήρα αυτό; Μεταβάλλονται άλλοι χαρακτήρες ταυτόχρονα; Ποια είναι η καλύτερη μέθοδος για την επιλογή και διασταύρωση ατόμων έτσι ώστε να προκύψουν οι επιθυμητοί φαινότυποι στους απογόνους;
Κληρονομικότητα των ποσοτικών χαρακτήρων Francis Galton, Karl Pearson τέλη του 19ου αι. Στατιστική συσχέτιση μεταξύ γονέων και απογόνων Έντονη διαφωνία για το αν κληρονομούνται με βάση τους νόμους του Mendel William Johansen 1903 Έδειξε ότι το βάρος των σπόρων της φασολιάς επηρεάζεται από γενετικούς και περιβαλλοντικούς παράγοντες Πολυπαραγοντικοί χαρακτήρες
Η πολυγονιδιακή υπόθεση 1909 Hermann Nilsson-Ehle χρώμα των σπόρων του σιταριού P: [κόκκινο] x [λευκό] F1: 100% [ενδιάμεσο] F2: 15[κόκκινα]: 1[λευκό] 1[βαθύ σκούρο κ.]: 4[σκούρο κ.]: 6[ενδιαμ]: 4[ανοιχτό κ.]: 1[λευκό] P: RRCC x rrcc F1: RrCc F2:1RRCC: 2RrCC: 1rrCC: 2RRCc: 4RrCc: 2rrCc: 1RRcc : 2Rrcc: 1rrcc
(p+q)4 = p4 + 4p3q + 6p2q2 + 4pq3+ q4 p = q = ½
Ποσοτική γενετική ανάλυση 1913 Emerson and East
Ποσοτικοί χαρακτήρες Συνεχείς χαρακτήρες Πολυγονιαδική υπόθεση Επίδραση του περιβάλλοντος Κανονική κατανομή
Έλεγχος κληρονομικής βάσης ενός χαρακτήρα
Η κανονική κατανομή Μέσος όρος μ Διακύμανση σ2 Τυπική απόκλιση σ μ: τα μισά άτομα θα έχουν τιμές μεγαλύτερες του μέσου και τα υπόλοιπα μισά τιμές μικρότερες του μέσου σ2 : είναι ένα άθροισμα τετραγώνων και έχει πάντα θετική τιμή. Η ποσότητα που υψώνεται στο τετράγωνο είναι η απόκλιση της κάθε μέτρησης από τον μέσο του πληθυσμού. Άρα όσο περισσότερο αποκλίνει μια τιμή από τον μέσο τόσο περισσότερο συμβάλει στο μέγεθος της διασποράς των τιμών από τον μέσο. Και όσο πιο πολλές παρατηρήσεις εμφανίζουν μεγαλύτερες αποκλίσεις τόσο μεγαλύτερη η διακύμανση. Έτσι η διακύμανση είναι πιο ευαίσθητος δείκτης ποικιλομορφίας από ότι η ελάχιστη και η μέγιστη τιμή. Το γεγονός ότι η διασπορά είναι άθροισμα σημαίνει ότι αν χωρίσω τις παρατηρήσεις μου σε ομάδες, η διασπορά του συνόλου των παρατηρήσεων είναι το άθροισμα της διασποράς μέσα στις ομάδες και της διασποράς μεταξύ των ομάδων. Δείγμα
Ίδιος μέσος όρος αλλά διαφορετικές τιμές διακύμανσης.
Το ποσοστό των δεδομένων που περιλαμβάνονται σε διαστήματα με κέντρο το μέσο όρο και πλάτος πολλαπλάσιο της τυπικής απόκλισης.
