Κατέβασμα παρουσίασης
1
Διασύνδεση μη ειδικής-ειδικής ανοσίας Πρόσληψη, επεξεργασία, παρουσίαση αντιγόνου.
Καραγκούνης Χρήστος Ειδικευόμενος Ιατρικής Βιοπαθολογίας, Γενικό Νοσοκομείο "ΙΠΠΟΚΡΑΤΕΙΟ", Θεσσαλονίκη Συντονίστριες: Λεμονιά Σκούρα-Ασημίνα Φυλάκτου

2
Βασικοί μηχανισμοί της Φυσικής και Επίκτητης Ανοσίας

3
Η φυσική ανοσία γνωστοποιεί τη φύση του αντιγόνου στα κύτταρα της ειδικής ανοσίας και αποτελεί απαραίτητη προϋπόθεση για να ξεκινήσει η ειδική ανοσιακή απόκριση

4
Προϋπόθεση της έναρξης της ειδικής ανοσιακής απόκρισης είναι η ειδική αναγνώριση του ξένου αντιγόνου από τους υψηλής εξειδίκευσης υποδοχείς των λεμφοκυττάρων για αυτό το αντιγόνο Οι υποδοχείς των Β λεμφοκυττάρων (B cell receptor / BCR)(ανοσοσφαιρίνες προσδεδεμένες στην επιφάνειά τους) αναγνωρίζουν ένα ευρύ φάσμα μακρομορίων (πρωτεΐνες, πολυσακχαρίτες, λιπίδια και νουκλεϊκά οξέα) χωρίς την ανάγκη επεξεργασίας του αντιγόνου και παρουσίασής του από αντιγονοπαρουσιαστικά κύτταρα
(ανοσοσφαιρίνες προσδεδεμένες στην επιφάνειά τους) αναγνωρίζουν ένα ευρύ φάσμα μακρομορίων (πρωτεΐνες, πολυσακχαρίτες, λιπίδια και νουκλεϊκά οξέα) χωρίς την ανάγκη επεξεργασίας του αντιγόνου και παρουσίασής του από αντιγονοπαρουσιαστικά κύτταρα.")
5
Ο Τ κυτταρικός υποδοχέας TCR αναγνωρίζει πεπτίδια που παρουσιάζονται στην επιφάνεια των αντιγονοπαρουσιαστικών κυττάρων (Antigen Presenting Cells-APCs) συνδεδεμένα με ένα «ίδιο» HLA μόριο (MHC περιορισμός) Ο MHC περιορισμός αποδεικνύει τον κεντρικό ρόλο του HLA μορίου στην ειδική ανοσιακή απάντηση

6
HLA μόρια τάξης Ι (HLA –A, -B, -C)
(HLA –DR, -DP, -DQ)
")
7
Τάξης Ι μόρια συνδέουν στη σχισμοειδή τους θήκη (peptide binding group) πεπτίδια 8-10 αμινοξέων
Το μήκος των πεπτιδίων που συνδέονται με τα τάξης ΙΙ μόρια δεν περιορίζεται (13-20 αμινοξέα) Η ειδικότητα ως προς την ακολουθία του πεπτιδίου παρέχεται από ένα μικρό αριθμό εσοχών (pockets) στο δάπεδο της θήκης που δεσμεύουν αμινοξέα του πεπτιδίου με συμπληρωματικό τρόπο
 Η ειδικότητα ως προς την ακολουθία του πεπτιδίου παρέχεται από ένα μικρό αριθμό εσοχών (pockets) στο δάπεδο της θήκης που δεσμεύουν αμινοξέα του πεπτιδίου με συμπληρωματικό τρόπο.")
8
Υπάρχουν συνήθως 2 ή 3 κύριες θέσεις πρόσδεσης χαρακτηριστικές και σταθερές για κάθε HLA μόριο (binding motif) Δυνατή η σύνδεση όλων των πεπτιδίων που φέρουν τα ίδια συνδετικά αμινοξέα στις κύριες θέσεις πρόσδεσης

