Κατέβασμα παρουσίασης
2
Enzime Ce sunt enzimele? Nomenclatura enzimelor
Caracteristici generale ale enzimelor Clasificarea enzimelor Structura enzimelor Specificitatea enzimelor Mecanismul catalizei enzimatice Cinetica enzimatica Cofactori (coenzime, grupari prostetice).
.")
3
Enzime Definiţie Biocatalizatori proteici care măresc viteza reacţiilor chimice, dar numai pentru reacţiile termodinamic posibile Coordonează toate procesele metabolice Catalizează un număr redus de reacţii, foarte frecvent numai una; se caracterizează printr-o înaltă specificitate Sunt implicate în majoritatea reacţiilor biochimice din celule, fiind importante în diagnostic şi tratament

4
ai

5
Enzime Caracteristici
Au o ToC si pH optim de functionare Nu sunt produse sau consumate in timpul unei reactii chimice Nu declanseaza o reactie chimica ci intensifica viteza reactiei chimice Enzimele modifica viteza reactiei dar nu constanta de echilibru a reactiei catalizate.

6
Enzime Nomenclatura 2 denumiri: A. Denumirea recomandată sau comună
1. “- ază “ la numele substratului reacţiei Ex.: glucozidază, urează sau 2. la formula care descrie acţiunea desfăşurată Ex.: lactat dehidrogenază, adenilat ciclază Există enzime care îşi păstrează în continuare formula originală, istorică, neavând legătură cu reacţia catalizată

7
Enzime B. Denumirea ştiinţifică sau sistematică
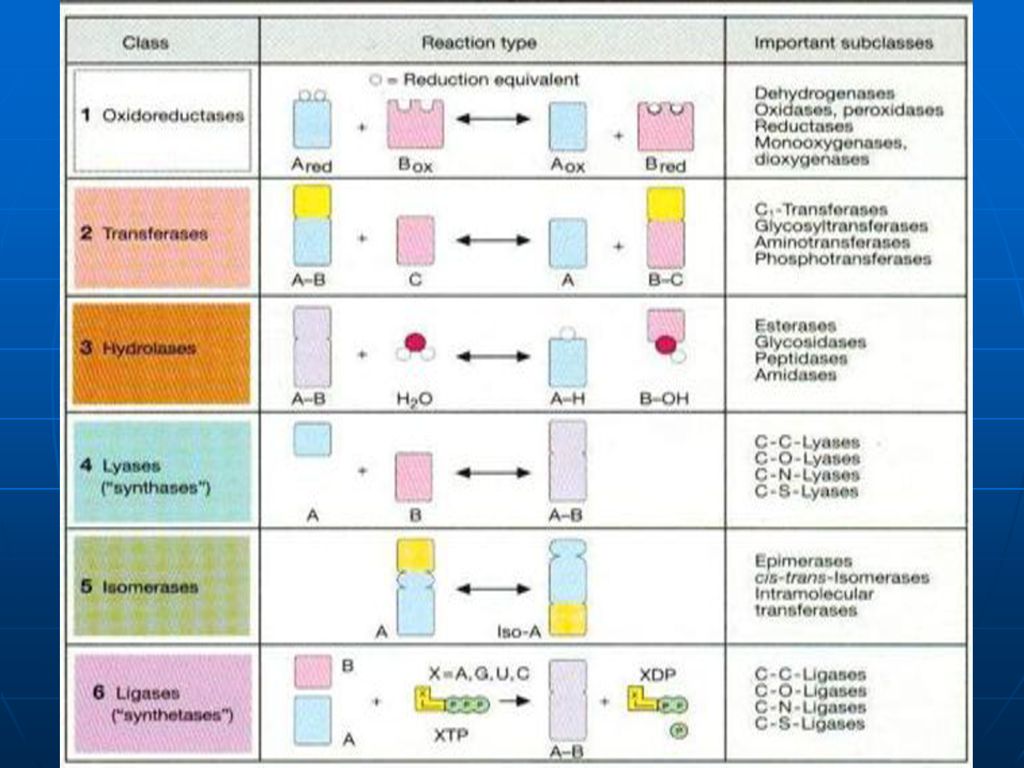
IUBMB (Uniunea Internaţională de Biochimie şi Biologie Moleculară): 6 clase de enzime, fiecare cu mai multe subclase Sufixul “- ază “ este ataşat unei formule descriptive complete a reacţiei chimice catalizate, incluzând denumirea tuturor substraturilor Ex. D-gliceraldehid 3 fosfat: NAD+ oxidoreductaza Denumire prea complexa pt a fi utilizata in practica uzuala
: 6 clase de enzime, fiecare cu mai multe subclase. Sufixul - ază este ataşat unei formule descriptive complete a reacţiei chimice catalizate, incluzând denumirea tuturor substraturilor. Ex. D-gliceraldehid 3 fosfat: NAD+ oxidoreductaza. Denumire prea complexa pt a fi utilizata in practica uzuala.")
9
Enzime- clasificare 1) Oxidoreductazele
- enzime ce catalizează reacţiile de oxidoreducere ce au loc în organismele vii; Lactat dehidrogenaza Piruvat Lactat

