Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
ΚΥΤΤΑΡΙΚΗ ΟΡΓΑΝΩΣΗ ΚΝΣ 100 δισ. νευρώνες!

2
Figure 21-99. A typical neuron of a vertebrate. The arrows indicate the direction in which signals are conveyed. The neuron shown is from the retina of a monkey. The longest and largest neurons in a human extend for about 1 million μm and have an axon diameter of 15 μm. (Drawing of neuron from B.B. Boycott in Essays on the Nervous System [R. Bellairs and E.G. Gray, eds.]. Oxford, UK: Clarendon Press, 1974.)
.")
3
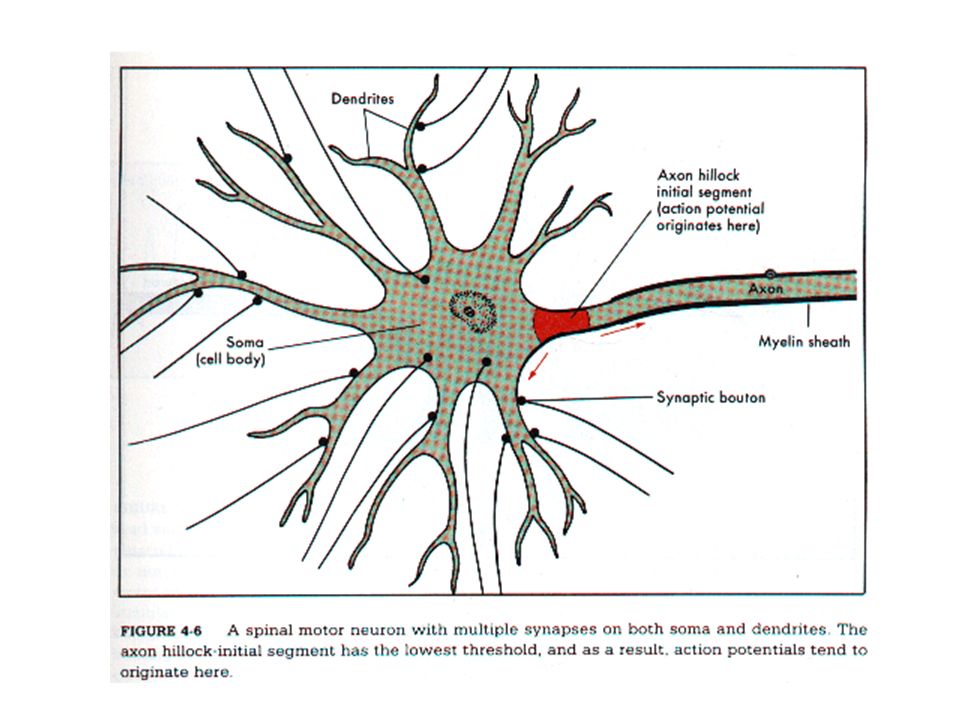
Κυτταρικό σώμα Δενδρίτες-προεκβολή, συσκευή υποδοχής σημάτων από άλλα νευρικά κύτταρα Νευράξονας-προεκβολή, μεταφέρει σήματα (δυναμικά ενέργειας) Προσυναπτικά κομβία
 Προσυναπτικά κομβία")
4
Figure 21-100. The complex organization of nerve cell connections. This semischematic drawing depicts a section through a small part of a mammalian brain - the olfactory bulb of a dog, stained by the Golgi technique. The black objects are neurons; the thin lines are axons and dendrites, through which the various sets of neurons are interconnected according to precise rules. (From C. Golgi, Riv. sper. freniat. Reggio-Emilia 1:405-425, 1875; reproduced in M. Jacobson, Developmental Neurobiology, 3rd ed. New York: Plenum, 1992.)
.")
5
Δυναμικά ενέργειας - όλον ή ουδέν, σύντομες παροδικές ώσεις, 100mV 1ms Η ώση αναγεννάται καθώς οδεύει κατά μήκος του νευράξονα Για να εξασφαλισθεί η ταχεία αγωγή μυελώδες έλυτρο-κόμβοι Ranvier ΣΥΝΑΨΗ, προσυναπτικός και μετασυναπτικός νευρώνας Οι περισσότεροι νευράξονες καταλήγουν κοντά στους δενδρίτες ενός μετασυναπτικού κυττάρου αλλά και στο κυτταρικό σώμα ή πιο σπάνια στο αρχικό ή τελικό τμήμα του νευράξονα.

6
Μονόπολα: μια αποφυάδα, πολλοί κλάδοι, ο ένας λειτουργεί σαν νευράξονας, οι άλλοι ως δενδρίτες υποδοχής (ασπόνδυλα, αυτόνομα γάγγλια) Δίπολα: δενδρίτης (μεταφέρει πληρ. από περιφ. στο σώμα, νευράξονας, μεταφέρει πληρ. από σώμα στο ΚΝΣ) [αισθητικοί] Πολύπολα:υπερτερούν στα σπονδυλωτά, μεγάλη ποικιλία, διαφορές σε μήκος αποφυάδων και αριθμό δενδριτών. Κινητικό κύτταρο του ΝΜ δέχεται ως και 10.000 επαφές (2000 στο σώμα, 8000 στους δενδρίτες). Κύτταρα Purkinje 150.000 επαφές!
 [αισθητικοί] Πολύπολα:υπερτερούν στα σπονδυλωτά, μεγάλη ποικιλία, διαφορές σε μήκος αποφυάδων και αριθμό δενδριτών. Κινητικό κύτταρο του ΝΜ δέχεται ως και επαφές (2000 στο σώμα, 8000 στους δενδρίτες). Κύτταρα Purkinje επαφές!.")
7
Οι νευρώνες ταξινομούνται με βάση τη λειτουργία τους σε 3 ομάδες Αισθητικοί, κινητικοί, διάμεσοι Αισθητικοί: μεταφέρουν πληροφορίες για αντίληψη και κινητικό συντονισμό Κινητικοί: μεταφέρουν εντολές σε μυς και αδένες Διάμεσοι : οι πλέον πολυάριθμοι μεταφέρουν πληρ. σε μεγάλες αποστάσεις ή σε τοπικά δίκτυα

