Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
FOTOSINTEZA Istorijat
početak razvoja fiziologije biljaka , kao posebne naučne oblasti, gotovo se potpuno podudara sa istorijom proučavanja fotosinteze to znači da su funkcije, obuhvaćene danas nazivom fotosinteza, bile prve posebne biljne odlike, koje su uopšte zapažene Istraživanja procesa fotosinteze mogu da se podijele na nekoliko perioda Prvi period Drugi period – 1900 Treći period – 1940 Period poslije

2
prvi period Pristley – biljke popravljaju vazduh (eksperiment je pokazao da se u zatvorenom prostoru pod staklenim zvonom svijeća gasi a da se životinje guše od »fiksiranog« vazduha; vazduh se može »popraviti« ako se pod zvono unese zelena biljka)- kiseonik nije bio poznat, Pristley ga je nazvao »deflorgistički vazduh« – Ingen-Housz – pokazao da biljke izdvajaju kiseonik samo na svjetlosti i da je ovaj proces vezan samo za zelene djelove biljke - listove, čime je ukazano na značaj hlorofila – Senébier – ustanovio da biljke ne izdvajaju samo kiseonik već istovremeno usvajaju ugljendioksid tkz. »fiksirani« vazduh. (Lavoisier otkriva u međuvremenu kiseonik i ugljendioksid tako da je Senébier koristio tačne nazive) – Saussure – kvantitativnim mjerenjem pokazao da je količina izdvojenog kiseonika jednaka količini usvojenog ugljendioksida ( poseban značaj- ukazao na značaj vode u ovom procesu, ali sa pogrešnom konstatacijom da izdvojeni kiseonik u procesu fotosinteze potiče od ugljendioksida a ne iz vode) – Mayer – utvrdio zakon o konzervaciji energije, a tri godine kasnije primijenio ovaj zakon u biološkim procesima - znači, polovinom XIX vijeka bilo je poznato da u procesu fotosinteza učestvuju svjetlost, voda i ugljendioksid
- kiseonik nije bio poznat, Pristley ga je nazvao »deflorgistički vazduh« – Ingen-Housz – pokazao da biljke izdvajaju kiseonik samo na svjetlosti i da je ovaj proces vezan samo za zelene djelove biljke - listove, čime je ukazano na značaj hlorofila – Senébier – ustanovio da biljke ne izdvajaju samo kiseonik već istovremeno usvajaju ugljendioksid tkz. »fiksirani« vazduh. (Lavoisier otkriva u međuvremenu kiseonik i ugljendioksid tako da je Senébier koristio tačne nazive) – Saussure – kvantitativnim mjerenjem pokazao da je količina izdvojenog kiseonika jednaka količini usvojenog ugljendioksida ( poseban značaj- ukazao na značaj vode u ovom procesu, ali sa pogrešnom konstatacijom da izdvojeni kiseonik u procesu fotosinteze potiče od ugljendioksida a ne iz vode) – Mayer – utvrdio zakon o konzervaciji energije, a tri godine kasnije primijenio ovaj zakon u biološkim procesima. - znači, polovinom XIX vijeka bilo je poznato da u procesu fotosinteza učestvuju svjetlost, voda i ugljendioksid.")
3
drugi period 1850 – 1900 za ovaj period su značajna istraživanja u oblasti energetike fotosinteze. Utvrđeno je da je proces fotosinteze povezan sa usvajanjem energije i da su hloroplasti jedino odgovorni za ovaj proces Timirjazev- ovom procesu pridaje kosmičku ulogu: »Zeleni list ili tačnije mikroskopsko zeleno zrno hloroplast jeste fokus tačke u svjetlosnom prostranstvu u koju s jedne strane pritiče energije sunca, a sa druge strane, nastaje početak svih pojava života na zemlji« 1862. – Sachs – dokazao obrazovanje skrobnih zrna za vrijeme fotosinteze tj. na svjetlosti i njihovo nestajanje – razgradnju u mraku, što znači da su ugljeni hidrati osnovni produkti fotosinteze – Engleman – pokazao direktnu vezu između izdvajanja kiseonika i hloroplasta i odnos između akcionog spektra fotosinteze i apsorpcionog spektra hlorofila tj. uticaj spektralnog sastava svjetlosti na proces fotosinteze