10
Enzime- clasificare 2) Transferazele
- enzime ce catalizează reacţii de transfer ale unor grupe de atomi (C-, N-, P-) ce pot fi şi grupări funcţionale de la un substrat la altul, fără ca aceste grupări să existe libere în timpul procesului; Metilen THF Serin-hidroximetil transferaza THF
 ce pot fi şi grupări funcţionale de la un substrat la altul, fără ca aceste grupări să existe libere în timpul procesului; Metilen. THF. Serin-hidroximetil. transferaza. THF.")
11
Enzime- clasificare 3) Hidrolazele
- enzime ce catalizează scindarea diferitelor legături covalente din molecula substratului cu participarea moleculelor de apă; + HOH ---- CO2 + NH3 ureaza UREE

12
Enzime- clasificare 4) Liazele
- catalizează clivarea legăturilor de tip C-C, C-S, C-N cu rearanjarea ulterioară a valenţelor; Piruvat decarboxilaza

13
Enzime- clasificare 5)Izomerazele
- enzime care catalizează diferite reacţii de izomerizare a substratelor;

14
Enzime- clasificare 6)Ligazele sau sintetazele
- enzime ce catalizează formarea unor noi legături chimice C – C sau C– O, S, N în majoritatea cazurilor utilizându-se energia stocată în legăturile macroergice ale moleculelor de ATP. Piruvat carboxilaza

15
Enzime

16
Structura enzimelor 2 categorii:
1. formate numai din AA; structură exclusiv proteică. Ex proteaze, lipaze, ribonucleaze 2. componenta proteică sau APOENZIMA + componenta neproteică sau COFACTOR; majoritatea enzimelor COFACTORI: A. Gruparea prostetică Componenta neproteică legată de apoE prin legături ferme, stabile, neputându-se desprinde de aceasta. Ex FMN, FAD

17
Structura enzimelor

18
Structura enzimelor COFACTORI: B. Coenzima
Derivat al unei vitamine, componenta neproteică legată de apoE prin legături labile Poate trece uşor de la o apoE la alta NAD+; NADH, H+; NADP+

19
Structura enzimelor

20
Structura enzimelor COFACTORI: B. Ionii metalici
Sunt necesari pentru pentru activitatea a numeroase enzime. După modul de legare a şi rolul ionului metalic, enzimele sunt: - metaloenzime ce conţin o cantitate bine definită de ion metalic funcţional, fiind strâns legat de apoE - enzime metaloactive, a căror activitate creşte în prezenţa ionului metalic, care leagă metalul slab, iar cantitatea acestuia nu este strict delimitată

21
Structura enzimelor COFACTORI: Metaloenzime:
- Fe-enzime: grup prostetic hem (citocromi, catalaze, peroxidaze); - Cu-enzime: citocromoxidaza, superoxid-dismutaza, tiroxin-hidroxilaza; - Mn-enzime: peptidaza, arginaza, izocitrat-dehidrogenaza; - Co-enzime: cobalamin-enzime; - Se-enzime: glutationperoxidaza
; - Cu-enzime: citocromoxidaza, superoxid-dismutaza, tiroxin-hidroxilaza; - Mn-enzime: peptidaza, arginaza, izocitrat-dehidrogenaza; - Co-enzime: cobalamin-enzime; - Se-enzime: glutationperoxidaza.")
22
Structura enzimelor

23
Structura enzimelor Unele enzime necesită pentru a fi active prezenţa coenzimelor, altele a gruparilor prostetice sau a ionilor metalici In reacţiile catalizate de enzime ce necesită prezenţa coenzimelor, acestea sunt considerate al doilea substrat, fiind numite cosubstrat Modificările pe care le suferă coenzima contrabalansează exact modificările de la nivelul substratului

24
Cofactori enzimatici

25
Structura enzimelor Regiunea din enzimă unde se fixează moleculele substratului şi se produc reacţiile chimice care transformă aceste molecule se numeşte centru activ.