10
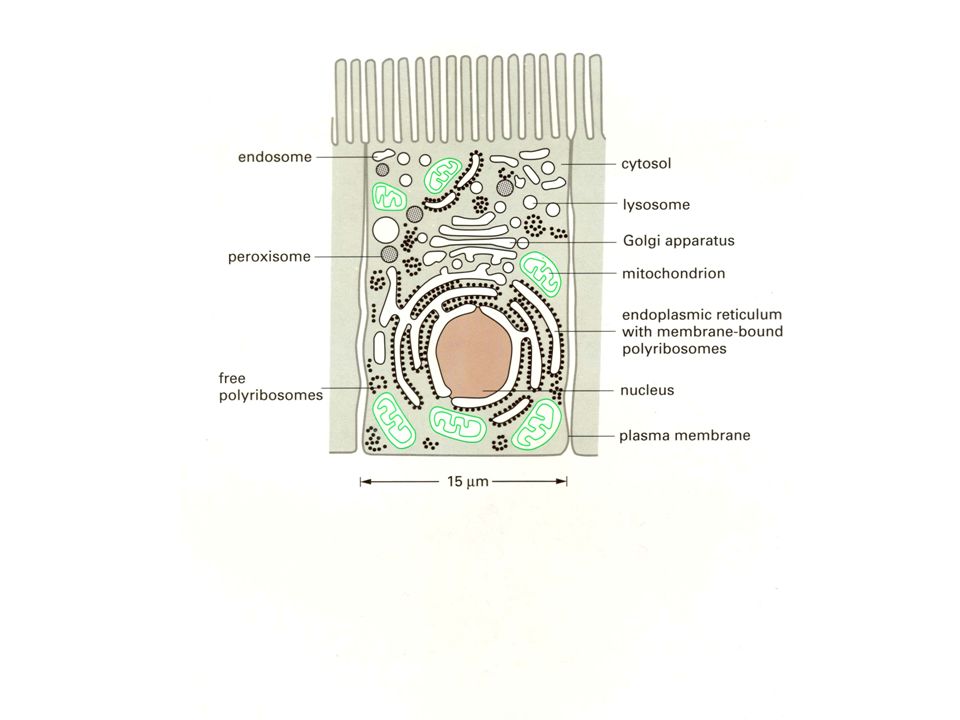
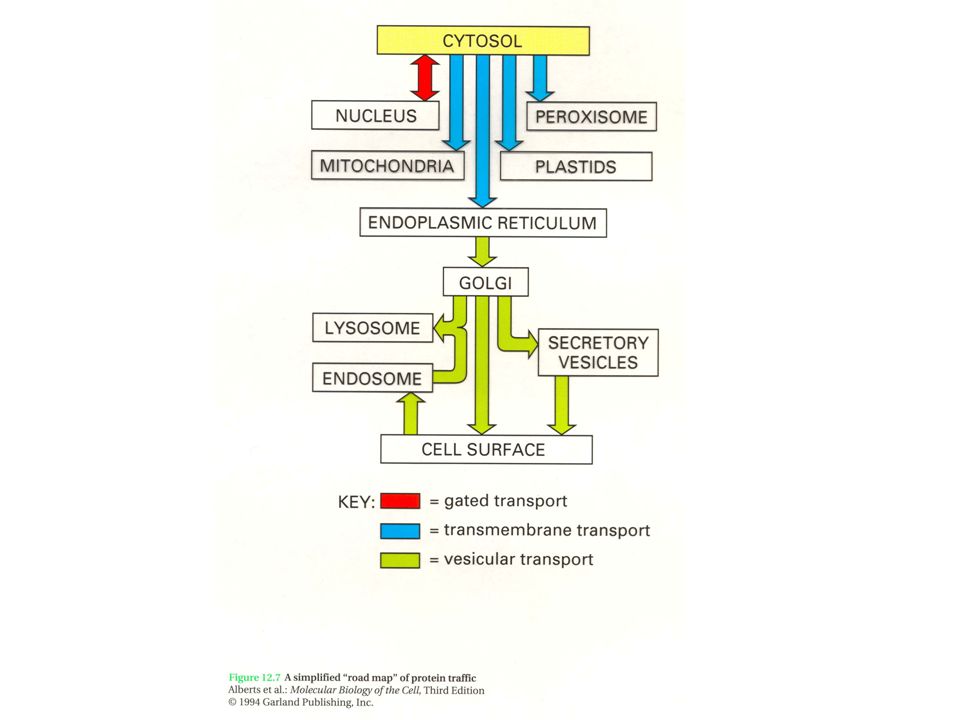
ΟΡΓΑΝΙΔΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΠΥΡΗΝΑΣ Πυρήνας σχετικά άμορφος εκτός του πυρηνίσκου 10-20χ περισσότερες αλληλουχίες mRNA από ότι στο νεφρό ή στο ήπαρ

11
ΟΡΓΑΝΙΔΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΚΥΤΤΑΡΙΚΗ ΜΕΜΒΡΑΝΗ Ειδικοί δίαυλοι ιόντων, υποδοχείς για νευροδιαβιβαστές, κλπ

12
Golgi apparatus in the thalamocortical relay neuron. N - nucleus, Golgi sacculi - red arrow, Golgi vesicles - blue arrow. Scale = 300 nm. (Rat, lateral geniculate nucleus.) ΟΡΓΑΝΙΔΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΣΤΗΜΑ GOLGI
 ΟΡΓΑΝΙΔΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΣΥΣΤΗΜΑ GOLGI.")
13
Axon initial segment of the pyramidal cell containing bunches of microtubules (arrow) which are linked together in transversal section (inset). Scale = 1 µm. (Mouse, neocortex.) ΟΡΓΑΝΙΔΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ
 ΟΡΓΑΝΙΔΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟΣ.")
14
Axon initial segment (arrow) originating from the axon hillock at the base of a pyramidal cell (N -nucleus). Scale = 1 µm. (Mouse, neocortex.) ΟΡΓΑΝΙΔΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΑΥΧΕΝΑΣ ΝΕΥΡΑΞΟΝΑ
 ΟΡΓΑΝΙΔΙΑ ΝΕΥΡΙΚΩΝ ΚΥΤΤΑΡΩΝ ΑΥΧΕΝΑΣ ΝΕΥΡΑΞΟΝΑ.")
16
Οι νευρώνες διατηρούν μια διαφορά φορτίου της τάξης των -65μV κατά μήκος της κυτταρικής μεμβράνης (δυναμικό ηρεμίας, το εσωτερικό αρνητικά φορτισμένο σε σχέση με το εξωτερικό). Αντλία Κ, Να αυτή κρατάει την [Να] χαμηλή εντός και την [Κ] υψηλή εντός. Το δυναμικό ηρεμίας οφείλεται σε 2 ιδιότητες α) διαφορές [] Κ και Να β) υψηλή διαπερατότητα της μεμβράνης από το Κ και χαμηλή από το Να. ‘Ετσι, το Κ φεύγει προς τα έξω με αποτέλεσμα να γίνεται το εσωτερικό πιο ηλεκτροαρνητικό. Αυτό πυροδοτεί δυναμικό ενέργειας=σήμα αγωγής του νευρώνα. Κατά την διάρκεια του ΔΕ η μεμβράνη γίνεται διαπερατή από Να και μετά από λίγο επιστρέφει στην κατάσταση ηρεμίας. Το ΔΕ αναγεννάται στους κόμβους του Ranvier.
 διαφορές [] Κ και Να β) υψηλή διαπερατότητα της μεμβράνης από το Κ και χαμηλή από το Να. ‘Ετσι, το Κ φεύγει προς τα έξω με αποτέλεσμα να γίνεται το εσωτερικό πιο ηλεκτροαρνητικό. Αυτό πυροδοτεί δυναμικό ενέργειας=σήμα αγωγής του νευρώνα. Κατά την διάρκεια του ΔΕ η μεμβράνη γίνεται διαπερατή από Να και μετά από λίγο επιστρέφει στην κατάσταση ηρεμίας. Το ΔΕ αναγεννάται στους κόμβους του Ranvier..")
17
Ταχεία νευραξονική μεταφορά ορθόδρομη (συναπτικά κυστίδια)—εξαρτάται από ATP και μικροσωληνίσκους Δημιουργούν μονοπάτια πάνω στα οποία μέσω κινησίνης (ATPάση) τα οργανίδια κινούνται με άλματα ανάδρομη –επιστρέφουν υλικά από τις απολήξεις στο κυτταρικό σώμα, ταχύτητα κατά το ήμισυ, επί μικροσωληνίσκων, κινητήριο μόριο δυνεΐνη Εκτός από την συλλογή «απορριμμάτων» η ανάδρομη μεταφορά παίζει ρόλο στην πληροφόρηση του σώματος για τα συμβάντα στα άκρα (π.χ. NGF) Βραδεία αξονοπλασματική ροή Κυτοσόλιο, 2 κινητικά στοιχεία(νευρικά νημάτια)
 Βραδεία αξονοπλασματική ροή Κυτοσόλιο, 2 κινητικά στοιχεία(νευρικά νημάτια).")
18
The individual polypeptides of intermediate filaments are elongated molecules with an extended central α-helical domain that forms a parallel coiled coil with another monomer. A pair of parallel dimers then associates in an antiparallel fashion to form a staggered tetramer. This tetramer represents the soluble subunit that is analogous to the αβ-tubulin dimer, or the actin monomerpolypeptidesintermediate filamentsmoleculesdomain monomerantiparallelsubunittubulinactinmonomer