4
treći period 1900 – 1940 obuhvata istraživanje u oblasti hemije pigmenata, strukture hloroplasta, hromatografije i biohemije fotosinteze 1903. – Cvet – prvo razdvajane pigmenata 1913. – Willstätter i sar. – odredili strukturu hlorofila 1905. – Blackman i Mattaci – predpostavili da se proces fotosinteze sastoji od »svijetlih« i »tamnih« reakcija, što su eksperimentalno potvrdili Emerson i sar. tridesetih godina tridesetih godina dvadesetog vijeka – Van Neil – pokazao da je fotosinteza nekih bakterije slična fotosintezi zelenih biljaka; predpostavio da je porijeklo izdvojenog kiseonika iz vode – Hill – potvdio Neil-ovu predpostavku na izolovanim hloroplastima – koristeći vještačke akceptore elektrona (soli Fe) pokazao je da izolovani hloroplasti izdvajaju O2 u odsustvu CO2 i da za ovaj proces nije neophodno da ćelije budu cijele
 pokazao je da izolovani hloroplasti izdvajaju O2 u odsustvu CO2 i da za ovaj proces nije neophodno da ćelije budu cijele.")
5
period poslije 1940 primjena radioizotopa kao obilježivača, posebno radioaktivnog ugljenika (Ruben i sar. 1939), i primjena papirne hromatografije (Berson, Calvin i dr.) omogućile su da se definiše put ugljenika u procesu fotosinteze Arnon i sar. – izolovani hloroplasti sintetišu ATP na svjetlosti iz ADP i Pi - pokazano je da se konzervacija sunčene energije u hemijsku energiju vrši u lamelama hloroplasta koje sadrže hlorofil i da je ovaj proces nezavisan od drugih enzimskim sistema ili organela kako se do tada predpostavljalo (ova fosforilacija je definisana kao fotofosforilacija ili fotosintetička fosforilacija da bi se razlikovala od oksidativne fosforilacije u mitohondrijama) - razlikujemo klasičan C3 put karboksilacije i C4 put karboksilacije ( npr. kukuruz, šećerna trska kao monokotile, i neke dikotile kao Amerantus edulis)
, i primjena papirne hromatografije (Berson, Calvin i dr.) omogućile su da se definiše put ugljenika u procesu fotosinteze Arnon i sar. – izolovani hloroplasti sintetišu ATP na svjetlosti iz ADP i Pi. - pokazano je da se konzervacija sunčene energije u hemijsku energiju vrši u lamelama hloroplasta koje sadrže hlorofil i da je ovaj proces nezavisan od drugih enzimskim sistema ili organela kako se do tada predpostavljalo (ova fosforilacija je definisana kao fotofosforilacija ili fotosintetička fosforilacija da bi se razlikovala od oksidativne fosforilacije u mitohondrijama) - razlikujemo klasičan C3 put karboksilacije i C4 put karboksilacije ( npr. kukuruz, šećerna trska kao monokotile, i neke dikotile kao Amerantus edulis)")
6
Specifičnost procesa fotosinteze u živom svijetu
- FOTOSINTEZA predstavlja niz reakcija u kojima se pomoću energije svjetlosti iz neogranskih jedinjenja CO2 i vode, sintetišu organska jedinjenja bogata energijom koja je neophodna za održavanje života - ZELENE BILJKE zasad predstavljaju jedini sistem pomoću koga se usvaja ugljenik iz vazduha i koji sunčev zrak, neuhvatljiv za čovijeka i životinje transformiše u hemijsku energiju organskih jedinjenja - prema tome, FOTOSINTEZA je proces bez koga se održavanje života na Zemlji nebi moglo zamisliti - čovijek i životinje u svojoj ishrani koriste gotova organska jedinjenja koja su biljke izgradile Danas se samo neznatan dio fotosintetičke produkcije biljaka koristi kao hrana (npr. - na zemljinoj kugli ima oko 4,4 milijarde hektara šuma odnosno ¼ kopna koje stvaraju oko 36 milijardi tona organske materije – ona se kao hrana (lov, divlji plodovi, pečurke...) koristi najviše u iznosu od 1% - još manje se koristi fotosintetička produkcija vodenih biljaka koje stvaraju godišnje 325 milijardi tona organske materije i nju uglavnom koriste samo ribe) - čovijek mnogo više koristi fotosintetičku produkciju gajenih biljaka i to direktno (voće, povrće...) i ili pad preko posrednika tj. životinja
 koristi najviše u iznosu od 1% - još manje se koristi fotosintetička produkcija vodenih biljaka koje. stvaraju godišnje 325 milijardi tona organske materije i nju uglavnom koriste samo ribe) - čovijek mnogo više koristi fotosintetičku produkciju gajenih biljaka i to direktno (voće, povrće...) i ili pad preko posrednika tj. životinja.")
7
STRUKTURA I HEMIJSKI SASTAV HLOROPLASTA
- HLOROPLASTI se nalaze u svim zelenim djelovima biljke – najviše ih je u palisadnom i sunđerastom parenhimu lista - HLOROPLASTI su FOTOSINTETIČKI APARAT biljke – u njima se vrši proces fotosinteze tj. preobraćanje sunčeve energije i njeno akumiliranje u obliku jedinjenja bogatih energijom HEMIJSKI SASTAV % Bjelančevine Lipidi Ugljeni hidrati Hlorofili Karotenoidi ,5 RNK DNK ,5 Citohrom f ,1 Vitamin K ,004 Vitamin E ,08 Mineralne materije - BJELANČEVINE – nerastvorljivi oblik vezane za lipide i molekule hlorofila - rastvorljivi oblik – enzimi hloroplasta (najvažniji su enzimi elektrontransportnog lanca, tamne faze fotosinteze, proteini membrana tj. strukturni proteini) - mineralni sastav- u tkivima lista i hloroplastima je veoma različit ( 80% ukupne količine Fe se nalazi u hloroplastima)
 - mineralni sastav- u tkivima lista i hloroplastima je veoma različit ( 80% ukupne količine Fe se nalazi u hloroplastima)")
8
MORFOLOGIJA HLOROPLASTA