26
Centrul activ al enzimelor
O regiune a componentei proteice în care resturile de AA sunt aranjate astfel încât să permită o interacţiune adecvată cu substratul, numit la modul generic ligand

27
Centrul activ al enzimelor
Complexul ES de la nivelul centrului activ este stabilizat prin: Atracţii electrostatice Legături de hidrogen Interacţiuni hidrofobe După fixarea S la centrul activ al E numeroase resturi laterale ale AA din vecinătatea ligandului catalizează transformarea acestuia.

28
Centrul activ al enzimelor
Centrul activ cuprinde resturi de fixare a S cât şi resturi catalitice Centrul activ ocupă o porţiune mică din enzimă şi este o entitate tridimensională Cofactorii joacă un rol important atât în fixarea S cât şi în procesul catalitic S trebuie să aibă o formă potrivită pentru a pătrunde în centrul activ al unei E

29
Centrul activ al enzimelor
1890 Fisher propune modelul “broască- cheie”: S trebuie să se potrivească perfect la centrul activ pentru a suferi procesul catalitic Fisher

30
Centrul activ al enzimelor
1958 Koshland susţine că centru activ al enzimei nu este rigid Forma acestuia se modifică în momentul legării S= recunoastere dinamica a S de catre E

31
Centrul activ al enzimelor
S se leagă de E prin forţe relativ slabe; complexul ES care se formează trece în produşii de reacţie cu eliberarea E sau poate reface S iniţial

32
Centrul activ al enzimelor
Apare sub forma unei despicături în apoE căptuşită cu AA hidrofobi , apa fiind exclusă în mare parte Conţine şi aa polari esenţiali în procesul de cataliză Pe lângă resturile catalitice aflate în centru activ, care participă direct la transformarea S, apoE conţine şi aşa-numitele resturi de specificitate, unde sunt prezenti aa cu rol în recunoaşterea S Ex. ribonucleaza scindeaza ARN Centrul activ prezinta: resturi catalitice (His 12 si His 119) si resturi de specificitate: intre aa exista 5 aa bazici care recunosc ARN cu caracter acid a
 si resturi de specificitate: intre aa exista 5 aa bazici care recunosc ARN cu caracter acid. a.")
33
Centrul activ al enzimelor
Contine: Ser, Cys, His, Tyr, Lys Enzimele alosterice contin pe langa centrul activ si centrul alosteric; Aici se fixeaza efectorii alosterici care nu intervin direct in cataliza, dar pot influenta procesul catalitic prin modificarea conformatiei spatiale a enzimei

34
Centrul activ al enzimelor

35
Specificitatea enzimelor
Specificitate de substrat: absoluta, relativa si stereochimica specificitate de legatura: enzimele recunosc un anumit tip de legatura chimica Esterazele recunosc legaturile de tip esteric Tripsina recunoaste legatura dintre Arg-Lys Trombina recunoaste legatura dintre Arg-Gly Arg Lys

36
Specificitatea enzimelor
Alte exemple: Chimotripsina actioneaza asupra legaturilor peptidice la care participa AA aromatici Carboxipeptidazele scindeaza legatura peptidica cea mai apropiata de capatul C-terminal, daca ultimul AA este aromatic Glicozidazele scindeaza legaturile glicozidice si sunt specifice unei monozaharide particulare: glucozidaza, galactozidaza etc

37
Specificitatea enzimelor
Acetilcolinesteraza hidrolizeaza esterul acidului acetic cu colina Substituirea radicalului acetil din S cu alt radical alifatic diminua mult activitatea enzimatica (cu cat radicalul este mai lung Butirilcolina este hidrolizata cu o viteza de 100 ori mai mare ca acetilcolina

38
Specificitatea enzimelor
1. Specificitate de substrat relativa Enzimele actioneaza asupra unui grup de substraturi cu structura asemanatoare ADH (alcool dehidrogenaza) transforma toti alcooli cu nr mic de atomi de C
 transforma toti alcooli cu nr mic de atomi de C.")
39
Specificitatea enzimelor
1. Specificitate de substrat relativa. Alte exemple

40
Specificitatea enzimelor
2. Specificitate de substrat absoluta O enzima actioneaza asupra unui singur substrat Ureaza, arginaza

41
Specificitatea enzimelor
2. Specificitate de substrat absoluta

42
Specificitatea enzimelor
3. Stereospecificitate Specificitatea stereochimica. Existenta unor compusi biologici sub forma de izomeri determina posibilitatea actiunii selective a unor enzime asupra unuia dintre ei Enzima leaga substratul pe baza complementaritatii intre centrul activ si un fragment din molecula, respectata numai in cazul unuia dintre izomeri Celalalt izomer avand o dispozitie spatiala diferita nu se poate lega de enzima.