19
Figure 16-20 Two types of intermediate filaments in cells of the nervous system (A) Freeze-etch electron microscopic image of neurofilaments in a nerve cell axon, showing the extensive cross-linking through protein cross-bridges—an arrangement believed to give this long cell process great tensile strength. The cross-bridges are formed by the long, nonhelical extensions at the C-terminus of the largest neurofilament protein (NF-H). (B) Freeze-etch image of glial filaments in glial cells, showing that these intermediate filaments are smooth and have few cross-bridges. (C) Conventional electron micrograph of a cross section of an axon showing the regular side-to-side spacing of the neurofilaments, which greatly outnumber the microtubules. (A and B, courtesy of Nobutaka Hirokawa; C, courtesy of John Hopkins.)electronneurofilamentsnerve cellaxonproteinneurofilamentproteinglial cellsintermediate filamentselectronmicrographsectionaxonneurofilamentsmicrotubules Intermediate filamentsIntermediate filaments are particularly prominent in the cytoplasm of cells that are subject tocytoplasm mechanical stress, and their major function seems to be to impart physical strength to cells and tissues
. (B) Freeze-etch image of glial filaments in glial cells, showing that these intermediate filaments are smooth and have few cross-bridges. (C) Conventional electron micrograph of a cross section of an axon showing the regular side-to-side spacing of the neurofilaments, which greatly outnumber the microtubules. (A and B, courtesy of Nobutaka Hirokawa; C, courtesy of John Hopkins.)electronneurofilamentsnerve cellaxonproteinneurofilamentproteinglial cellsintermediate filamentselectronmicrographsectionaxonneurofilamentsmicrotubules Intermediate filamentsIntermediate filaments are particularly prominent in the cytoplasm of cells that are subject tocytoplasm mechanical stress, and their major function seems to be to impart physical strength to cells and tissues.")
20
Three types of neurofilament proteins (NF-L, NF-M, NF-H) coassemble in vivo, forming heteropolymers that contain NF-L plus one of the others. The NF-H and NF-M proteins have lengthy C-terminal tail domains that bind to neighboring filaments, generating aligned arrays with a uniform interfilament spacing. During axonal growth, new neurofilament subunits are incorporated all along the axon in a dynamic process that involves the addition of subunits along the filament length, as well as the addition of subunits at the filament ends. After an axon has grown and connected with its target cell, the diameter of the axon may increase as much as fivefold. The level of neurofilament gene expression seems to directly control axonal diameter, which in turn controls how fast electrical signals travel down the axon.neurofilamentproteins in vivoproteinsdomainsneurofilamentsubunitsaxonsubunits axon neurofilamentgeneexpressionaxon

22
Figure 21-102. Diagram of an early (2 1/2-day) chick embryo, showing the origins of the nervous system. The neural tube (light green) has already closed, except at the tail end, and lies internally, beneath the ectoderm, of which it was originally a part (see Figure 21-10). The neural crest (red) lies dorsally beneath the ectoderm, in or above the roof of the neural tube. In addition, thickenings, or placodes (dark green), in the ectoderm of the head give rise to some of the sensory transducer cells and neurons of that region, including those of the ear and the nose. The cells of the retina of the eye, by contrast, originate as part of the neural tube.
 chick embryo, showing the origins of the nervous system. The neural tube (light green) has already closed, except at the tail end, and lies internally, beneath the ectoderm, of which it was originally a part (see Figure 21-10). The neural crest (red) lies dorsally beneath the ectoderm, in or above the roof of the neural tube. In addition, thickenings, or placodes (dark green), in the ectoderm of the head give rise to some of the sensory transducer cells and neurons of that region, including those of the ear and the nose. The cells of the retina of the eye, by contrast, originate as part of the neural tube..")
23
2-day chick embryo.