9
Τα MHC τάξης Ι εκφράζονται σε όλα τα εμπύρηνα κύτταρα

10
Τα HLA μόρια παρουσιάζουν τεράστιο πολυμορφισμό και ποικιλία

11
Αντιγονοπαρουσιαστικά κύτταρα
Τα εξειδικευμένα κύτταρα τα οποία παραλαμβάνουν, επεξεργάζονται και παρουσιάζουν το ξένο αντιγόνο στα Τ λεμφοκύτταρα είναι τα αντιγονοπαρουσιαστικά κύτταρα (APCs) εκφράζουν MHC μόρια απαραίτητα για την ενεργοποίηση των παρθένων Τ λεμφοκυττάρων παρέχουν επιπλέον σήματα ώστε να ενεργοποιηθούν τα παρθένα Τ λεμφοκύτταρα είναι κυρίως τα δενδριτικά κύτταρα (DCs), τα μακροφάγα (ΜΦ) και τα Β κύτταρα που χαρακτηρίζονται ως «επαγγελματικά» professional APCs τα κύτταρα που εκφράζουν ΜHC τάξης Ι μόρια λειτουργούν ως κύτταρα στόχοι
 εκφράζουν MHC μόρια. απαραίτητα για την ενεργοποίηση των παρθένων Τ λεμφοκυττάρων. παρέχουν επιπλέον σήματα ώστε να ενεργοποιηθούν τα παρθένα Τ λεμφοκύτταρα. είναι κυρίως τα δενδριτικά κύτταρα (DCs), τα μακροφάγα (ΜΦ) και τα Β κύτταρα που χαρακτηρίζονται ως «επαγγελματικά» professional APCs. τα κύτταρα που εκφράζουν ΜHC τάξης Ι μόρια λειτουργούν ως κύτταρα στόχοι.")
12
Τα DCs είναι τα πιο αποτελεσματικά APCs στην πρωτογενή Τ ανοσιακή απάντηση και προσλαμβάνουν το αντιγόνο με διάφορους μηχανισμούς Τα μικροβιακά αντιγόνα προκαλούν ανοσιακές αντιδράσεις, μη ειδικής ανοσίας διαμέσου της σύνδεσής τους με TLRs Αποτέλεσμα παραγωγή των φλεγμονωδών κυτταροκινών (IL-1, TNF-α), ωρίμανση των DCs (έκφραση MHC και συνδιεγερτικών μορίων) και μετανάστευση στις Τ κυτταρικές περιοχές των λεμφαδένων, όπου μεταφέρουν το αντιγόνο
, ωρίμανση των DCs (έκφραση MHC και συνδιεγερτικών μορίων) και μετανάστευση στις Τ κυτταρικές περιοχές των λεμφαδένων, όπου μεταφέρουν το αντιγόνο.")
13
Τα πεπτίδια που συνδέονται με τη σχισμοειδή θήκη του MHC, είναι αποτέλεσμα μιας σειράς μεταβολικών φαινομένων, που οδηγούν στη διάσπαση των συνήθως ευμεγεθών και πολύπλοκων αντιγονικών μορίων, διαδικασία γνωστή ως επεξεργασία του αντιγόνου

14
Ενδογενής οδός διαμέσου των HLA τάξης Ι μορίων για πεπτίδια που προέρχονται από ενδοκυττάριες πρωτεΐνες (ιικές, νεοπλασματικές) του κυτταροπλάσματος και παρουσιάζονται στα CD8+ T λεμφοκύτταρα (CTL) Εξωγενής οδός διαμέσου των HLA τάξης IΙ μορίων για πεπτίδια εξωκυττάριων αντιγόνων σε ενδοκυτταροπλασματικά κυστίδια και παρουσιάζονται στα CD4+ T λεμφοκύτταρα