Φαινοτυπική και Γενετική διακύμανση
Φαινοτυπική ποικιλομορφία (διακύμανση) VP = VG + VE + VGxE VG = VA + VD + VI VE VP = VA + VD + VI + VE + VGxE
Συντελεστής κληρονομικότητας Το ποσοστό της φαινοτυπικής ποικιλομορφίας ενός πληθυσμού που αποδίδεται σε γενετικούς παράγοντες Η2 = VG/VP (ευρεία έννοια) h2 = VA/VP (στενή έννοια) Δηλαδή σε ποιο βαθμό τα γονίδια προσδιορίζουν την παρατηρούμενη ποικιλομορφία του χαρακτήρα στον συγκεκριμένο πληθυσμό Όχι σε ποιο βαθμό τα γονίδια προσδιορίζουν τον χαρακτήρα Αφορά πληθυσμούς και όχι άτομα Δεν υπάρχει καθολικό H2, h2 για ένα χαρακτήρα Υψηλές τιμές του H2 , h2 δεν αποκλείουν το ενδεχόμενο επιρροής του χαρακτήρα από το περιβάλλον
Επιλογή των ποσοτικών χαρακτήρων Σταθεροποιούσα επιλογή (Stabilizing selection) Κατευθύνουσα επιλογή (Directional selection) Διαφοροποιούσα επιλογή (Disruptive selection) Σχέση φαινοτυπικής τιμής και συντελεστών προσαρμογής. Α) Σταθεροποιούσα επιλογή παρατηρείται όταν ο ενδιάμεσος φαινότυπος έχει τον υψηλότερο συντελεστή προσαρμογής. Επιλογή υπέρ των ετεροζυγωτών που είδαμε ότι στην ισορροπία διατηρεί και τα δύο αλληλόμορφα στον πληθυσμό (qe= s/(s+t), wAA = 1-s, wAa=1, waa=1-t). B) Κατευθύνουσα επιλογή παρατηρείται όταν ο ακραίος φαινότυπος έχει τον υψηλότερο συντελεστή προσαρμογής. Στην περίπτωση αυτή η μέση προσαρμογή του πληθυσμού αυξάνεται μονοτονικά και τείνει προς την μονάδα. Επιλογή υπέρ ΑΑ ή υπέρ αα. Γ) Διαφοροποιούσα επιλογή παρατηρείται όταν και οι δύο ακραίοι φαινότυποι έχουν υψηλότερο συντελεστή προσαρμογής από ότι οι ενδιάμεσοι φαινότυποι. Επιλογή εναντίον των ετεροζυγωτών. (βλέπε διάλεξη 5).
Κατευθύνουσα επιλογή και ποσοτικοί χαρακτήρες Η πιο συχνή μέθοδος τεχνητής επιλογής Επιλέγονται ως γεννήτορες άτομα που ο φαινότυπός τους υπερβαίνει ένα όριο στην τιμή του ποσοτικού χαρακτήρα που μας ενδιαφέρει Η τεχνητή επιλογή εστιάζεται συνήθως σε ένα χαρακτήρα και όχι στο σύνολο της προσαρμογής Η τεχνητή επιλογή εφαρμόζεται σε μικρούς (εργαστηριακούς) πληθυσμούς που είναι μικρότεροι από τους φυσικούς πληθυσμούς και κλειστοί, δηλαδή γενετικά απομονωμένοι από άλλους πληθυσμούς
Κατευθύνουσα επιλογή ποσοτικών χαρακτήρων και h2 R = XF1 - XP Η ικανότητα να προβλέψουμε το R, αν είναι γνωστό το h2, έγκειται στο γεγονός ότι μας επιτρέπει να σχεδιάσουμε προγράμματα βελτίωσης και ιδιαίτερα να συγκρίνουμε διαφορετικά βελτιωτικά σχήματα μεταξύ τους. Άρα όλοι οι παράγοντες που μπορεί να επηρεάζουν την ακρίβεια της πρόβλεψης αυτής είναι σημαντικοί για τον βελτιωτή. Μια πρώτη ματιά εντοπίζει 3 παράγοντες που ενδέχεται να επηρεάσουν το R. Εδώ, ο συντελεστής κληρονομικότητας είναι αυτός με τη στενή έννοια, δηλαδή αφορά μόνο το ποσοστό της φαινοτυπικής διακύμανσης που οφείλεται σε αλληλόμορφα με προσθετική δράση. Αυτό είναι λογικό από τη στιγμή που ο κάθε γονέας μεταβιβάζει αλληλόμορφα, μέσω των γαμετών, στους απογόνους του και όχι το σύνολο του γονοτύπου του. Με άλλα λόγια, καθώς δεν μεταβιβάζει τις σχέσεις των αλληλομόρφων του (κυριαρχία ή επίσταση), το μέρος του γενετικού του δυναμικού που μπορεί να μεταβιβάσει είναι αυτό που εκφράζεται από την προσθετική δράση των αλληλομόρφων του (την κληροδοτική αξία του γονοτύπου του Α, βλέπε διάλεξη 7- διαφάνεια 13). S = Xe-XP S = Διαφορικό Επιλογής R = Απόκριση στην επιλογή
R= h2 x S μο = ΧΡ μ = Χe μ1 = XF1