24
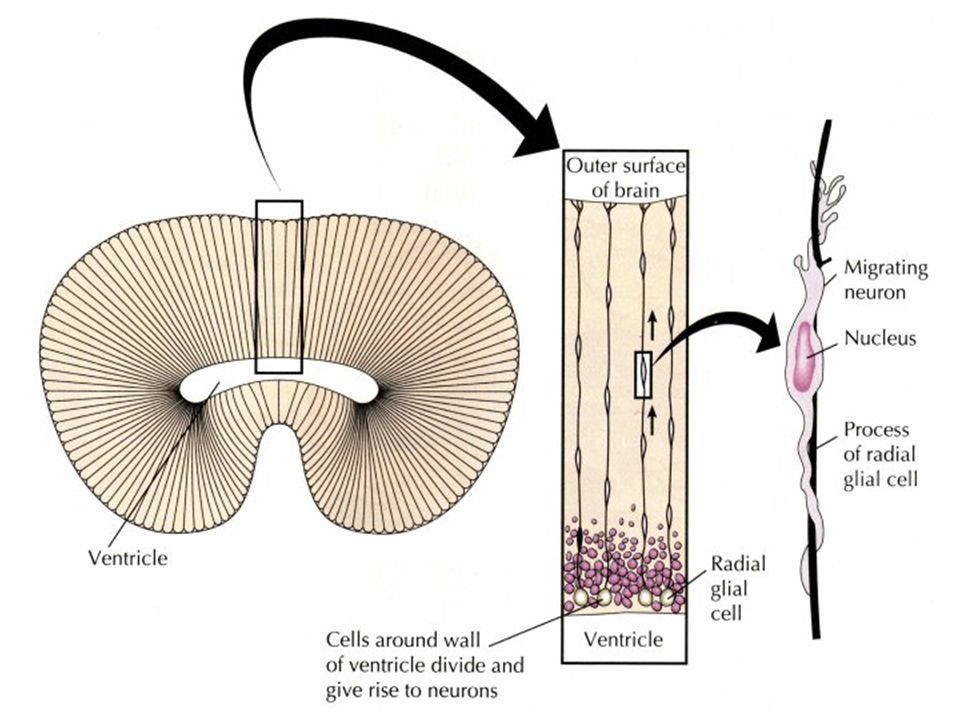
Figure 21-104. Migration of immature neurons along radial glial cells. The diagrams are based on reconstructions from sections of the cerebral cortex of a monkey (part of the neural tube). The neurons are born close to the inner, luminal surface of the neural tube and migrate outward. The radial glial cells can be considered as persisting cells of the original columnar epithelium of the neural tube that become extraordinarily stretched as the wall of the tube thickens. (After P. Raki#180c, J. Comp. Neurol. 145:61-84, 1972
. The neurons are born close to the inner, luminal surface of the neural tube and migrate outward. The radial glial cells can be considered as persisting cells of the original columnar epithelium of the neural tube that become extraordinarily stretched as the wall of the tube thickens. (After P. Raki#180c, J. Comp. Neurol. 145:61-84,")
25
Figure 21-105. Comparison of the layering of neurons in the cortex of normal and reeler mice. In the reeler mutant an abnormality of cell migration causes an approximate inversion of the normal relationship between neuronal birthday and position. The misplaced neurons nevertheless differentiate according to their birthdays and make the connections appropriate to their birthdays

26
Figure 21-106. Growth cones in the developing spinal cord of a 3-day chick embryo. The drawing shows a cross-section stained by the Golgi technique. Most of the neurons, apparently, have as yet only one elongated process - the future axon. The growth cones of the interneurons remain inside the spinal cord, those of the motor neurons emerge from it (to make their way toward muscles), and those of the sensory neurons grow into it from outside (where their cell bodies lie). Many of the cells in the more central regions of the embryonic spinal cord are still proliferating and have not yet begun to differentiate as neurons or glial cells. (From S. Ramón y Cajal, Histologie du Système Nerveux de l'Homme et des Vertébrés. Paris: Maloine, 1909-1911; reprinted, Madrid: C.S.I.C., 1972.)
, and those of the sensory neurons grow into it from outside (where their cell bodies lie). Many of the cells in the more central regions of the embryonic spinal cord are still proliferating and have not yet begun to differentiate as neurons or glial cells. (From S. Ramón y Cajal, Histologie du Système Nerveux de l Homme et des Vertébrés. Paris: Maloine, ; reprinted, Madrid: C.S.I.C., 1972.).")
27
Figure 21-107. Formation of axon and dendrites in culture. A young neuron has been isolated from the brain of a mammal and put to develop in culture, where it sends out processes. One of these processes, the future axon, has begun to grow out faster than the rest (the future dendrites) and has bifurcated. (A) A phase-contrast picture; (B) the pattern of staining with fluorescent phalloidin, which binds to filamentous actin. Actin is concentrated in the growth cones at the tips of the processes that are actively extending and at some other sites of lamellipodial activity. (Courtesy of Kimberly Goslin, from Z.W. Hall, An Introduction to Molecular Neurobiology. Sunderland, MA: Sinauer, 1992.)
 and has bifurcated. (A) A phase-contrast picture; (B) the pattern of staining with fluorescent phalloidin, which binds to filamentous actin. Actin is concentrated in the growth cones at the tips of the processes that are actively extending and at some other sites of lamellipodial activity. (Courtesy of Kimberly Goslin, from Z.W. Hall, An Introduction to Molecular Neurobiology. Sunderland, MA: Sinauer, 1992.).")
28
Figure 21-108. Neural growth cones. (A) Scanning electron micrograph of growth cones at the end of a neurite put out by a chick sympathetic neuron in culture. Here a previously single growth cone has recently divided in two. Note the many filopodia and the taut appearance of the neurite, due to tension generated by the forward movement of the growth cones, which are often the only firm points of attachment to the substratum. (B) Scanning electron micrograph of the growth cone of a sensory neuron in vivo crawling over the inner surface of the epidermis of a Xenopus tadpole. (A, from D. Bray, in Cell Behaviour [R. Bellairs, A. Curtis, and G. Dunn, eds.].
 Scanning electron micrograph of growth cones at the end of a neurite put out by a chick sympathetic neuron in culture. Here a previously single growth cone has recently divided in two. Note the many filopodia and the taut appearance of the neurite, due to tension generated by the forward movement of the growth cones, which are often the only firm points of attachment to the substratum. (B) Scanning electron micrograph of the growth cone of a sensory neuron in vivo crawling over the inner surface of the epidermis of a Xenopus tadpole. (A, from D. Bray, in Cell Behaviour [R. Bellairs, A. Curtis, and G. Dunn, eds.]..")
29
Figure 21-109. NGF effects on neurite outgrowth. Dark-field photomicrographs of a sympathetic ganglion cultured for 48 hours with (above) or without (below) NGF. Neurites grow out from the sympathetic neurons only if NGF is present in the medium. Each culture also contains Schwann (glial) cells that have migrated out of the ganglion; these are not affected by NGF. Neuronal survival and maintenance of growth cones for neurite extension represent two distinct effects of NGF. The effect on growth cones is local, direct, rapid, and independent of communication with the cell body; when NGF is removed, the deprived growth cones halt their movements within a minute or two. The effect of NGF on cell survival is less immediate and is associated with uptake of NGF by endocytosis and its intracellular transport back to the cell body. (Courtesy of Naomi Kleitman.)
 or without (below) NGF. Neurites grow out from the sympathetic neurons only if NGF is present in the medium. Each culture also contains Schwann (glial) cells that have migrated out of the ganglion; these are not affected by NGF. Neuronal survival and maintenance of growth cones for neurite extension represent two distinct effects of NGF. The effect on growth cones is local, direct, rapid, and independent of communication with the cell body; when NGF is removed, the deprived growth cones halt their movements within a minute or two. The effect of NGF on cell survival is less immediate and is associated with uptake of NGF by endocytosis and its intracellular transport back to the cell body. (Courtesy of Naomi Kleitman.).")
30
Figure 21-111. The regeneration of connections between eye and brain in an amphibian after one eye has been rotated. The axons from each part of the rotated retina regenerate so as to reconnect with the part of the tectum appropriate to the original positions of the retinal bodies. Thus, for example, light falling on the ventral part of the rotated retina is perceived as though it were falling on the dorsal part, and the animal sees the world upside down; if food is dangled above it, it makes a lunge downward, and so on.

