Κατέβασμα παρουσίασης
1
I: Σύνθεση και επεξεργασία προκαρυωτικού RNA
Η μεταγραφή I: Σύνθεση και επεξεργασία προκαρυωτικού RNA

2
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.1 Η λειτουργία της RNA πολυμεράσης είναι να μεταγράφει μια αλυσίδα δίκλωνου DNA σε RNA. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

3
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.2 Μια μεταγραφική μονάδα μεταγράφεται σε ένα ενιαίο RNA. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

4
Η μεταγραφή αποτελεί το πρώτο βήμα στη γονιδιακή έκφραση και το κυριότερο στάδιο ελέγχου της.
Πώς η RNA πολυμεράση εντοπίζει τους υποκινητές πάνω στο DNA; Πώς οι ρυθμιστικές πρωτεΐνες αλληλεπιδρούν με την RNA πολυμεράση (και μεταξύ τους) για να ενεργοποιήσουν ή να καταστείλουν συγκεκριμένα βήματα της μεταγραφής;
 για να ενεργοποιήσουν ή να καταστείλουν συγκεκριμένα βήματα της μεταγραφής;")
5
Η μεταγραφή γίνεται με ζευγάρωμα βάσεων σε μια «θηλιά» αποδιαταγμένου DNA

6
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.3 Οι αλυσίδες DNA διαχωρίζονται για να σχηματίσουν μια μεταγραφική θηλιά. Το RNA συντίθεται με το ζευγάρωμα συμπληρωματικών βάσεων με μία από τις αλυσίδες του DNA. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

7
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.5 Κατά τη μεταγραφή, η θηλιά διατηρείται μέσα στη βακτηριακή RNA πολυμεράση, η οποία αποδιατάσσει και επαναδιατάσσει το DNA συνθέτοντας RNA. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

8
Η αντίδραση της μεταγραφής συντελείται σε τέσσερα στάδια
Αναγνώριση της μήτρας Κλειστό σύμπλοκο Ανοικτό σύμπλοκο Έναρξη Ολοκληρώνεται εάν και εφόσον το ένζυμο καταφέρει να μετακινηθεί από την αρχική του θέση. Επιμήκυνση Τερματισμός

9
Η σύνδεση της RNA πολυμεράσης στον υποκινητή

10
Εικόνα 9.6 Η μεταγραφή έχει τέσσερα στάδια που ενέχουν διαφορετικές αλληλεπιδράσεις της RNA πολυμεράσης με το DNA. Το ένζυμο προσδένεται στον υποκινητή και αποδιατάσσει το DNA, παραμένει στάσιμο στην έναρξη, μετακινείται κατά μήκος του DNA στην επιμήκυνση και αποσυνδέεται στον τερματισμό.

11
Η βακτηριακή RNA pol αποτελείται από πολλές υπομονάδες

12
Σχηματική απεικόνιση του διαχωρισμού των υπομονάδων της E
Σχηματική απεικόνιση του διαχωρισμού των υπομονάδων της E. coli RNA πολυμέρασης με SDS-PAGE

13
Απομόνωση των υπομονάδων της E. coli RNA πολυμεράσης

14
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.16 Οι RNA πολυμεράσες των ευβακτηρίων αποτελούνται από τέσσερα είδη υπομονάδων: οι α, β και β΄ έχουν σχετικά σταθερά μεγέθη σε διάφορα είδη βακτηρίων, ενώ η σ ποικίλλει σε μεγαλύτερο βαθμό. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

16
Η βακτηριακή RNA pol αποτελείται από το κεντρικό ένζυμο και τον παράγοντα σ

17
Ο κύκλος του παράγοντα σ

18
Εικόνα 9.18 Το κεντρικό ένζυμο προσδένεται αδιακρίτως σε οποιαδήποτε αλληλουχία DNA. Σημαντικό ρόλο για τη χαλαρή αυτή πρόσδεση παίζει η ηλεκτρο-στατική έλξη ανάμεσα στη βασική πρωτεΐνη και το όξινο νουκλεϊκό οξύ. Το σύμπλοκο αυτό είναι σταθερό με χρόνο ημιζωής ~60 min. Ο παράγοντας σίγμα μειώνει τη συγγένεια για τις τυχαίες αλληλουχίες κατά ~104 και το χρόνο ημιζωής στο ~1 sec. Όμως, ο παράγοντας σ παρέχει εξειδίκευση για τους υποκινητές. Έτσι, το ολοένζυμο έχει σταθερά πρόσδεσης ~1000 φορές αυξημένη σε σχέση με το κεντρικό ένζυμο και με χρόνο ημιζωής αρκετών ωρών.