43
Specificitatea enzimelor
Stereospecificitate Enzimele pot sa determine: transformarea unui anumit izomer geometric (cis sau trans); formarea unui anumit izomer geometric sau optic; transformarea unui substrat apartinand unei anumite serii sterice D sau L.
; formarea unui anumit izomer geometric sau optic; transformarea unui substrat apartinand unei anumite serii sterice D sau L.")
44
Specificitatea enzimelor
Stereospecificitate

45
Specificitatea enzimelor
Stereospecificitate Lactat dehidrogenaza (LDH) este o enzima care determina transformarea acidului L-lactic dar nu si pe cea a acidului D-lactic Determina transformarea unui compus optic inactiv, acidul piruvic, intr-un compus optic activ, acidul L-lactic, realizand o sinteza asimetrica L-aminoacid oxidaza care catalizeaza reactiile de dezaminare a L-aminoacizilor si D-aminoacid oxidaza care transforma D-aminoacizii;
 este o enzima care determina transformarea acidului L-lactic dar nu si pe cea a acidului D-lactic. Determina transformarea unui compus optic inactiv, acidul piruvic, intr-un compus optic activ, acidul L-lactic, realizand o sinteza asimetrica. L-aminoacid oxidaza care catalizeaza reactiile de dezaminare a L-aminoacizilor si D-aminoacid oxidaza care transforma D-aminoacizii;")
46
Specificitatea enzimelor
Succinat dehidrogenaza (SDH) catalizeaza preferential dehidrogenarea acidului succinic cu formarea acidului fumaric (izomerul trans).
 catalizeaza preferential dehidrogenarea acidului succinic cu formarea acidului fumaric (izomerul trans).")
47
Specificitatea enzimelor
Specificitate de actiune Enzimele catalizeaza un anumit tip de reactie, in functie de modul de actiune asupra substratului Acest tip de specificitate sta la baza clasificarii enzimelor si este datorat cofactorului care guverneaza tipul de reactie in care se va angaja substratul.

48
Specificitatea enzimelor
De exemplu, un anumit α-aminoacid poate fi substrat pentru mai multe enzime care au cofactori diferiti.

49
Mecanismul de actiune al enzimelor

50
Mecanismul de actiune al enzimelor
Intr-o reactie S P, la un moment dat un anumit numar de molecule de substrat au energia necesara sa atinga o stare reactiva, denumita stare de tranzitie, intermediara intre S si P si instabila Aceasta poate fie sa se transforme in produsul P fie sa reformeze substratul si se situeaza la punctul maxim al diagramei energetice ce caracterizeaza relatia intre S si P

52
Mecanismul de actiune al enzimelor
Reactia se desfasoara cu o viteza proportionala cu concentratia moleculelor de substrat ce au energia necesara pentru a realiza tranzitia Cu cat este mai mare aceasta energie fata de energia medie cu atat sunt mai putine molecule capabile sa realizeze tranzitia si reactia se desfasoara mai greu Bariera de energie dintre energia starii de tranzitie si energia libera medie a lui S se numeste energie de activare.

53
Mecanismul de actiune al enzimelor

54
Mecanismul de actiune al enzimelor

55
Mecanismul de actiune al enzimelor
Enzimele actioneaza prin scaderea energiei de activare si nu prin cresterea energiei reactantilor, combinandu-se cu substratul astfel incat sa permita trecerea acestuia in starea de tranzitie Complexele formate ES si EP sunt intermediari avand energia punctelor minime in diagrama variatiei energiei dintre S si P Energia de activare este o bariera energetica pentru reactiile chimice, moleculele cu o energie de activare mare fiind mai stabile. Fara aceasta bariera energetica complexele macromoleculare ar reveni spontan la moleculele simple ce le compun si nu ar putea sa existe procesele metabolice si structurile ordonate.

56
Mecanismul de actiune al enzimelor
Enzimele nu acţionează toate după un singur mecanism de reacţie universal valabil Dintre numeroasele mecanisme de acţiune propuse, cele mai importante sunt: cataliza acido-bazică, cataliza covalentă cataliza prin ioni metalici cataliza prin distorsiune.