- HLOROPLASTI se razlikuju po obliku, veličini, broju, kao i po vrsti pigmenata koje sadrže tipični hloroplast viših biljka ima ovalan ili elipsoidan oblik, dužine 4-6 µm, a debljine 1-2 µm kod nižih biljaka – algi oblik može biti veoma različit i služi kao jedan od taksonomskih pokazatelja broj hloroplasta u ćeliji varira i zavisi od vrste biljaka i tkiva u kome se nalaze (u palisadnim ćelijama , dok u ćelijama sunđerastog parenhima samo nekoliko) – zato i masa hloroplasta prema masi lista varira – obično 20-30%
 – zato i masa hloroplasta prema masi lista varira – obično 20-30%")
9
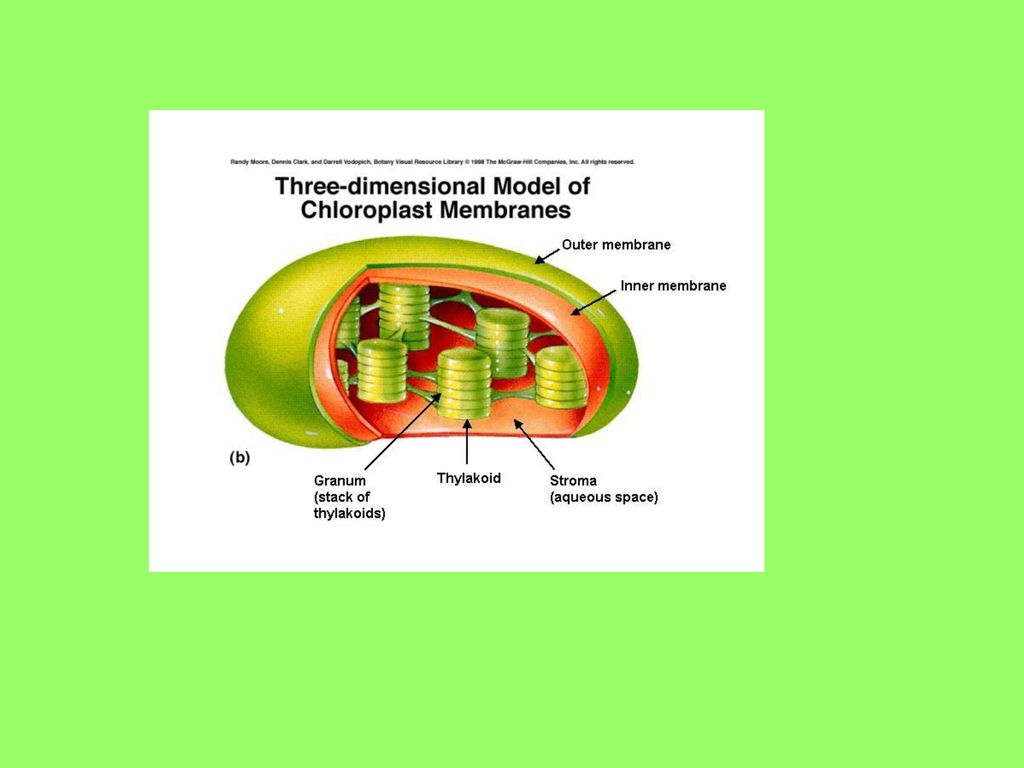
ULTRASTRUKTURA HLOROPLASTA
- spolja – dvojna membrana - unutrašnjost - ispunjena sadržajem koji se naziva STROMA u STROMI se nalazi veći ili manji broj lamela koje grade spljoštene vezikule nazvane TILAKOIDI na pojedinim mjestima tilakoidi su agregirani tj. složeni jedan iznad drugog u vidu paketića tkz. GRANE HLOROPLASTA koje se pod svjetlosnim mikroskopom ocrtavaju kao zrnca, po čemu su ove tvorevine dobile ime ( granum-zrno) pojedini tilakoidi protežu se kroz nekoliko grana i čine tkz. STROMA TILAKOIDE, na ovaj način tilakoidi sistem je povezana cjelina - pored granuralne strukture neke alge i biljke sa tkz. »Kranc« anatomijom lista imaju agranalne hloroplaste – kod viših biljaka oko sprovodnih snopića – parenhimskoj sari – oni sadrže samo lamele tilakoida koje su iste debljine i paralelno se pružaju po dužini hloroplasta (postojenje dimorfnih hloroplasta je prvo pokazano kod kukuruza)
 pojedini tilakoidi protežu se kroz nekoliko grana i čine tkz. STROMA TILAKOIDE, na ovaj način tilakoidi sistem je povezana cjelina. - pored granuralne strukture neke alge i biljke sa tkz. »Kranc« anatomijom lista imaju agranalne hloroplaste – kod viših biljaka oko sprovodnih snopića – parenhimskoj sari – oni sadrže samo lamele tilakoida koje su iste debljine i paralelno se pružaju po dužini hloroplasta (postojenje dimorfnih hloroplasta je prvo pokazano kod kukuruza)")
11
- sve MEMBRANE koje ulaze u sastav hloroplasta - tečni mozaični model (lipidni dvosloj sa hidrofilnim glavama na površini i hidrofobnim lancima masnih kiselina u sredini; integrisani tj. unutrašnji i asocirani tj. spoljašnji proteini koji nose lance oligosaharida) OMOTAČ – SPOLJAŠNJA membrana je glatka, a UNUTRAŠNJA može biti manje ili više naborana - nabori su invaginacije koje se sastoje od razgranatih tubula gradeći tkz. PERIFERNI RETIKULUM HLOROPLASTA (PRH je posebno razvijen kod C4 biljaka i ima važnu ulogu u transportu metabolita između hloroplasta i okolne sredine) - spoljašnje membrana je više propustljiva za određene hemijske komponente, dok je unutrašnja više semipermeabilna
 - spoljašnje membrana je više propustljiva za određene hemijske komponente, dok je unutrašnja više semipermeabilna.")
12
MEMBRANE TILAKOIDA – se znatno razlikuju jer se lipidi koji ulaze u njihov sastav veoma rijetko se nalaze u nefotosintetičkim biološkim membranama - acil-lipidna komponenta čini 35% težine membrane – u njoj 70% čine galaktolipidi i 10% sulfolipidi znatan dio ne-acilnih lipida čine pigmenti i kinoni hlorofil čini 20% svih lipidnih jedinjenja masne kiseline u sastavu lipida su izuzetno visokoj mjeri nezasićene, a pošto i sterola ima malo (0,5%) membrane tilakoida imaju najveću moguću fluidnost i omogućavaju visoku lateralnu pokretljivost integrisanih proteina proteini čine 50% membrane, sastoje se uglavnom od više polipeptida, koji nastaju u hloroplastu ili citoplazmi - pri formiranju membrana oni se sakupljaju i obrazuju jedinstvene komplekse - proteini u kompleksima imaju različite enzimske funkcije, ali prvenstveno služe kao NOSAČI, za koje su na specifičnim mjestima vezani pigmenti i prenosioci elektrona - ovim vezivanjem je obezbjeđena potrebna distanca između dva prenosioca, kao i njihova orjentacija, tako da se prenos svjetlosne energije ili elektrona kroz membranu može obavljati po utvrđenom redosljedu i pravcu
 membrane tilakoida imaju najveću moguću fluidnost i omogućavaju visoku lateralnu pokretljivost integrisanih proteina. proteini čine 50% membrane, sastoje se uglavnom od više polipeptida, koji nastaju u hloroplastu ili citoplazmi. - pri formiranju membrana oni se sakupljaju i obrazuju jedinstvene komplekse. - proteini u kompleksima imaju različite enzimske funkcije, ali prvenstveno služe kao NOSAČI, za koje su na specifičnim mjestima vezani pigmenti i prenosioci elektrona. - ovim vezivanjem je obezbjeđena potrebna distanca između dva prenosioca, kao i njihova orjentacija, tako da se prenos svjetlosne energije ili elektrona kroz membranu može obavljati po utvrđenom redosljedu i pravcu.")
13
- moderne metode u elektronskoj mikroskopiji – duboko zamrzavanje, cijepanje i sječenje u vakumu – mogu se posmatrati površine membrana - spoljašnja prema stromi - unutrašnja prema lumenu tilakoida - dvije površine koje postaju dostupne pošto membrana pod pritiskom prsne duž hidrofobnog dijela dvojnog lipidnog sloja pošto membrane tilakoida grana i tilakoida strome nisu iste na preparatima može da se razlikuje osam različitih površina – tako su dobijene informacije o rasporedu proteinskih globula u membranama, o njihovoj veličini, obliku i učestalostu - potvrđeno je da su membrane asimetrične što je bitno za obavljanje njihovih funkcija