31
Figure 21-112. Selectivity of retinal axons growing over tectal membranes. The culture substratum has been coated with alternating stripes of membrane prepared either from posterior tectum (P) or from anterior tectum (A); the anterior tectal stripes are made visible by staining them with a fluorescent marker in the vertical strips at the sides of the picture. Axons of neurons from the temporal half of the retina (growing in from the left) follow the stripes of anterior tectal membrane but avoid the posterior tectal membrane, while axons of neurons from the nasal half of the retina (growing in from the right) do the converse. Thus anterior tectum differs from posterior tectum and nasal retina from temporal retina, and the differences guide selective axon outgrowth. These experiments have been done with cells from the chick embryo. (From Y. von Boxberg, S. Diess, and U. Schwarz, Neuron 10:345-357, 1993.)
 or from anterior tectum (A); the anterior tectal stripes are made visible by staining them with a fluorescent marker in the vertical strips at the sides of the picture. Axons of neurons from the temporal half of the retina (growing in from the left) follow the stripes of anterior tectal membrane but avoid the posterior tectal membrane, while axons of neurons from the nasal half of the retina (growing in from the right) do the converse. Thus anterior tectum differs from posterior tectum and nasal retina from temporal retina, and the differences guide selective axon outgrowth. These experiments have been done with cells from the chick embryo. (From Y. von Boxberg, S. Diess, and U. Schwarz, Neuron 10: , 1993.).")
32
Figure 21-113. Sharpening of the retinotectal map by synapse elimination. At first the map is fuzzy because each retinal axon branches widely to innervate a broad region of tectum overlapping the regions innervated by other retinal axons. The map is then refined by synapse elimination. Where axons from separate parts of the retina synapse on the same tectal cell, competition occurs, eliminating the connections made by one of the axons. But axons from cells that are close neighbors in the retina cooperate, maintaining their synapses on shared tectal cells. Thus each retinal axon ends up innervating a small tectal territory, adjacent to and partly overlapping the territory innervated by axons from neighboring sites in the retina

33
Figure 21-114. Synapse elimination and its dependence on the pattern of excitation. In the experiment illustrated schematically here, a neuron and a muscle cell from an embryo have been allowed to form a synapse in vitro. The muscle cell is then stimulated with puffs of acetylcholine (mimicking neural stimulation) either alone or in synchrony with electrical excitation of the neuron. The results illustrate a general principle: each excitation of a target cell tends to cause the rejection of any synapse where the presynaptic axon terminal has just been quiet but to maintain synapses where the presynaptic axon terminal has just been active.
 either alone or in synchrony with electrical excitation of the neuron. The results illustrate a general principle: each excitation of a target cell tends to cause the rejection of any synapse where the presynaptic axon terminal has just been quiet but to maintain synapses where the presynaptic axon terminal has just been active..")
34
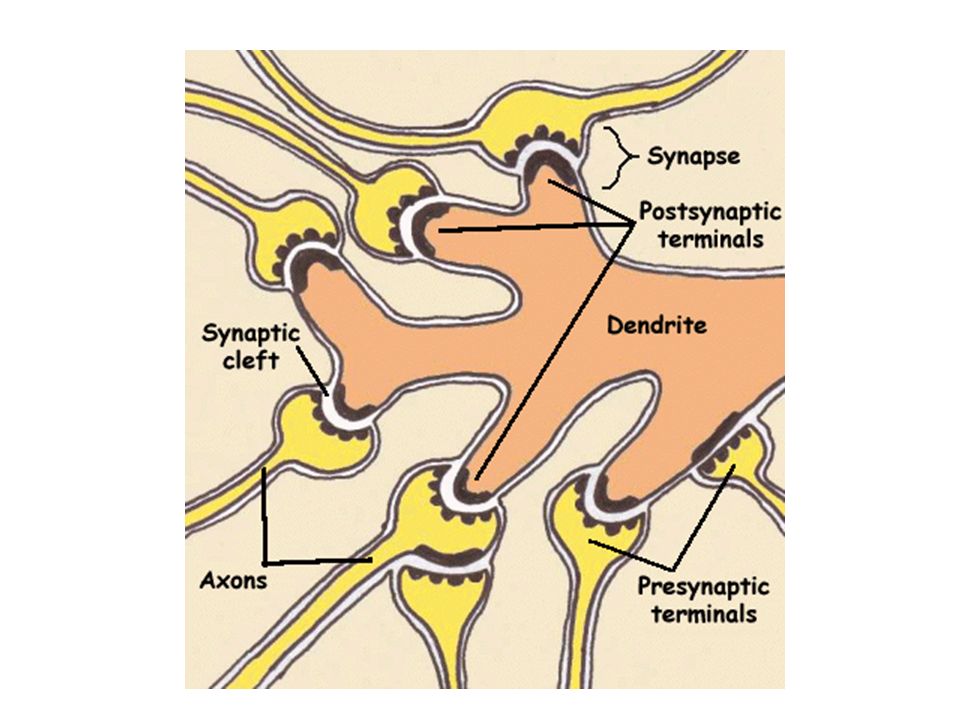
ΣΥΝΑΨΗ

35
ΔΟΜΗ ΣΥΝΑΨΗΣ

36
ΣΥΝΑΠΤΙΚΑ ΚΥΣΤΙΔΙΑ

37
Mitochondria (transversally sectioned) in the presynaptic axonal bouton of an axo-somatic synapse. P - Purkinje cell soma. Scale = 250 nm. (Mouse, cerebellar cortex.) ΜΙΤΟΧΟΝΔΡΙΑ ΣΕ ΣΥΝΑΨΗ ΠΑΡΟΧΗ ΕΝΕΡΓΕΙΑΣ
 ΜΙΤΟΧΟΝΔΡΙΑ ΣΕ ΣΥΝΑΨΗ ΠΑΡΟΧΗ ΕΝΕΡΓΕΙΑΣ.")
39
GLIAL CELLS Oligodendroglial cell Astroglia Microglia