15
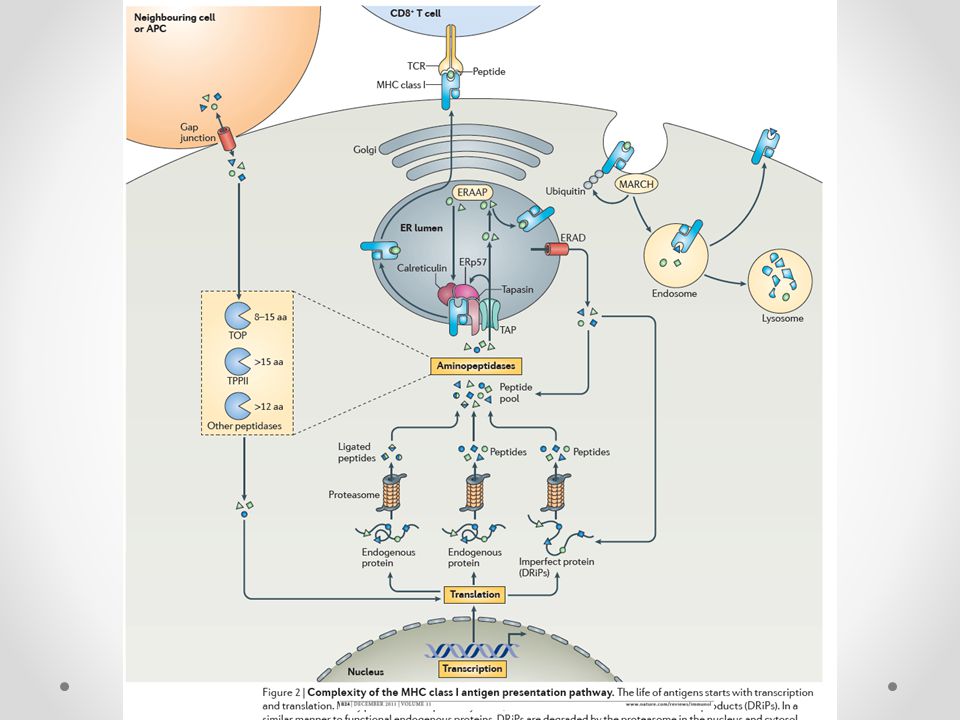
Ενδογενής οδός επεξεργασίας
Η «ενδογενής οδός» επεξεργασίας μπορεί να συνοψιστεί στα ακόλουθα βήματα: 1) διάσπαση των πρωτεϊνών από το πρωτεάσωμα και τις αμινοπεπτιδάσες 2) μεταφορά στο ενδοπλασματικό δίκτυο μέσω των μεταφορέων TAP1, TAP2 3) σύνδεση με τα MHC Ι μόρια στο ΕΔ με τη βοήθεια των chaperones (συνοδευτικών μορίων) 4) το σύμπλεγμα μεταφέρεται μέσω του συστήματος Golgi στην επιφάνεια του κυττάρου προς παρουσίαση στα CD8+ T λεμφοκύτταρα
 διάσπαση των πρωτεϊνών από το πρωτεάσωμα και τις αμινοπεπτιδάσες. 2) μεταφορά στο ενδοπλασματικό δίκτυο μέσω των μεταφορέων TAP1, TAP2. 3) σύνδεση με τα MHC Ι μόρια στο ΕΔ με τη βοήθεια των chaperones (συνοδευτικών μορίων) 4) το σύμπλεγμα μεταφέρεται μέσω του συστήματος Golgi στην επιφάνεια του κυττάρου προς παρουσίαση στα CD8+ T λεμφοκύτταρα.")
16
1) Η διάσπαση των πρωτεϊνών στο κυτταρόπλασμα
Κατ’ αρχήν σύνδεσή τους με μια μικρού ΜΒ πρωτεΐνη την ουμπικουιτίνη Είσοδος στο πρωτεάσωμα (πολυκαταλυτικό πρωτεολυτικό σύστημα- 20S πυρήνας με δύο 19S καλύμματα) Σημείο κλειδί στην ενδογενή οδό επεξεργασίας Ανοσπρωτεάσωμα (η IFN-γ μεταβάλλει τη δράση του πρωτεασώματος με την ενσωμάτωση στον πυρήνα του τριών β υπομονάδων (LMP2, LMP7, MECL1) αυξημένη διάσπαση πεπτιδίων μετά τα υδροφοβικά αμινοξέα και παραγωγή πεπτιδίων με καρβοξυτελικά που προτιμούνται από τα MHC I
 Σημείο κλειδί στην ενδογενή οδό επεξεργασίας. Ανοσπρωτεάσωμα (η IFN-γ μεταβάλλει τη δράση του πρωτεασώματος με την ενσωμάτωση στον πυρήνα του τριών β υπομονάδων (LMP2, LMP7, MECL1) αυξημένη διάσπαση πεπτιδίων μετά τα υδροφοβικά αμινοξέα και παραγωγή πεπτιδίων με καρβοξυτελικά που προτιμούνται από τα MHC I.")
17
Διαμέσου της ιντερφερόνης-γ ενεργοποίηση του πρωτεασώματος με το σύμπλεγμα PA28
Αύξηση του ρυθμού παραγωγής πεπτιδίων Αποφυγή της επιπλέον επεξεργασίας σε βάρος της αντιγονικότητας