14
FORMIRANJE TILAKOIDA – ranije se smatralo da nastaju invaginacijom unutrašnje membrane omotača
ali, membrane tilakoida se po hemijskom sastavu razlikuju od membrana omotača – smatra se da unutrašnji membranski sistem hloroplasta nastaje rastenjem primarnih lamela proplastida lamele su po ivici perforirane i na obodu perforacija se pojavljuju izraštaji koji se proširuju i obrazuju spljoštenu vezikulu – tilakoid oni se slažu jedan preko drugog i obrazuju grane, ili se izdužuju kroz stromu i povezuju sa susjednim naslagama tilakoida formiranje je završeno kada se u njih ugrade kompleksi proteina i pigmenata - pigmenti se izgrađuju u stromi, a većina proteina u citoplazmi i oni ulaze posebnim mehanizmima u hloroplaste pri njihovom formiranju

15
SASTAV STROME – to je rastvor sličan citosolu
u njoj se nalaze brojni ribozomi, plastoglobule, skrobna zrna, niti hloroplastne DNK, RNK, kao i brojni enzimi (za odvijanje Krebsovog ciklusa, sintezu proteina...) ribozomi su manji od onih u citoplazmi- tip 70 S i veličina 15-20nm u zavisnosti od starosti listova u hloroplastima može da se uoči manji ili veći broj plastoglobula – osmiofilna tjelašca veličine nm - plastoglobule uglavnom sadrže lipide i lipoproteine tj. komponente za izgradnju membrana tilakoida broj skrobnih zrna može biti pokazatelj fiziološke aktivnosti aktivnosti hloroplasta najviše je zastupljen enzim ribuloza-bifosfat karboksilaza/oksigenaza jedan od najraširenijih proteina na našoj planeti
 ribozomi su manji od onih u citoplazmi- tip 70 S i veličina 15-20nm. u zavisnosti od starosti listova u hloroplastima može da se uoči manji ili veći broj plastoglobula – osmiofilna tjelašca veličine nm. - plastoglobule uglavnom sadrže lipide i lipoproteine tj. komponente za izgradnju membrana tilakoida. broj skrobnih zrna može biti pokazatelj fiziološke aktivnosti aktivnosti hloroplasta. najviše je zastupljen enzim ribuloza-bifosfat karboksilaza/oksigenaza jedan od najraširenijih proteina na našoj planeti.")
16
KOMPARMENTI U HLOROPLASTU – tri odvojena prostora
- periplastidijalni prostor – prostor između spoljašnje i unutrašnje membrane – reguliše se transport između hloroplasta i citosola - lumen svih tilakoida – vjerovatno je jedinstven u jednom hloroplastu - stroma – tilakoidni sistem je suspendovan komparmentni su razdvojeni jedan od drugog jer su membrane nepropustljive za rastvorene supstance i jone ali, specifični proteini regulišu transport u oba pravca, tako da postoji neprekidna razmjena svih produkata fotosinteze, drugih metabolita i jona među kompartmentima kao i između hloroplasta i citosola

17
FOTOSINTETIČKI PIGMENTI
svjetlost može biti aktivna u fotohemijskim procesima samo ako je apsorbovana apsorpciju svjetlosti određenih talasnih dužina vrše pigmenti pigmenti učestvuju u procesu fotosinteze, ali time se ne iscrpljuje njihova uloga u životu biljaka u izvjesnoj mjeri oni učestvuju u procesima rastenja i razvića biljaka npr. ponici biljaka gajeni u uslovima tame su etiolirani i duži nego oni gajenji na svjetlosti – ova pojava ima veliki biološki značaj i može biti objašnjena procesima koji zavise od prisustva hlorofila - procesi fotoperiodizma, sa kojima je povezano proticanje svjetlosnog stadijuma, pokazali su se zavisnim od pigmentno-bjelančevinastog kompleksa koji ima sposobnost apsorpcije kako u crvenom tako i u bliskom infracrvenom dijelu spektra - U životu životinja pigmenti imaju značajnu ulogu kao provitamini. Hlorofil se može smatrati provitaminom K, a karoten provitaminom A. Neki ksantofili učestvuju u reakcijama koje se događaju pri radu čula vida.

18
U hloroplastima se nalaze TRI GRUPE pigmenata:
HLOROFILI ima ih oko 10, to su jedinjenja zelene boje, KAROTENOIDI ima ih oko 80 i to 6-8 KAROTENA i preko 70 KSANTOFILA, koji su žute, naranđaste ili crvenkaste boje FIKOBILINI ima 2, a boje su plave ili crvene.