40
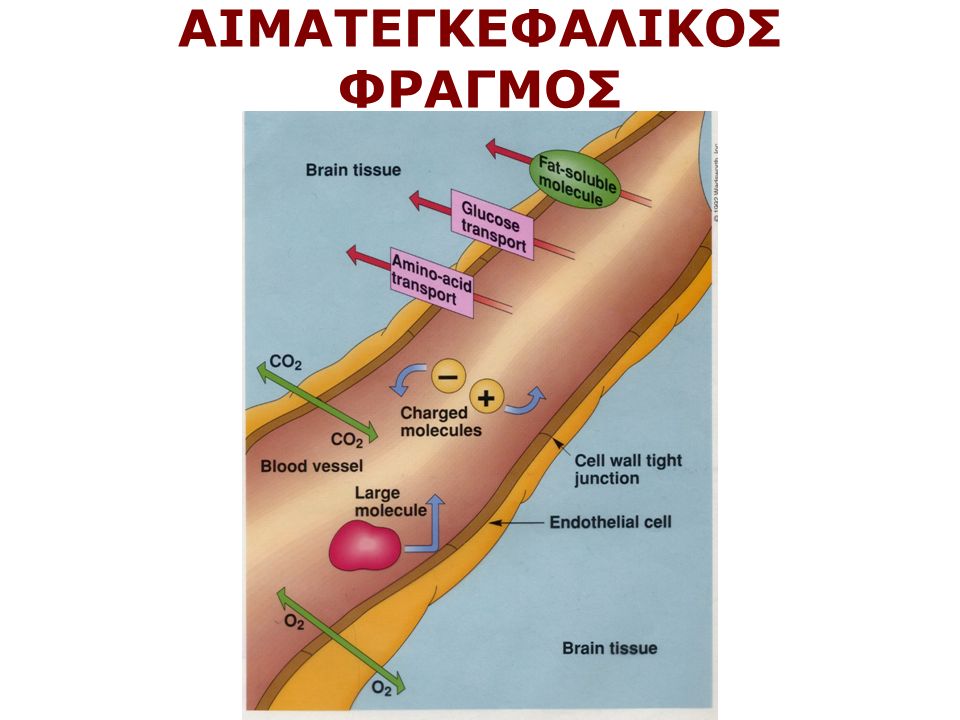
Παρέχουν στήριξη στους νευρώνες Παρέχουν θρεπτικά υλικά στους νευρώνες Λειτουργούν ως απορριμματοσυλλέκτες Συμμετέχουν στη ρύθμιση της συναπτικής λειτουργίας π.χ. Αστροκύτταρα Α. προσλαμβάνοντας ορισμένους νευροδιαβιβαστές π.χ. γλουταμικό, αφού αυτοί δράσουν στη σύναψη Β. ρυθμίζοντας την ιοντική ομοιόσταση Συμμετέχουν στην απάντηση του νευρικού ιστού στο τραύμα (μικρογλοία, αστρογλοίωση) Συμμετέχουν στη δημιουργία του αιματεγκεφαλικού φραγμού π.χ. Αστροκύτταρα Καθοδηγούν τους νευράξονες κατά την ανάπτυξη (Ακτινωτή γλοία) Κάνουν τη μυελινοποίηση (ολιγοδενδροκύτταρα και κύτταρα Schwann) ΛΕΙΤΟΥΡΓΙΕΣ ΤΩΝ ΓΛΟΙΟΚΥΤΤΑΡΩΝ
 Συμμετέχουν στη δημιουργία του αιματεγκεφαλικού φραγμού π.χ. Αστροκύτταρα Καθοδηγούν τους νευράξονες κατά την ανάπτυξη (Ακτινωτή γλοία) Κάνουν τη μυελινοποίηση (ολιγοδενδροκύτταρα και κύτταρα Schwann) ΛΕΙΤΟΥΡΓΙΕΣ ΤΩΝ ΓΛΟΙΟΚΥΤΤΑΡΩΝ.")
41
Τρεις κύριοι τύποι νευρογλοιακών κυττάρων (1) Ολιγοδενδροκύτταρα (2) Κύτταρα Schwann (3) Αστροκύτταρα 1,2 μυελίνωση, μικρά κύτταρα, λιγες αποφυάδες, 1 ολιγο-πολλοί άξονες, 1 Schwann, 1 άξονας 3 τα πιο πολλά, μερικές αποφυάδες σχηματίζουν ποδίσκους στα νεύρα. Άλλα σε επαφή με τα αιμοφόρα αγγεία και αναγκάζουν τα ενδοθηλιακά κύτταρα Να σχηματίσουν στενές συνδέσεις (π.χ. στον αιματεγκεφαλικό φραγμό)
.")
42
ΑΣΤΡΟΚΥΤΤΑΡΑ ΣΕ ΣΥΝΑΨΗ

43
Dendritic spine (S) in the vicinity of neuronal perikaryon (P), showing three postsynaptic densities (asterisks) which belong to only one large perforated synapse. The spine is surrounded by astrocyte processes (A). Note a neuronal subsurface cistern in the proximity of the spine. Scale = 200 nm. (Mouse, neocortex.) ΑΣΤΡΟΚΥΤΤΑΡΑ ΣΕ ΣΥΝΑΨΗ
. Note a neuronal subsurface cistern in the proximity of the spine. Scale = 200 nm. (Mouse, neocortex.) ΑΣΤΡΟΚΥΤΤΑΡΑ ΣΕ ΣΥΝΑΨΗ.")
44
ΑΙΜΑΤΕΓΚΕΦΑΛΙΚΟΣ ΦΡΑΓΜΟΣ

46
Τα Γλοιακά Κύτταρα Καθοδηγούν τους Αναπτυσσόμενους Νευράξονες ΑΚΤΙΝΩΤΗ ΓΛΟΙΑ

51
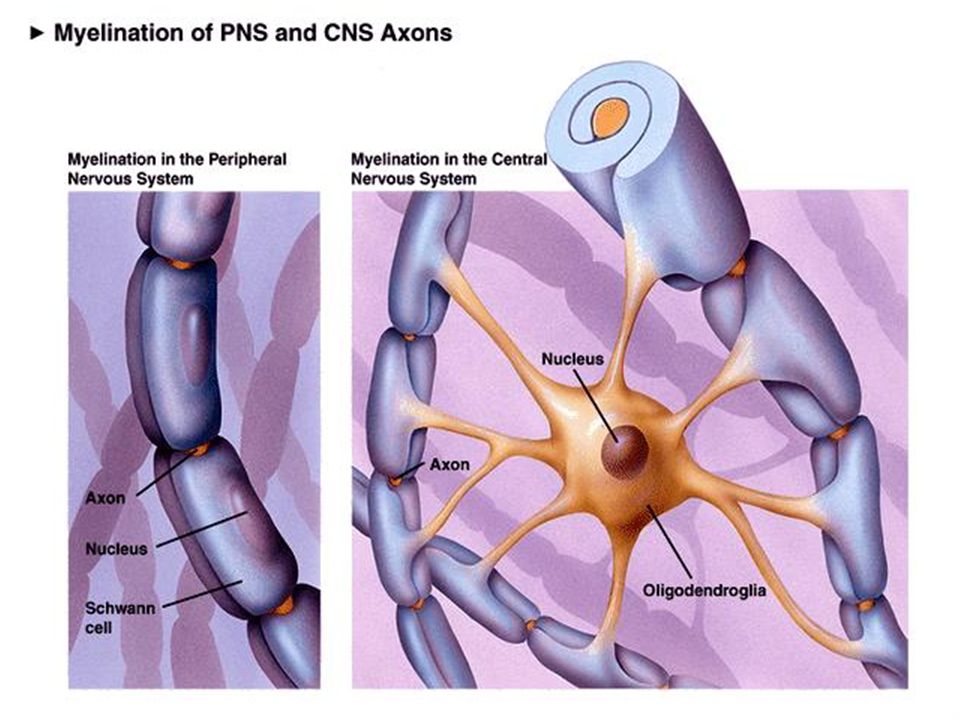
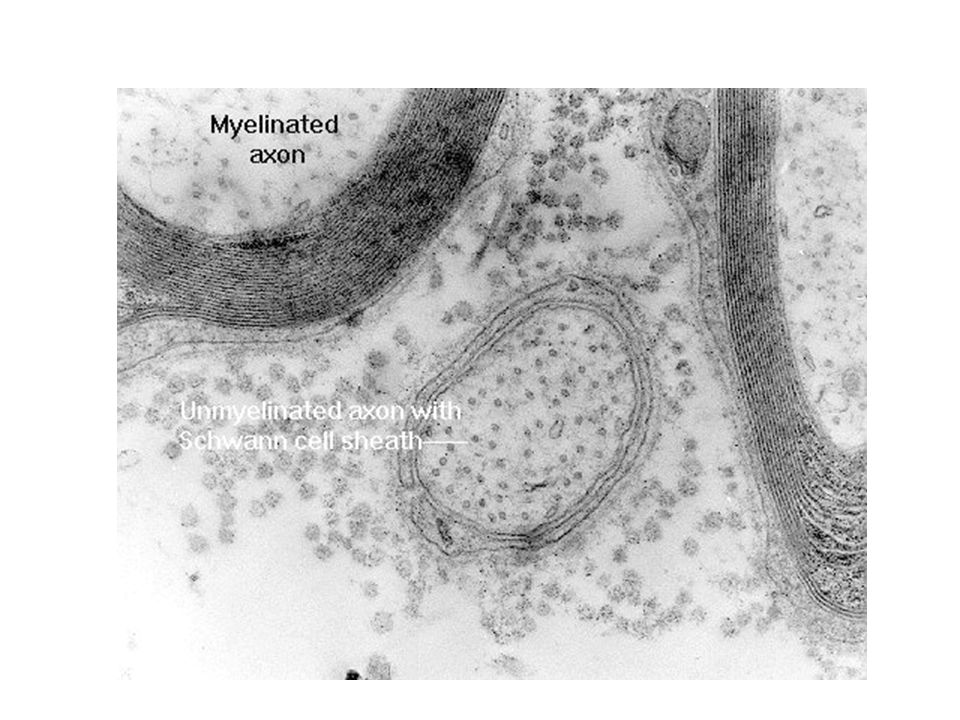
Περισφίξεις του Ranvier