18
Τα ιικά αντιγόνα παρουσιάζονται στα HLA τάξης Ι πολύ ταχύτερα από το φυσιολογικό χρόνο ημίσειας ζωής τους Επιτυγχάνεται μέσω των DRiPs (defective ribosomal products) (30-70% των παραγόμενων πρωτεϊνών) Το πρωτεάσωμα παράγει πεπτίδια τα οποία είναι έτοιμα για το ενδοπλασματικό δίκτυο (ΕΔ) Κυτταρικές πρωτεΐνες (chaperons) όπως το σύμπλεγμα TRiC (TCP-1 ring complex) προστατεύει αυτά τα πεπτίδια από περαιτέρω διάσπαση Ανάγκη δράσης της αμινοπεπτιδάσης ERAAP (aminopeptidase associated with antigen processing) στο ΕΔ
 (30-70% των παραγόμενων πρωτεϊνών) Το πρωτεάσωμα παράγει πεπτίδια τα οποία είναι έτοιμα για το ενδοπλασματικό δίκτυο (ΕΔ) Κυτταρικές πρωτεΐνες (chaperons) όπως το σύμπλεγμα TRiC (TCP-1 ring complex) προστατεύει αυτά τα πεπτίδια από περαιτέρω διάσπαση. Ανάγκη δράσης της αμινοπεπτιδάσης ERAAP (aminopeptidase associated with antigen processing) στο ΕΔ.")
19
2) Μεταφορά στο ενδοπλασματικό δίκτυο
Τα πεπτίδια για να ενωθούν με τα τάξης Ι μόρια μεταφέρονται ενεργητικά από το κυτταρόπλασμα στο ΕΔ διαμέσου των TAP μορίων (transporters associated with antigen processing, TAP1 και TAP2) ATP εξερτώμενοι μεταφορείς πεπτιδίων Παρουσιάζουν ειδικότητα στο είδος των πεπτιδίων που μεταφέρουν Μεταλλάξεις στις πρωτεΐνες TAP εμποδίζουν την παρουσίαση του αντιγονικού πεπτιδίου από τα τάξης Ι μόρια Κωδικοποιούνται από γονίδια που βρίσκονται στο MHC και επάγεται η έκφρασή τους από ιντερφερόνες
 ATP εξερτώμενοι μεταφορείς πεπτιδίων. Παρουσιάζουν ειδικότητα στο είδος των πεπτιδίων που μεταφέρουν. Μεταλλάξεις στις πρωτεΐνες TAP εμποδίζουν την παρουσίαση του αντιγονικού πεπτιδίου από τα τάξης Ι μόρια. Κωδικοποιούνται από γονίδια που βρίσκονται στο MHC και επάγεται η έκφρασή τους από ιντερφερόνες.")
20
3) Σύνδεση με τα MHC μόρια στο ΕΔ
Στο ΕΔ σχηματίζονται τα ετεροδιμερή από μια βαριά α-αλυσίδα και τη β2-μικροσφαιρίνη, MHC τάξης Ι μόρια τα οποία είναι συνδεδεμένα με καλνεξίνη που πιθανόν βοηθάει στην κατάλληλη διαμόρφωση της βαριάς αλυσίδας, ώστε να συνδεθεί με τη β2-m