19
HLOROFIL Nalazi se u tilakoidima hloroplasta raznih organa i tkiva biljaka. Nije primjećen u plastidima epidermalnih ćelija (izuzev ćelija koje zatvaraju stome). Može se često naći u parenhimskim ćelijama mladih grana i grančica, a kod nekih biljaka i u felodermu stabla. Nekad se čak nalazi i u korijenu koji se razvija ili nalazi na svjetlosti. Često se primjećuje u organima cvijeta, mladih plodova, a nekad čak i u klici sjemena. Poznati hlorofili kod raznih biljnih grupa i fotosintetizujućih bakterija Modrozelene alge
. Može se često naći u parenhimskim ćelijama mladih grana i grančica, a kod nekih biljaka i u felodermu stabla. Nekad se čak nalazi i u korijenu koji se razvija ili nalazi na svjetlosti. Često se primjećuje u organima cvijeta, mladih plodova, a nekad čak i u klici sjemena. Poznati hlorofili kod raznih biljnih grupa i fotosintetizujućih bakterija. Modrozelene alge.")
20
Kod viših biljaka su zastupljeni hlorofil a i b
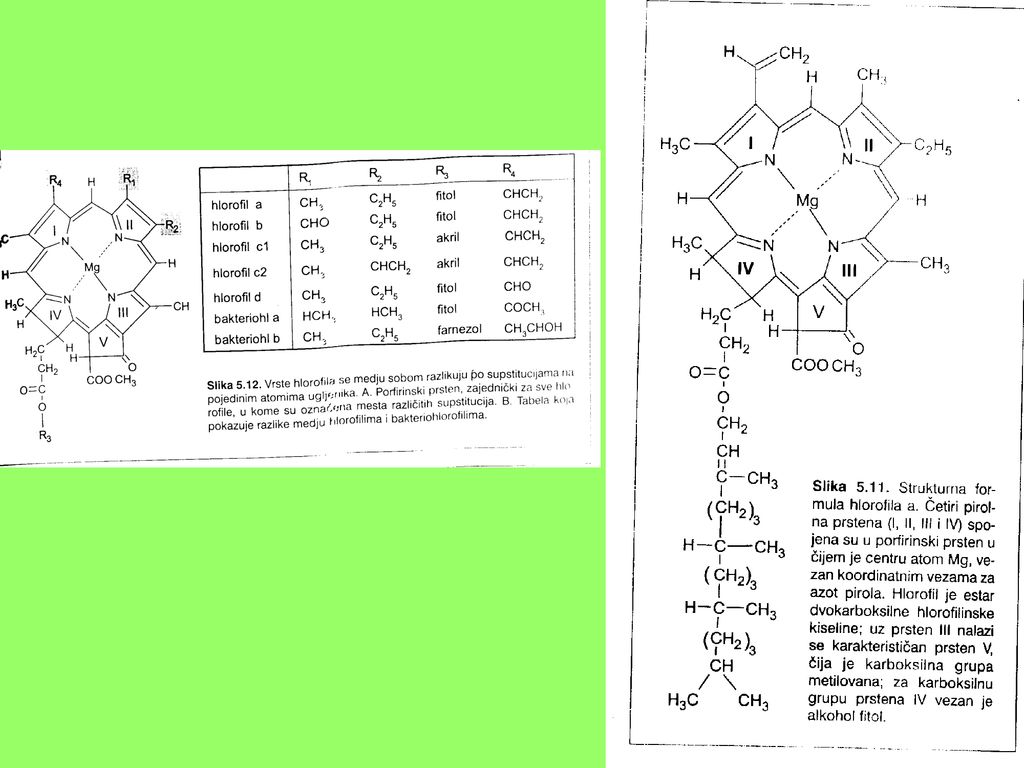
Hl a - C55H72O5N4Mg – molekulska masa 893 Hl b – C55H70O5N4Mg – molekulska masa 907 Po hemijskoj strukturi hlorofili su derivati cikličnih tetrapirola koji grade porfirinski prsten i srodni su sa drugim porfirinskim jedinjenjima, kao što je hem. Hlorofili a i b su estri dikarboksilne kiseline – hlorofilinske kiseline tj. hlorofilina, gdje je vodonik u jednoj karboksilnoj grupi zamjenjen ostatkom metilalkohola CH3OH, a u drugoj ostatkom alkohola fitola

22
Biosinteza hlorofila 1.Početno jedinjenje u sintezi porfirina je 5-aminolevulinska kiselina (ALA) Ona postaje na dva načina: u animalnim ćelijama, u ćelijama gljiva, kvasaca i nakih bakterija (Rhodobacter, Rhodospirillum, Rhizobium) ALA nastaje kondezacijom sukcinil-CoA i aminokiseline glicina, pod dejstvom enzima ALA-sintetaze uz odvajane CO2
 ALA nastaje kondezacijom sukcinil-CoA i aminokiseline glicina, pod dejstvom enzima ALA-sintetaze uz odvajane CO2.")
23
Kod viših biljaka, mahovina, mnogih Eubacteria i Cyanobacteria ona nastaje od intaktnog molekula glutaminske kiseline -glutamat tkz C5 put koja se vezuje za transfernu RNK glutamat glutamat semialdehidna aminotransferaza- mijenja mjesto amino grupe reduktaza sintetaza

24
2. Procesi od 5-aminolevulinske kiseline do protoporfirina IX
kondenzacija dva molekula ALA-nastaje prvo ciklično jedinjenje sa petočlanim prstenom – porfobilinogen – enzim porfobilinogen sintaza (ili ALA dehidrataza) - porfobilinogen je pirol, heterociklično jedinjenje, sa N u prstenu i slobodnim ostacima sirćetne i propionske kiseline - zatim se 4 molekula porfobilinogena kondezuju i obrazuju porfirinski prsten – enzim porfobilinogen deaminaza i uroporfinogen sintaza – prvo porfirinsko jedinjenje je uroporfirinogen III
 - porfobilinogen je pirol, heterociklično jedinjenje, sa N u prstenu i slobodnim ostacima sirćetne i propionske kiseline. - zatim se 4 molekula porfobilinogena kondezuju i obrazuju porfirinski prsten – enzim porfobilinogen deaminaza i uroporfinogen sintaza – prvo porfirinsko jedinjenje je uroporfirinogen III.")
25
prvo porfirinsko jedinjenje, koje nastaje kondenzacijom 4 porfobilinogena

26
- daljih nekoliko stupnjeva se dešava uglavnom na bočnim kiselinskim ostacima uroporfirinogena III:
dekarboksilacija - koproporfirinogen III oksidativna dekarboksilacija 2 lanca propionske kiseline dobijaju se dvije vinil grupe- protoporfirinogen IX - oksidacija gubitkom 6 protona i elektrona – protoporfirin IX – posljednji zajedički prekursor svih porfirina - prima atom metala u centar prstena; ako se ugrađuje Mg nastaje Mg-protoporfirin od koga nastaju hlorofili - ako se ugrađuje Fe nastaje hem koji je prostetična grupa hemoglobina, različitih citohroma, enzima katalaze, peroksidaze...; - ako dođe do raskidanja prstena bez vezivanja metala čime se dobija linearni niz 4 pirolova prstena (tako u životinjskim ćelijama nastaju prostetične grupe žučnih pigmenata – bilini; kod modrozelenih i crvenih algi pigmenti fikobilini; kod svih viših i nižih biljaka prostetična grupa pigmenta fitohroma