57
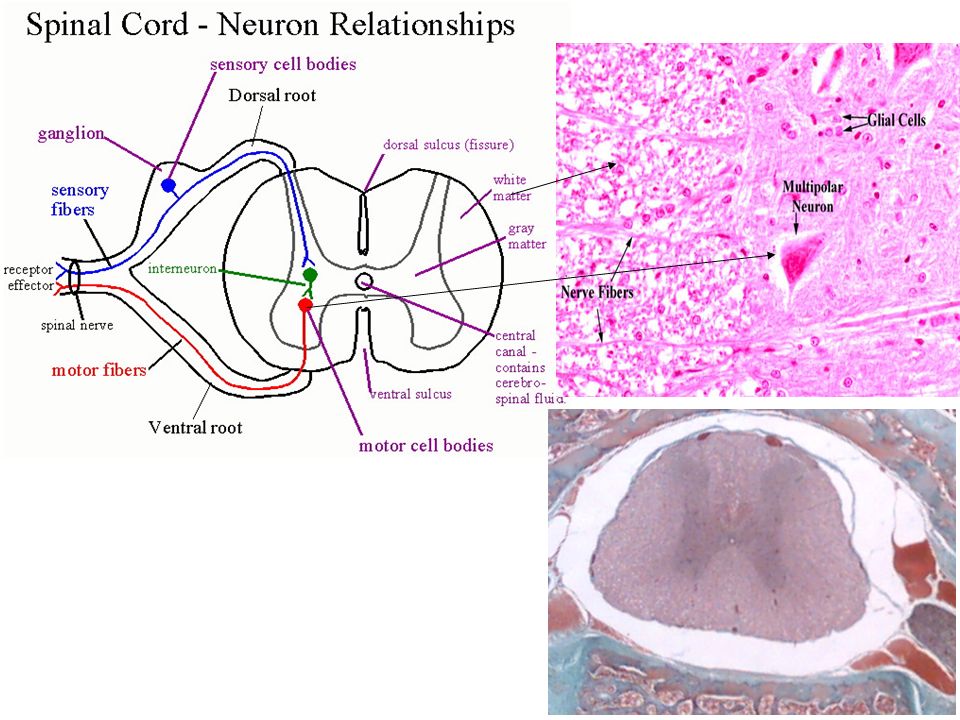
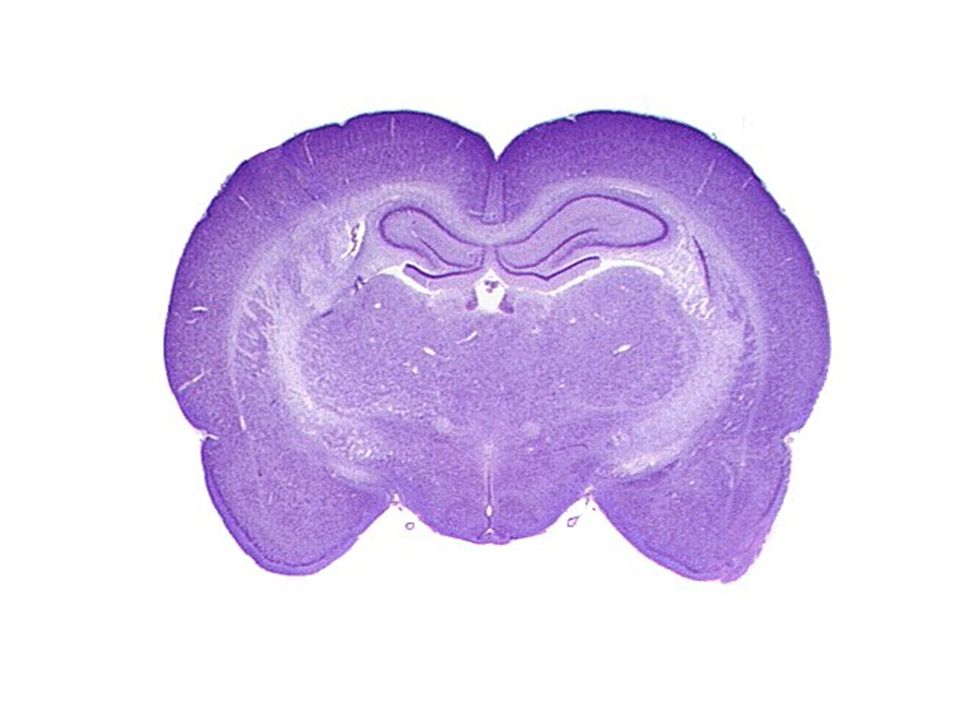
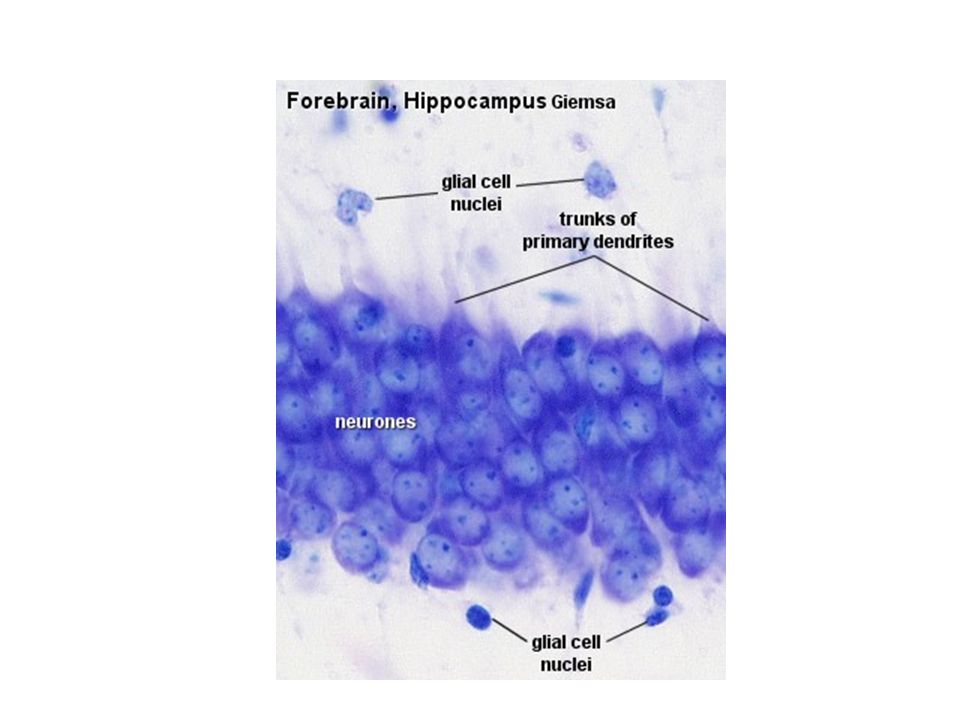
Ο φλοιός των εγκεφαλικών ημισφαιρίων είναι οργανωμένος σε στοιβάδες.