21
Μετά τη σύνδεση της α αλυσίδας με την β2-m, η καλνεξίνη αποδεσμεύεται και το MHC τάξης Ι μόριο συνδέεται με τα τσαπερόνια (chaperons) σταθεροποίησης από το ΕΔ, όπως η καλρετικουλίνη και το ERp57 Το σύμπλεγμα αντιδρά με τις ΤΑΡ πρωτεΐνες μέσω της ταπασίνης, η οποία έχοντας θέσεις δέσμευσης για τις ΤΑΡ1/ΤΑΡ2 λειτουργεί ως γέφυρα μεταξύ του συμπλέγματος MHC I μορίου/καλρετικουλίνης και ΤΑΡ1/ΤΑΡ2 πρωτεϊνών Η ταπασίνη φαίνεται να διατηρεί σταθερή τη διαμόρφωση των κενών MHC τάξης Ι μορίων (λειτουργία ανάλογη του HLA-DM μορίου για τα MHC τάξης ΙΙ μόρια) PLC (peptide loading complex): MHC I, καλρετικουλίνη, ταπασίνη, ERP57, TAP
 PLC (peptide loading complex): MHC I, καλρετικουλίνη, ταπασίνη, ERP57, TAP.")
22
Τα πεπτίδια που δεν ενώνονται με το MHC τάξης Ι μόριο όπως και τα MHC μόρια μεταφέρονται πίσω στο κυτταρόπλασμα με το ERAD (E-R associated protein degraded system) Peptide editing: διαδικασία ανταλλαγής πεπτιδίων Αν ένα τάξης Ι μόριο λοιπόν βρει ένα πεπτίδιο κατάλληλου μεγέθους και αλληλουχίας συνδέεται σταθερά μαζί του και αποδεσμεύεται από το PLC

23
4) το σύμπλεγμα μεταφέρεται μέσω του συστήματος Golgi στην επιφάνεια του κυττάρου προς παρουσίαση στα CD8+ T λεμφοκύτταρα
 το σύμπλεγμα μεταφέρεται μέσω του συστήματος Golgi στην επιφάνεια του κυττάρου προς παρουσίαση στα CD8+ T λεμφοκύτταρα")
25
Πολλοί ιοί έχουν αναπτύξει στρατηγικές διαφυγής που μπλοκάρουν το δρόμο παρουσίασης του αντιγόνου με τα MHC τάξης Ι μόρια γρήγορη απομάκρυνση των πρόσφατα σχηματισμένων μορίων MHC Ι από το ΕΔ παρεμπόδιση της μεταγραφής των γονιδίων του MHC παρεμπόδιση της μεταφοράς των πεπτιδίων στο ΕΔ με τα ΤΑΡ μόρια

26
Το PLC αποτελεί στόχο των στρατηγικών διαφυγής των ιών

27
Εξωγενής οδός Η πρόσληψη των εξωγενών μικροβίων ή μικροβιακών πρωτεϊνών από τα APCs γίνεται με διάφορους μηχανισμούς Μέσω υποδοχέων: ο αντιγονοειδικός υποδοχέας των Β λεμφοκυττάρων (BCR), Fc υποδοχείς (FCRs), υποδοχείς για το συμπλήρωμα, υποδοχείς μαννόζης, υποδοχείς Toll (TLRs) κ.ά. Φαγοκυττάρωση Πινοκύττωση
, Fc υποδοχείς (FCRs), υποδοχείς για το συμπλήρωμα, υποδοχείς μαννόζης, υποδοχείς Toll (TLRs) κ.ά. Φαγοκυττάρωση. Πινοκύττωση.")
28
Μετά την πρόσληψη του αντιγόνου από το APC, αυτό εισέρχεται σε ένα σύστημα ενδοκυττάριων κυστιδίων που καλούνται ανάλογα ενδοσώματα ή φαγοσώματα, τα οποία συγχωνεύονται με τα λυσοσώματα δημιουργώντυας τα φαγολυσοσώματα ή δευτερογενή λυσοσώματα

29
το αντιγόνο αποδομείται και διασπάται σταδιακά εκτειθέμενο σε περιβάλλον μειούμενου pH και συνεχώς αυξανόμενης ενζυμικής δραστηριότητας διαφόρων πρωτεασών (ενδοπεπτιδάσες, εξωπεπτιδάσες, λυσοσωμική θειολρεδουκτάση/ γ-interferon induced lysosomal thiol reducase/ GILT) Η ενζυμική δραστηριότητα των πρωτεασών αυξάνεται από το όξινο pH των κυστιδίων, την IFN-γ, τον TNF-α και την IL-1β, ενώ ελαττώνεται από την IL-10, τις τοξίνες και τους πεπτιδικούς αναστολείς των μικροβίων, όπως και από ενδογενείς πεπτιδικούς αναστολείς