27
3. Procesi od Mg-protoporfirina do hlorofila
- promjene na bočnim lancima prstena III: - esterifikacija ostataka propionske kiseline metil grupom obrazuje se peti izociklični prsten – odlika hlorofila redukcija bočnog vinil lanca na prstenu II – postaje protohlorofilid a (od hlorofila a se razlikuje po tome što nema fitol i što prsten IV nije redukovan nego ima nezasićenu vezu) - redukcijom na prstenu IV nastaje hlorofilid a , ali se prema načinu redukcije biljke dijele na dvije velike grupe. Fotosintetičke bakterije, alge, mahovine i paprati mogu ovaj proces obavljati u mraku. Angiosperme mogu da obavljaju redukciju protohlorofilida a isključivo na svjetlosti, a kod gimnospermi se u tom pogledu razlikuju embrionalni i adultni listovi. U kotiledonima gimnospermi se redukcija obavlja u mraku, ali je svjetlost neophodna za redukciju u svim drugim listovima. - dodaje se fitolna grupa i nastaje hlorofil a a njegovom oksidacijom nastaje hlorofil b
 - redukcijom na prstenu IV nastaje hlorofilid a , ali se prema načinu redukcije biljke dijele na dvije velike grupe. Fotosintetičke bakterije, alge, mahovine i paprati mogu ovaj proces obavljati u mraku. Angiosperme mogu da obavljaju redukciju protohlorofilida a isključivo na svjetlosti, a kod gimnospermi se u tom pogledu razlikuju embrionalni i adultni listovi. U kotiledonima gimnospermi se redukcija obavlja u mraku, ali je svjetlost neophodna za redukciju u svim drugim listovima. - dodaje se fitolna grupa i nastaje hlorofil a a njegovom oksidacijom nastaje hlorofil b.")
28
KAROTENOIDI rasprostranjenost karotenoida zavisi od mnogih bioloških osobina biljaka kao i od njihovih organa u listovima viših biljaka nalaze se: i karoteni, lutein, zaeksantin, violaksantin, neoksantin, dok se u bakterijama, gljivama i algama nalazi i veliki br. drugih karotenoida dijele se u dvije grupe: karotene, koji su po hemijskom sastavu ugljovodonici i ksantofile, koji pored ugljenika i vodonika sadrže kiseonik nalaze se u hloroplastima – učestvuju u fotosintezi i u hromoplastima – daju žutu, narandžastu ili crvenu boju raznim cvjetovima, plodovima i dr djelovima biljke

29
nalaze se manje više u svim organima biljke
podzemni organi šećerne repe, mrkve, krompira i dr. sadrže čitav niz karotenoida (najvažniji izvor karotenoida je korjen mrkve i slatkog krompira, osnovni karotenoid u korjenu mrkve je karoten, a manje i kao i lutein) u listovima se nalaze zajedno sa zelenim pigmentima u hloroplastima (najrasprostranjeniji je karoten, a manje . Dosad nije nađen likopen i karoten,izuzetak je vilina kosica gdje je karoten osnovni karoten) u plodovima su najviše zastupljeni likopeni i karoten (u plodovima se mogu susresti predstavnici karotenoida raznovrsniji u odnosu na listove zelenih biljaka, pored karotena tu su rodoksantin, likopen, lipoksantin, rubiksantin, teraksantin, mutatohrom, kapsantin, celaksantin i anteraksantin) - u cvjetovima pored karotenoida koji se sreću u listovima nalaze se i specijalni karotenoidi: ešelaksantin, petaloksantin, auroksantin, hrizantemaksantin, rubihrom i dr. - u prašniku i polenu se nalaze u prvom redu karoteni i lutein.
 u listovima se nalaze zajedno sa zelenim pigmentima u hloroplastima (najrasprostranjeniji je karoten, a manje . Dosad nije nađen likopen i karoten,izuzetak je vilina kosica gdje je karoten osnovni karoten) u plodovima su najviše zastupljeni likopeni i karoten (u plodovima se mogu susresti predstavnici karotenoida raznovrsniji u odnosu na listove zelenih biljaka, pored karotena tu su rodoksantin, likopen, lipoksantin, rubiksantin, teraksantin, mutatohrom, kapsantin, celaksantin i anteraksantin) - u cvjetovima pored karotenoida koji se sreću u listovima nalaze se i specijalni karotenoidi: ešelaksantin, petaloksantin, auroksantin, hrizantemaksantin, rubihrom i dr. - u prašniku i polenu se nalaze u prvom redu karoteni i lutein.")
30
Uopšte govoreći oni imaju žutu, narandžastu ili crvenu boju.
Karoteni , i C40H mol. masa Likopin C40H mol. masa Lutein C40H56O mol. masa Zeaksantin C40H56O mol. masa Flavoksantin C40H56O mol. masa Neoksantin C40H56O mol. masa i dr. Pojedini karotenoidi imaju iste skupne formule ali se razlikuju po broju dvogubih veza odnosno po strukturnim formulama – zato se različito ponašaju prema apsorpciji svjetlosti. Utvrđeno je postojenje različitih karotenoida, a za oko 70 je proučena strukturna formula. Svi karotenoidi su po strukturi bliski – čine derivat istog kostura C40 fitoena koji se sastoji od 8 izoprenskih jedinica i koji nastaje dimerizacijom dvije jedinice C20 (geranil geranil pirofosfat).
.")
31
Biosinteza izoprenoidnih jedinjenja
u osnovi izoprenoidnih jedinjenja je IZOPREN – zajednička gradivna jedinica IZOPREN ne postoji slobodan nego kao kao 5- C segment izopentenil-PP ugrađuje u ova jedinjenja izopren izopentenil pirofosfat - IPP nastaje različito u citosolu i hloroplastima u citosolu kondenzacijom 3 molekula acetil CoA nastaje mevalonska kiselina, koja se uz učešće 3 ATP fosforiliše i dekarboksiliše i nastaje IPP u hloroplastima se pod dejstvom transketolaze kondezuje dvokarbonski ostatak piruvata sa 3-gliceraldehid fosfatom, nastaje jedan prekursor, a zatim IPP IPP i njegov izomer DMAPP su hemiterpeni od kojih postaje gas izopren, koji se izlučuje iz listova u atmosferu imaju jedan račvast kraj “glavu” i fosforilovan kraj “rep” kondezacija po principu vezivanja glava-rep biogeno izoprensko pravilo