58
Πυραμιδικοί Νευρώνες στον Φλοιό

62
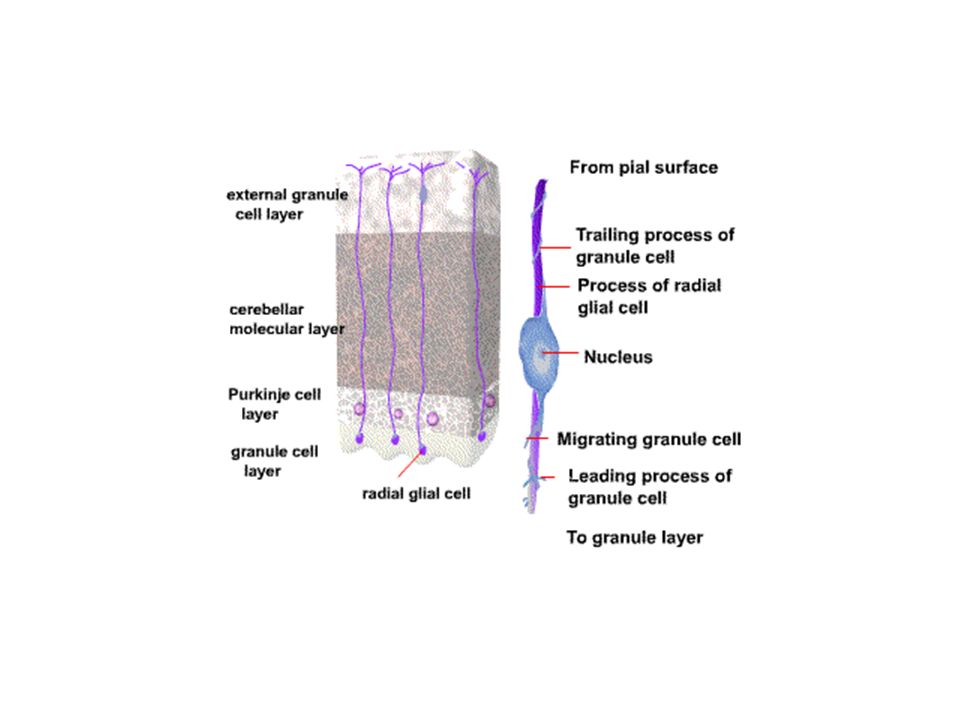
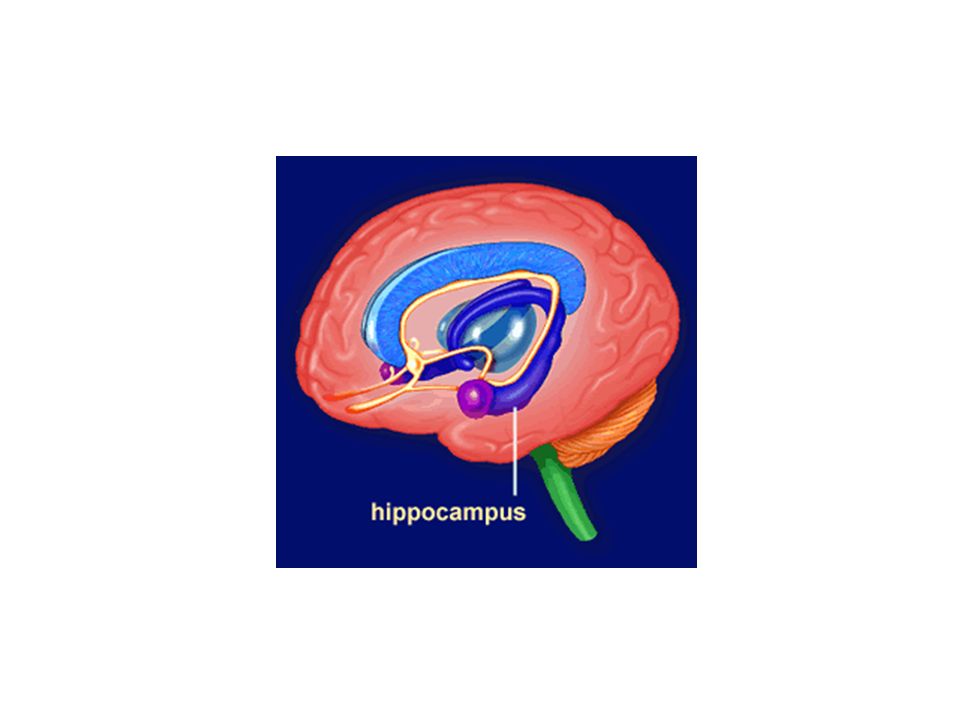
Παρεγκεφαλίδα

63
Μοριώδης στοιβάδα Κοκκιώδης Purkinje Κύτταρα

Παρόμοιες παρουσιάσεις











, Performance Indicators (PIs), Key Performance Indicators (KPIs)>")

 Διδάσκων: Καθηγητής Χρήστος.>")



 Όραση Μαρία Κουτρομάνου. Structure of the Eye: Iris The iris is similar to the diaphragm in a camera Your iris widens in dim light and.>")