30
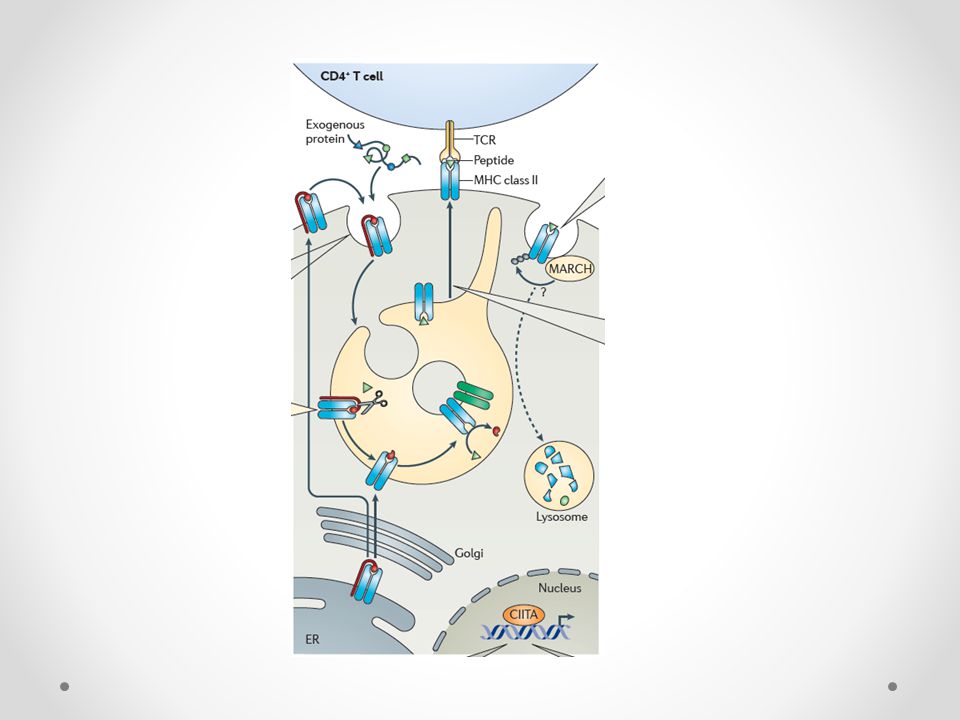
Τα APCs συνθέτουν συνεχώς ΜHC μόρια τάξης ΙΙ στο ενδοπλασματικό δίκτυο (ΕΔ)
Ο κύριος ρυθμιστής της έκφρασης των τάξης ΙΙ μορίων είναι το μόριο CIITA (MCH class II transactivator)
")
31
Κάθε μόριο τάξης ΙΙ φέρει συνδεδεμένη στη σχισμοειδή του θήκη μια μη πολυμορφική αλυσίδα την invariant chain (Ii), που αποτελεί μια τύπου ΙΙ διαμεμβρανική πρωτεΐνη κι εμποδίζει τη σύνδεση του MHC τάξης ΙΙ μορίου με ενδογενή πεπτίδια του ΕΔ

32
Το σύμπλεγμα MHC-Ii μεταφέρεται στα διαμερίσματα των ενδοσωμάτων/λυσοσωμάτων προκειμένου να συναντήσει τα πεπτίδια Διάσπαση της Ii Τα διασπασμένα ενδοκυτταρωμένα πεπτίδια στα ενδοσώματα δεν συνδέονται με τα MHC μόρια λόγω της CLIP Απομάκρυνση της CLIP με τη βοήθεια του μορίου HLA-DM