32
Geranil geranil PP x 2 dimerizacijom GGPP dobijaju se tetraterpeni, među kojima je i fitoen, neposredni prekursor karotenoida većina karotenoida se karakteriše ciklizacijom na jednom ili oba kraja lanca, čime se formira jonski prsten karotenoidi se dijele na karotene (samo C i H) i ksantofile koji nastaju vezivanjem OH grupe (C,H i O)
 i ksantofile koji nastaju vezivanjem OH grupe (C,H i O)")
33
Funkcija karotenoida u hloroplastima učestvuju u apsorpciji svjetlosti, koja se koristi u fotosintezi, ali imaju i druge funkcije kada je list izložen svjetlosti visokog intenziteta, količina apsorbovane energije je često veća od one koja može da se iskoristi u procesu fotosinteze karotenoidi imaju ulogu u gašenju te energije (poznati su mutanti nekih algi i bakterija koji nemaju karotene i koji na svjetlosti brzo gube hlorofil, pa nisu sposobni za autotrofan način života) - postoje neki herbicidi, kao što je norfurazon, koji inhibiraju sintezu karotenoida (biljke koje su njime tretirane u mraku ne razlikuju se od kontrolnih, ali na svjetlosti gube hlorofil i ne preživljavaju) - imaju značajne funkcije u apsorpciji svjetlosti koja izaziva lokomotorne pokrete – fototaksije - možda učestvuju u prenošenju drugih svjetlosnih signala – npr. u fotoperiodizmu - boja mnogih cvjetova je uslovljena njihovim prisustvom – uloga u polinaciji, razmnožavanju i dr. - korišćenje u ishrani biljaka koje sadrže karoten značajno jer je on prekursor vitamina A
 - postoje neki herbicidi, kao što je norfurazon, koji inhibiraju sintezu karotenoida (biljke koje su njime tretirane u mraku ne razlikuju se od kontrolnih, ali na svjetlosti gube hlorofil i ne preživljavaju) - imaju značajne funkcije u apsorpciji svjetlosti koja izaziva lokomotorne pokrete – fototaksije. - možda učestvuju u prenošenju drugih svjetlosnih signala – npr. u fotoperiodizmu. - boja mnogih cvjetova je uslovljena njihovim prisustvom – uloga u polinaciji, razmnožavanju i dr. - korišćenje u ishrani biljaka koje sadrže karoten značajno jer je on prekursor vitamina A.")
34
FIKOBILINI - samo kod modrozelenih i crvenih algi rastvorljivi u vodi i sastoje se od proteina i prostetične grupe koja je hromofor- nosilac boje Proteinski dio molekula je izgrađen od dva peptidna lanca (α i β), sa molekulskom masom i D i za njega su kovalentno vezana 1-2 molekula hromofora - hromofori su fikobilini – jedinjenja sa 4 pirolova prstena povezana u otvoren niz koji nastaje od protoporfirina IX - fikobiliproteini se nalaze u naročitim globulama – fikobilozomima i vezane su spolja za tilakoidne membrane postoji više fikobilina: fikocijanini – plavozelene boje kod Cyanobacteria i fikoeritrini – crvene boje kod Rhodophyta
, sa molekulskom masom i D i za njega su kovalentno vezana 1-2 molekula hromofora. - hromofori su fikobilini – jedinjenja sa 4 pirolova prstena povezana u otvoren niz koji nastaje od protoporfirina IX. - fikobiliproteini se nalaze u naročitim globulama – fikobilozomima i vezane su spolja za tilakoidne membrane. postoji više fikobilina: fikocijanini – plavozelene boje kod Cyanobacteria i fikoeritrini – crvene boje kod Rhodophyta.")
35
ZNAČAJ SUNČEVE SVJETLOSTI ZA PROCES FOTOSINTEZE
Za obavljanje procesa fotosinteze neophodna je svjetlost. Ali, SUNČEVA SVJETLOST nije homogena već se sastoji os svjetlosnih talasa različitih talasnih dužina. Na zemljinu površinu padaju zraci sunca čija je talasna dužina različita. Ljudsko oko je osjetljivo samo na talasne dužine između 380 i 750 nm - to je vidljivi spektar. Smješa svih talasnih dužina u vidljivom polja ima intenzitet poput onog dobijenog od tijela na temperaturi bijelog žara to je bijela svjetlost. Ako svjetlost, koja pada na mrežnjaču oka ne sadrži sve talasne dužine vidljivog spektra, ili ako je intenzitet neke talasne dužine smanjen, govorimo o osjetljivosti oka za boje.

36
UKUPNA SUNČEVA RADIJACIJA
40% - vidljivi dio spektra 51% - infracrveni dio spektra – može da je usvaja voda u biljci 9% ultraljubičasti dio spektra – može da ga usvajaju neke organske komponente FOTOSINTEZA ZELENIH BILJAKA – u granicama vidljivog spektra PURPURNE BAK: – 950 nm ZELENE BAK: - između zelenih bilj. i pur. bak.