19
Η σύνδεση με τον παράγοντα σ μεταβάλλεται κατά την έναρξη

20
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.19 Η RNA πολυμεράση διέρχεται από αρκετά στάδια πριν τη φάση επιμήκυνσης. Ένα κλειστό δυαδικό σύμπλοκο μετατρέπεται σε μια ανοικτή μορφή και μετά σε ένα τριαδικό σύμπλοκο. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

21
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.26 Ο παράγοντας σίγμα και το κεντρικό ένζυμο ανακυκλώνονται σε διαφορετικές φάσεις κατά τη μεταγραφή. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

22
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.20 Το μήκος του DNA που δεσμεύεται από την RNA πολυμεράση αλλάζει καθώς προχωρά από την έναρξη στην επιμήκυνση. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

23
Η αναγνώριση του υποκινητή εξαρτάται από πρότυπες αλληλουχίες

24
T82T84G78A65C54A T80A95T45A60A50T96 Εικόνα 9.27 Ένας τυπικός υποκινητής αποτελείται από τρία στοιχεία: τις πρότυπες αλληλουχίες στις θέσεις -35 και -10 και το σημείο έναρξης.

25
Εικόνα 9.28 Η αλληλουχία -35 χρησιμοποιείται για την αρχική αναγνώριση και η αλληλουχία -10 για την αντίδραση τήξης, που μετατρέπει ένα κλειστό σύμπλοκο σε ανοικτό. Οι μειορυθμικές μεταλλάξεις στην –35 μειώνουν το ρυθμό σχηματισμού του κλειστού συμπλόκου. Οι μειορυθμικές μεταλλάξεις στην –10 δεν επηρεάζουν τον αρχικό σχηματισμό του συμπλόκου, όμως επιβραδύνουν τη μετατροπή του σε ανοικτό. Άρα: Η -35 παρέχει το σήμα αναγνώρισης για την RNApol Η -10 επιτρέπει τη μετατροπή από κλειστή σε ανοικτή μορφή (Τ/Α)
")
26
Η RNA pol προσδένεται στη μία επιφάνεια του DNA

27
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.29 Η μέθοδος της ιχνηλάτησης προσδιορίζει τις θέσεις πρόσδεσης πρωτεϊνών στο DNA βάσει της προστασίας που παρέχουν ενάντια στη δημιουργία εγκοπών. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

28
DNase I footprinting

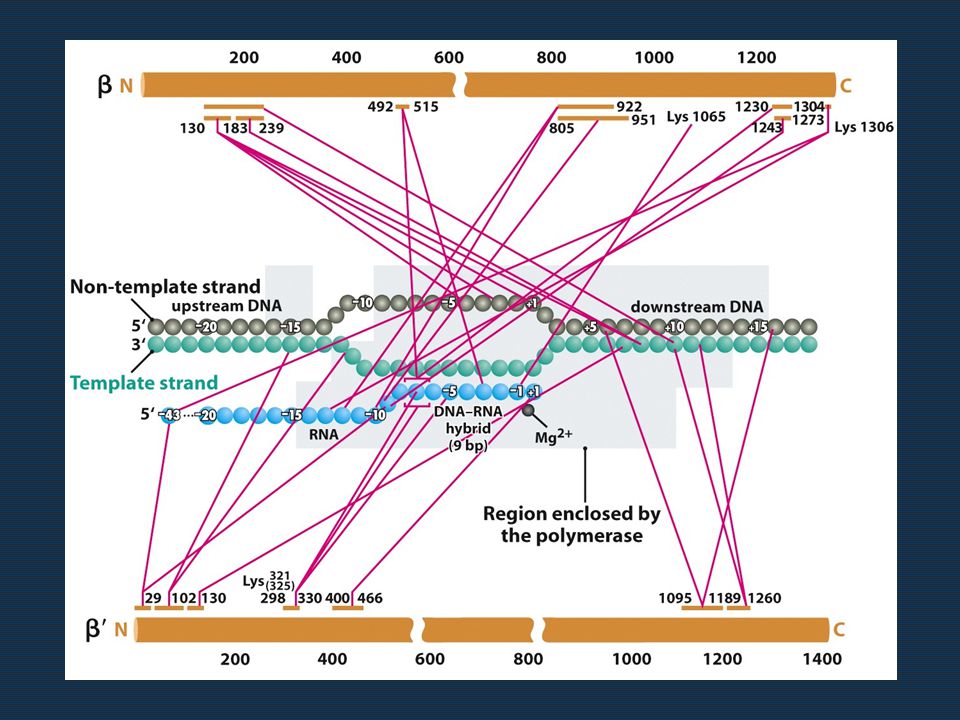
29
Εικόνα 9.30 Η μία όψη του υποκινητή περιέχει τα σημεία επαφής με την RNA πολυμεράση.