33
MIIC (MHC II compartment) το όψιμο ενδόσωμα όπου γίνεται η σύνδεση του MHC με το πεπτίδιο
Απαιτείται η ύπαρξη σε αυτό των μορίων MHC II, HLA-DM, καθεψινών (S, L) και τετρασπανινών Είναι σύστημα πολυμορφικό
 και τετρασπανινών. Είναι σύστημα πολυμορφικό.")
34
Η μεταφορά του συμπλέγματος MHC II- πεπτιδίου από το MIIC ρυθμίζεται από παράγοντες όπως η χοληστερόλη, το κυτταροπλασματικό pH, κινάσες και GTPάσες Η ημίσεια ζωή του MHC II μορίου αυξάνεται καθώς το δενδριτικό κύτταρο ωριμάζει

36
Αυτοφαγία Σε ορισμένες περιπτώσεις, MHC τάξης ΙΙ μόρια μπορεί να δεσμεύουν πεπτίδια προερχόμενα από την αποδόμηση πρωτεϊνών ενδογενούς προέλευσης διαμέσου της διαδικασίας της αυτοφαγίας Η αυτοφαγία αποτελεί διαδικασία αυτοπεψίας και προάγει τη διανομή ενδοκυττάριων οργανυλίων και κυτταροπλασματικών πρωτεϊνών από το κυτταρόπλασμα στα λυσοσσωμάτια για αποδόμηση και ανακύκλωση διατηρώντας την ομοιοστασία

37
Αποτελεί βασικό μηχανισμό σε όλα τα εμπύρηνα κύτταρα και μειώνει το οξειδωτικό stress στοχεύοντας στα κατεστραμμένα μιτοχόνδρια επιτείνεται σε κατάσταση stress όπως η λοιμοκτονία κατά την οποία καταβολίζονται ενδοκυττάριες πρωτεΐνες για την παραγωγή ενέργειας

38
Cross Presentation (Διασταυρούμενη Παρουσίαση)
Η διαδικασία της ενδογενούς επεξεργασίας και παρουσίασης του αντιγονικού πεπτιδίου από το MHC τάξης Ι στα CD8 T λεμφοκύτταρα διασφαλίζει τη δυνατότητα ανίχνευσης των μολυσμένων από τα παθογόνα κύτταρα και την καταστροφή τους από τα κυτταροτοξικά Τ λεμφοκύτταρα. Αλλά, πώς αρχικά ενεργοποιείται ένα παρθένο CD8 T λεμφοκύτταρο ειδικό για ένα πεπτίδιο; απαραίτητη η ύπαρξη ενός δενδριτικού APC κυττάρου που να εκφράζει το ίδιο σύμπλεγμα MHC Ι-πεπτιδίου με το μολυσμένο από τον ιό κύτταρο Κάποιοι ιοί όπως ο HIV δεν μολύνουν δενδριτικά κύτταρα

39
Με τη διασταυρούμενη παρουσίαση-ενεργοποίηση (cross presentation-priming) κάποιες ομάδες δενδριτικών (CD24, CD89, CD103) κυττάρων μπορούν να σχηματίσουν συμπλέγματα πεπτιδίου- MHC τάξης Ι μορίου από πεπτίδια που δεν σχηματίστηκαν στο δικό τους κυτταρόπλασμα Πρόκειται για ένα σύνδεσμο μεταξύ των δύο μονοπατιών Φαγοκυτταρωμένα νεκρά κύτταρα που μολύνθηκαν από κυτταροπλασματικά παθογόνα μπορούν να παρουσιαστούν με τα MHC τάξης Ι μόρια του δενδριτικού κυττάρου Ειδικός τρόπος παρουσίασης? (insoulin regulated aminopeptidase στα ενδοσώματα) Τρόπος ενδοκύττωσης (Fc receptors, c-type lectin)
 Τρόπος ενδοκύττωσης (Fc receptors, c-type lectin)")
40
TNF-related-apoptosis inducing ligand
Programmed cell death ligand TNF-related-apoptosis inducing ligand Μηχανισμοί διασταυρούμενης παρουσίασης και ενεργοποίησης από τα δενδριτικά κύτταρα