37
Bez obzira o kom dijelu spektra se radi, njegova ENERGIJA se izražava
PLANKOVOM jednačinom E= h = h c / h – konstanta Planka 6,625 x J/s c – brzina svjetlosti 3 x 1010 cm/s - talasna dužina svjetlosti 10-9 cm - frekvencija odgovarajuće svjetlosti s-1 FOTOHEMIJSKI EKVIVALENT POJEDINAČNOG MOLEKULA će reagovati samo poslije apsorpcije energije jednog fotona (h ) – zakon Anštajna Stoga, jedan mol (gram mol) nekog jedinjenja mora apsorbovati N fotona (N= Avogardov broj = 6,024 x 1023) tj. N h da bi reakcija startovala Zato se i energija apsorbovanih fotona od strane jednog molekula nekog jedinjenja naziva ANŠTAJN (E) ili MOL ENERGIJE.
 – zakon Anštajna. Stoga, jedan mol (gram mol) nekog jedinjenja mora apsorbovati N fotona (N= Avogardov broj = 6,024 x 1023) tj. N h da bi reakcija startovala. Zato se i energija apsorbovanih fotona od strane jednog molekula nekog jedinjenja naziva ANŠTAJN (E) ili MOL ENERGIJE.")
38
U granicama vidljivog dijela spektra nalazi se svjetlost različitog kvaliteta – ona različito utiče na proces fotosinteze. Naime, svjetlost je sastavljena iz sitnih čestica koje se nazivaju FOTONI i oni izazivaju FOTOHEMIJSKE REAKCIJE. ENERGIJA koju nosi jedan FOTON naziva se KVANT. Ako je TALASNA DUŽINA svjetlosti MANJA utoliko je ENERGIJA VEĆA FOTOHEMIJSKA REAKCIJA može da se odvija samo u slučaju ako energija kvanta nije manja od energije koja je neophodna da bi se savladala ENERGETSKA BARIJERA REAKCIJE – FOTOELEKTRIČNI PRAG. Zato su zraci KRATKIH koji nose VEĆU E KVANTA FOTOHEMIJSKI NAJAKTIVNIJI ( a zraci DUGIH koji imaju MANJU E KVANTA su fotohemijski slabo aktivni. ALI, oni se mnogo usvajaju od strane hlorofila zelenih biljaka i obezbjeđuju fotohemijsku reakciju fotosinteze)
")
39
Često se kaže da se svjetlost apsorbuje od strane biljke ili njenih organa a specijalno lista što nije korektno – svjetlost se usvaja FOTORECEPTORIMA a to su molekuli, kompleksi, atomi, joni i radikali a ne biološke strukture kao što su jedra, mitohondrije, hloroplasti i dr. FOTORECEPTORI su komponente ćelije koje su sposobne da selektivno usvajaju svjetlost pojedinih djelova sunčevo9g spektra u oblasti od nm PORED VRSTE bitna je i koncentracija FOTORECEPTORA, jer je usvajanje svjetlosti rezultat »slučajnog susreta« kvanta svjetlosti i fotoreceptora. Pri istom broju kvanata vjerovatnoća susreta je veća ako je koncentracija fotoreceptora veća.

40
APSORPCIONI SPEKTRI PIGMENATA
fotosintetički pigmenti apsorbuju svjetlost određenih talasnih dužina i stoga imaju boju propuštene ili odbijene svjetlosti. svi hlorofili imaju dvije zone apsorpcije, jednu u plavom a drugu u crvenom dijelu spektra, ali su talasne dužine maksimalne apsorpcije različite hlorofil a u eterskom rastvoru apsorbuje najviše svjetlosti od 430 i 663 nm, a hlorofil b na 450 i 643 nm karotenoidi apsorbuju samo u ljubičastom i plavom dijelu spektra za sve njih je karakterističan spektar sa dva ili tri vrha - karoten ima plato na 420 nm i vrhove na 460 i 480 nm - fukoksantin, ksantofil koji se nalazi kod mrkih algi, apsorbuje i plavo-zeleni dio spektra, sve do 500 nm fikobilini apsorbuju svjetlost između zelenog i crvenog dijela spektra, i to fikocijanin između 500 i 618 nm, a fikoeritrin od 450 do 576 nm ukupna apsorcija lista ili ćelije algi predstavlja zbir apsorpcija pojedinih pigmenata - raznolikost pigmenata omogućava da biljke u fotosintezi iskoriste veći opseg svjetlosnog zračenja

41
AKCIONI SPEKTRI FOTOSINTEZE
U fiziološkim procesima u kojima učestvuje svjetlost od velikog značaje je utvrđivanje akcionog spektra svjetlosti za taj proces. Akcioni spektri se mjere na sledeći način: Biljke se osvjetljavaju monohromatskom svjetlošću rastućih talasnih dužina sa 10 ili 20 nm razmaka, pri čemu se vodi računa da upadna energija svjetlosti za sve talasne dužine bude jednaka. Pri svakom osvjetljenju mjeri određuje se intenzitet fotosinteze mjerenjem nekog parametra npr. količine oslobođenog O2 – akcioni spektar se predstavlja grafički kao intenzitet procesa u funkciji talasne dužine. - PODUDARNOST AKCIONOG I APSORPCIONOG SPEKTRA U svakom fiziološkom procesu može biti aktivirana samo ona svjetlost koja je apsorbovana. Ukupni apsorpcioni spektri lista ili ćelije algi se podudaraju sa akcionim spektrom fotosinteze (izuzetak je zona u plavom dijelu spektra oko 480 nm i u crvenom dijelu spektra oko 700 nm gdje je apsorpcija veća od aktivnosti). Hl a 430 i 663 nm Hl b 450 i 643 nm
. Hl a 430 i 663 nm. Hl b 450 i 643 nm.")
42
KOMPLEMENTERNA HROMATIČNA ADAPTACIJA
Biljke se adaptiraju u odnosu na sredinu tako što izgrađuju pigmente čije je boja komplementarna boji raspoložive svjetlosti. Do mnogih mjesta nakojima biljke žive ne dopiru jednako svi zraci bijele sunčeve svjetlosti. Tako u morskoj vodi samo slojevi blizu površine imaju punu bijelu svjetlost- ti najčešće žive zelene alge koje imaju hlorofile i karotenoide i koriste plavi i crveni dio spektra. Na nešto većoj dubini ovi djelovi spektra nedostaju i tu se javljaju mrke alge kojima pigment fukoksantin omogućava da apsorbuju plavu-zelenu svjetlost koju gornji slojevi propuštaju. Na još većim dubinama, do kojih dopire uglavnom zelena, žuta i narandžasta svjetlost, žive crvene alge koje upravo ovaj dio spektra mogu da koriste zahvaljujući prisustvu fikobilina. Neke modrozelene alge su vrlo prilagodljive - ako se gaje na crvenoj svjetlosti imaju više fikoeritrina a na crvenoj imaju više fikocijanina.

Παρόμοιες παρουσιάσεις

>")
















 III℠, X 2016. Силвија Мијатовић.>")