30
Είναι αξιοσημείωτο ότι σε διαφορετικούς υποκινητές οι ίδιες θέσεις παρέχουν τα σημεία επαφής, αν και μπορεί στις θέσεις αυτές να βρίσκονται διαφορετικές βάσεις. Αυτό δείχνει ότι ο μηχανισμός πρόσδεσης της RNA pol είναι κοινός, αν και η αντίδραση δεν εξαρτάται από την παρουσία συγκεκριμένων βάσεων σε ορισμένα σημεία επαφής. Για το λόγο αυτό ορισμένα σημεία επαφής δεν αποτελούν θέσεις μετάλλαξης. Επίσης, δεν αντιστοιχούν όλες οι μεταλλάξεις σε θέσεις επαφής. Δηλαδή: άλλο οι θέσεις αναγνώρισης, άλλο οι θέσεις επαφής

31
Η έναρξη της μεταγραφής μπορεί να ρυθμίζεται με αντικατάσταση παραγόντων σίγμα

32
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.32 Ο παράγοντας σίγμα συνδεδεμένος με το κεντρικό ένζυμο καθορίζει την ομάδα των υποκινητών όπου θα αρχίσει η μεταγραφή. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

33
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.33 Εκτός από τον σ70, η E. coli έχει αρκετούς παράγοντες σίγμα που επάγονται σε συγκεκριμένες περιβαλλοντικές συνθήκες. Ο εκθέτης στο όνομα του παράγοντα υποδηλώνει τη μάζα του. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

34
Στην E. coli η έκφραση 17 πρωτεϊνών θερμικού σοκ ελέγχεται από το γονίδιο rpoH που παράγει τον εναλλακτικό παράγοντα σ32 Η συσσώρευση αποδιαταγμένων πρωτεϊνών που προκύπτουν από την αύξηση θερμοκρασίας επάγει τον σ32 Ο σ70 και ο σ32 ανταγωνίζονται για τη διαθέσιμη ποσότητα κεντρικού ενζύμου Ο σ32 είναι ασταθής, γεγονός που επιτρέπει την ταχεία αύξηση ή μείωση της ποσότητάς του

35
Ο παράγοντας σΕ αποκρίνεται σε πιο ακραίες θερμοκρασιακές μεταβολές από τον σ32
Εικόνα 9.34 Η RseA συντίθεται ως μία πρωτεΐνη της εσωτερικής μεμβράνης. Η κυτταροπλασματική της επικράτεια προσδένεται στον παράγοντα σE. Η συσσώρευση αποδιαταγμένων πρωτεϊνών ενεργοποιεί στον περιπλασματικό χώρο την DegS που αποκόπτει το C-άκρο της RseA και στη συνέχεια η YaeL αποκόπτει το Ν-άκρο από το κυτταρόπλασμα. Η αποκοπή από το κυτταρόπλασμα απελευθερώνει τον σE.

36
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.35 Οι παράγοντες σίγμα της E. coli αναγνωρίζουν υποκινητές με διαφορετικές πρότυπες αλληλουχίες. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

37
Οι παράγοντες σ δημιουργούν άμεσες επαφές με το DNA
Άμεση απόδειξη προέρχεται από ορισμένες μεταλλάξεις του σ που αντισταθμίζουν κάποιες άλλες μεταλλάξεις στις περιοχές –35 και –10.

38
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.36 Χάρτης του παράγοντα σ70 της E. coli στον οποίο προσδιορίζονται οι συντηρημένες περιοχές. Οι περιοχές 2.1 και 2.2 συνδέονται με την κεντρική πολυμεράση, η 2.3 χρειάζεται για την τήξη και οι 2.4 και 4.2 συνδέονται με τις αλληλουχίες -10 και -35 του υποκινητή. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

39
Εικόνα 9. 37 Τα αμινοξέα στην α-έλικα 2
Εικόνα 9.37 Τα αμινοξέα στην α-έλικα 2.4 του σ70 δημιουργούν ειδικές επαφές με βάσεις στην περιοχή -10 της κωδικής αλυσίδας του υποκινητή

40
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.38 Το Ν-τελικό άκρο του σίγμα εμποδίζει τις επικράτειες πρόσδεσης στο DNA να προσδεθούν σε αυτό. Όταν σχηματίζεται ένα ανοικτό σύμπλοκο, το Ν-τελικό άκρο μετακινείται 20 Ǻ μακριά και οι δύο επικράτειες πρόσδεσης στο DNA απομακρύνονται η μία από την άλλη κατά 15 Ǻ Αν το Ν-τελικό άκρο του παράγοντα σ απαλειφθεί, τότε μπορεί να συνδεθεί με το DNA. Έξτρα μεταλλάξεις μπορούν να προσδιορίσουν τα ακριβή σημεία επαφής του σ με τις περιοχές -35 και -10. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

41
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.39 Ο παράγοντας σίγμα έχει μια επιμήκη δομή που εκτείνεται κατά μήκος της επιφάνειας των υπομονάδων του κεντρικού ενζύμου όταν σχηματίζεται το ολοένζυμο. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