57
Mecanismul de actiune al enzimelor
Cataliza prin ioni metalici

58
Mecanismul de actiune al enzimelor
Cataliza acido-bazica Este cel mai frecvent mecanism întâlnit în cataliza enzimatică Consta in procese de transfer de protoni intre gruparile acido-bazice ale enzimei si cele ale substratului În calitate de catalizatori acido-bazici generali pot funcţiona grupările carboxilice sau aminice libere ale resturilor aminoacizilor acizi sau bazici; gruparea tiol a cisteinei, gruparea imidazol a histidinei şi gruparea hidroxil a tirozinei.

59
Mecanismul de actiune al enzimelor
Cataliza acido-bazica Viteza reactiilor catalizate de acizi sau baze este influentata de doi factori importanti: Taria acizilor sau a bazelor: gruparea imidazol a histidinei are pka=6, actionand ca donor/acceptor la pH fiziologic Viteza cu care acidul sau baza cedeaza/accepta protoni; si din acest punct de vedere este eficienta His In chimotripsina, aminoacizii Asp 102 si His 57 functioneaza ca grupe cu caracter acid si, respectiv caracter bazic.

60
Mecanismul de actiune al enzimelor
Cataliza covalenta Formarea complexului enzimă-substrat presupune formarea unei legături puternice, de tip covalent (punere in comun a electronilor), între S şi E In acest tip de cataliza, atacul unei grupe electrofile sau nucleofile din situsul activ al enzimei asupra substratului determina atasarea acestuia prin legaturi covalente la enzima. Multe enzime formeaza legaturi covalente cu substratul, labilizandu-l fata de cel in forma nelegata
, între S şi E. In acest tip de cataliza, atacul unei grupe electrofile sau nucleofile din situsul activ al enzimei asupra substratului determina atasarea acestuia prin legaturi covalente la enzima. Multe enzime formeaza legaturi covalente cu substratul, labilizandu-l fata de cel in forma nelegata.")
61
Mecanismul de actiune al enzimelor
Cataliza covalenta Serin proteazele (tripsina, chimotripsina, trombina) actioneaza dupa acest mecanism. Inainte de a se lega la enzima, substratul adopta o stare de tranzitie caracterizata prin entropie scazuta, pentru care enzima are afinitate mai mare
 actioneaza dupa acest mecanism. Inainte de a se lega la enzima, substratul adopta o stare de tranzitie caracterizata prin entropie scazuta, pentru care enzima are afinitate mai mare.")
62
Enzime care actioneaza prin cataliza covalenta

63
Mecanismul de actiune al enzimelor
Cataliza prin distorsiune Se aplică în special la enzimele hidrolitice Accelerarea vitezei de reacţie prin acest mecanism s-ar datora inducerii unei tensiuni în molecula substratului, ceea ce ar determina “activarea” sa.

64
Mecanismul de actiune al enzimelor
Se presupune astfel că legarea specifică a substratului de molecula enzimei induce apariţia unei tensiuni sau a unei deformări în legătura ce urmează a fi scindată Deformarea survenită permite complexului enzimă-substrat să fie mai reactiv decât substratul singur şi să se transforme mai rapid în produşii de reacţie.

65
Cinetica enzimatică

66
Cinetica enzimatică pH-ul
Pentru enzimele din organismul uman, pH-ul optim de actiune este pH-ul normal al mediului in care ele isi manifesta actiunea catalitica Centrul activ al enzimelor contine grupe ionizabile, acide sau bazice, deci modificarea pH-ului are ca efect modificarea gradului de disociere si, implicit, modificarea vitezei de reactie.

67
Cinetica enzimatică

68
Cinetica enzimatică Pentru enzimele din organismul uman pH-ul optim este intre 5-9, dar exista enzime ce actioneaza si in afara acestui interval. De exemplu, pepsina actioneaza la pH 1,5-2, fosfatazele alcaline la pH 9-10, iar fosfatazele acide la pH 4,5-5. Pepsina Colinesteraza Tripsina

69
Cinetica enzimatică Temperatura
Efectul temperaturii asupra activitatii enzimelor este rezultanta actiunii a doi factori opusi, cresterea vitezei de reactie cu temperatura si denaturarea termica a enzimelor Activitatea oricarei enzime variaza cu temperatura Temperatura optima pentru enzimele din organismul uman este C, la plante C, iar pentru cele din microorganismele din apele termale este C Determinarea activitatii unei enzime trebuie sa se faca la o temperatura intre 25-37C.