41
Παρουσίαση του αντιγόνου
Τα MHC τάξης ΙΙ μόρια συνδεδεμένα με πεπτίδια εξωγενών αντιγόνων διεγείρουν τα CD4+ βοηθητικά Τ λεμφοκύτταρα, να ενεργοποιήσουν τα Β λεμφοκύτταρα και τα ΜΦ, είτε με απευθείας επαφή είτε μέσω κυτταροκινών, ώστε να παράγουν ειδικά αντισώματα ή να καταστρέψουν το φαγοκυτταρωμένο αντιγόνο αντίστοιχα Τα MHC τάξης Ι μόρια που εκφράζονται σε όλα τα εμπύρηνα κύτταρα συνδεδεμένα με πεπτίδια ενδογενών αντιγόνων διεγείρουν τα CD8+ κυτταροτοξικά Τ λεμφοκύτταρα να αναγνωρίσουν και να καταστρέψουν τα κύτταρα στόχους

42
Η σταθερότητα του συμπλέγματος MHC μορίου–πεπτιδίου, στην επιφάνεια του κυττάρου είναι απαραίτητη, προκειμένου να υπάρξει επαρκής σηματοδότηση της ύπαρξης μολυσμένου κυττάρου Αν το σύμπλεγμα διασπούνταν γρήγορα θα μπορούσε το παθογόνο να διαφύγει της αναγνώρισης από τον Τ κυτταρικό υποδοχέα MHC μόρια στην επιφάνεια μη μολυσμένων κυττάρων θα μπορούσαν να συνδεθούν με πεπτίδια που απελευθερώνονταν από συμπλέγματα με MHC μόρια μολυσμένων κυττάρων και να προκαλείται κυτταροτοξική αντίδραση έναντι υγειών κυττάρων

43
Τα μόρια προσκόλλησης διαμεσολαβούν αρχικά για την αλληλεπίδραση του αντιγονοπαρουσιαστικού κυττάρου με τα παρθένα Τ λεμφοκύτταρα

44
Αυτή η παροδική αλληλεπίδραση σταθεροποιείται από την ειδική αναγνώριση του αντιγόνου

45
Τα αντιγονοπαρουσιαστικά κύτταρα μέσω της παροχής 3 ειδών σηματοδότησης προκαλούν την πολλαπλασιασμό και τη διαφοροποίσηση των παρθένων Τ λεμφοκυττάρων

46
Η απουσία του 2ου σήματος έκφρασης συνδιεγερτικών μορίων οδηγεί σε απενεργοποίηση των Τ κυττάρων

47
Συμπερασματικά...... Η βιολογία της παρουσίασης των αντιγόνων διαμέσου των μορίων MHC I και ΙΙ έχει μελετηθεί εκτενώς λόγω του κεντρικού ρόλου της στον έλεγχο της ανοσιακής απάντησης και κατά συνέπεια στη λοίμωξη, στην αυτοανοσία, στον εμβολιασμό και στη μεταμόσχευση Εντούτοις πολλές πτυχές της όλης διαδιακασίας παραμένουν αδιευκρίνιστες και αποτελούν σημεία αντιπαράθεσης Πόσο μπορεί προβλεφθεί το είδος του ανοσογονικά επικρατούντος πεπτιδίου στη σχισμοειδή θήκη του MHC? Γιατί κάποια MHC αλλήλια σχετίζονται με αυτοάνοσα νοσήματα?

48
Ποιοι κανόνες διέπουν τη δημιουργία και επιλογή των πεπτιδίων από τις κυτταροπλασματικές πεπτιδάσες και το PLC? Ποιος ο ρόλος των DRiPs? Ποιος ο ακριβής μηχανισμός των HLA-DM ή του PLC στην επιλογή του κατάλληλου πεπτιδίου? Πως τα παθογόνα και οι TLRs ελέγχουν τα MHC II? Ποιος ο μηχανισμός παρουσίασης κυτταροπλασματικών αντιγόνων μέσω του MCH II? Ποιος ο μηχανισμός παρουσίασης εξωκυυτάριων παθογόνων με MHC I

49
Ωστόσο ελπίζουμε ότι η βαθύτερη κατανόηση του συστήματος που προσφέρει η εφαρμογή των νέων τεχνολογιών θα δώσει απαντήσεις στα επόμενα χρόνια, που θα μεταφραστούν σε νέες θεραπευτικές επιλογές Ευχαριστώ






, είναι δύο μέλη της οικογένειας του ιού του έρπητα, herpesviridae, που μολύνει.>")