42
Η βακτηριακή RNA pol τερματίζει τη μεταγραφή σε διακριτές θέσεις

43
Εικόνα 9.46 Οι αλληλουχίες DNA που απαιτούνται για τον τερματισμό εντοπίζονται πριν την αλληλουχία τερματισμού. Ίσως είναι αναγκαίος ο σχηματισμός μιας φουρκέτας στο RNA. Το 3’ άκρο του μορίου μπορεί να έχει δημιουργηθεί με αποκοπή του πρωτογενούς μεταγράφου και επομένως να μην αντιστοιχεί στην πραγματική θέση τερματισμού της μεταγραφής. Ο αντιτερματισμός έχει σαν αποτέλεσμα την αναγνωστική διέλευση. Άρα, ο τερματισμός μπορεί να αποτελέσει μια ευκαιρία ελέγχου γονιδιακής έκφρασης

44
Υπάρχουν δύο τύποι αλληλουχιών τερματισμού στην E. coli

45
Εικόνα 9.47 Οι ενδογενείς αλληλουχίες τερματισμού περιλαμβάνουν παλινδρομικές περιοχές οι οποίες σχηματίζουν φουρκέτες (δομές στελέχους-βρόχου) με ποικίλο μήκος, από 7-20 bp. Η δομή στελέχους-βρόχου περιλαμβάνει μια περιοχή πλούσια σε G-C και ακολουθείται από μια σειρά καταλοίπων U. Υπάρχουν ~1100 τέτοιες αλληλουχίες στο γονιδίωμα της E. coli => τα μισά γονίδια φέρουν ενδογενείς τερματιστές. Ποια είναι η επίδραση της φουρκέτας; Πιθανώς μια παύση (~60sec) που αποσταθεροποιεί ακόμα περισσότερο τα ζευγάρια Α=U. Η αποτελεσματικότητα τερματισμού in vitro είναι από 2-90%. Συνεπώς η φουρκέτα και τα U είναι αναγκαία αλλά όχι επαρκή.
 που αποσταθεροποιεί ακόμα περισσότερο τα ζευγάρια Α=U. Η αποτελεσματικότητα τερματισμού in vitro είναι από 2-90%. Συνεπώς η φουρκέτα και τα U είναι αναγκαία αλλά όχι επαρκή.")
46
Πώς λειτουργεί ο παράγοντας ρ (Rho)
Ο παράγοντας ρ είναι μια απαραίτητη πρωτεΐνη στην E. coli που λειτουργεί αποκλειστικά στο στάδιο του τερματισμού Αποτελείται από έξι όμοιες υπομονάδες και έχει μοριακό βάρος ~275 kD. Κάθε υπομονάδα έχει μια επικράτεια πρόσδεσης στο RNA και μια επικράτεια υδρόλυσης του ΑΤΡ.

47
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.48 Ένας Rho-εξαρτώμενος τερματιστής έχει αλληλουχία πλούσια σε C και φτωχή σε G, και εντοπίζεται πριν από την ακριβή θέση (ή θέσεις) τερματισμού. Η αλληλουχία παρουσιάζεται με τη μορφή του RNA. Αντιπροσωπεύει το 3΄ άκρο του RNA. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
 τερματισμού. Η αλληλουχία παρουσιάζεται με τη μορφή του RNA. Αντιπροσωπεύει το 3΄ άκρο του RNA. Genes VIII - Ακαδημαϊκές Εκδόσεις")
48
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.49 Ο παράγοντας Rho «καταδιώκει» την RNA πολυμεράση κατά μήκος του RNA και μπορεί να προκαλέσει τερματισμό όταν προλάβει το στάσιμο ένζυμο σε μια Rho-εξαρτώμενη αλληλουχία τερματισμού. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

49
Πολικότητα (Polarity): σε μερικές περιπτώσεις, μια ανερμηνεύσιμη μετάλλαξη σε ένα γονίδιο μιας μεταγραφικής μονάδας εμποδίζει τη μεταγραφή των επόμενων γονιδίων της ίδιας μονάδας. Πώς εξηγείται;;

50
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.50 Η δράση του παράγοντα Rho μπορεί να συνδέσει τη μεταγραφή με τη μετάφραση όταν η Rho-εξαρτώμενη αλληλουχία τερματισμού βρίσκεται λίγο μετά από μια ανερμηνεύσιμη μετάλλαξη.

51
Τι να διαβάσετε Genomes Genes VIII Κεφάλαιο 9: Κεφάλαιο 11.2 και 11.3
9.3 9.6 – 9.8 9.12 – 9.14 9.16 – 9.17 9.20 – 9.22 Genomes Κεφάλαιο 11.2 και 11.3 Κεφάλαιο 12.1