70
Cinetica enzimatică

71
Cinetica enzimatică Concentratia substratului
In reactiile cu un singur S, mentinand constanta [E], cresterea [S] determina marirea vitezei de reactie pana cand se atinge o valoare maxima peste care oricat ar creste concentratia substratului viteza reactiei enzimatice ramane nemodificata La concentratii mici de S, viteza de reactie este proportionala cu [S] (reactii de ordin1) La concentratii mari de S, viteza de reactie devine independenta de concentratia substratului (reactii de ordin 0), enzima saturandu-se cu S.
 La concentratii mari de S, viteza de reactie devine independenta de concentratia substratului (reactii de ordin 0), enzima saturandu-se cu S.")
72
Cinetica enzimatică Concentratia substratului
L. Michaelis si M. Menten au propus o teorie generala de actiune a enzimelor bazata pe ideea ca enzima si substratul se asociaza reversibil pentru a forma complexul ES. Ecuatia exprima faptul ca viteza unei reactii enzimatice este determinata de concentratia de substrat la acel moment si de constantele Km si vmax. Aspectul curbei descrise de ecuatia Michaelis-Menten este hiperbolic

73
Cinetica enzimatică

74
Cinetica enzimatică Semnificatia Km si vmax
Daca v = vmax/2 , inlocuind in ecuatia Michaelis-Menten se deduce faptul ca Km = [S], deci constanta Michaelis poate fi definita drept concentratia de substrat la care viteza de reactie este jumatate din viteza maxima Constanta Michaelis, Km, este un indice al afinitatii enzimei pentru substrat, variind invers proportional cu aceasta

75
Cinetica enzimatică Zona “a”- v creste proportional cu [s]
Enzime michaeliene Zona “a”- v creste proportional cu [s] Zona “b” – cresterea v cu [s] nu este proportionala Zona “c” – este atins Vmax la [s] infinita c b a
![Cinetica enzimatică Zona a - v creste proportional cu [s]](http://slideplayer.gr/slide/14363896/89/images/75/Cinetica+enzimatic%C4%83+Zona+a+-+v+creste+proportional+cu+%5Bs%5D.jpg "Enzime michaeliene. Zona a - v creste proportional cu [s] Zona b – cresterea v cu [s] nu este proportionala. Zona c – este atins Vmax la [s] infinita. c. b. a.")
76
Cinetica enzimatică Pentru orice E se poate defini, alaturi de Km si v max, numarul de turn-over al enzimei, ca fiind numarul de molecule de S convertite in produs de reactie pe molecula de E in unitatea de timp, atunci cand E este saturata cu S Ecuatia Lineweaver-Burk Deoarece reprezentarea ecuatiei Michaelis -Menten este curba a carei interpretare este dificila, s-au realizat reprezentari liniare ale dependentei vitezei de reactie de [S], folosite la interpretarea datelor experimentale: reprezentarea Lineweaver-Burk, reprezentarea Hanes-Woolf, etc.

77
Cinetica enzimatică Reciproca ecuatiei Michaelis-Menten

78
Cinetica enzimatică Enzimele alosterice sunt alcatuite din mai multe subunitati curba prezinta un aspect sigmoid

79
Cinetica enzimatică Influenta efectorilor enzimatici
Efectorii enzimatici sunt substante cu structuri chimice variate care aduse in mediul de reactie pot influenta activitatea enzimei care catalizeaza reactia Efectorii enzimatici pot fi activatori sau inhibitori.

80
Cinetica enzimatică zimogen Activatorii enzimatici sunt compusi care stimuleaza activitatea enzimelor. Acestia pot fi: Activatori ai unor proenzime Forma inactiva a enzimei se numeste proenzima sau zimogen, in care centrul activ al enzimei este mascat Prin eliminarea unei parti din molecula, centrul activ este demascat, enzima devenind activa Proteaze activator Clivare S

81
Proenzime sau zimogeni

82
Inhibitorii enzimatici
Inhibitorii enzimatici sunt compusi care influenteaza negativ activitatea enzimelor pe care o pot anula definitiv sau temporar Ei pot afecta situsul catalitic al enzimei sau orice alta regiune a moleculei, astfel influentand legarea substratului de enzima Actiunea inhibitorilor enzimatici este importanta pentru controlul proceselor biochimice, pentru a intelege mecanismele de actiune a unor medicamente, droguri, otravuri, pentru a urmari etape dintr-un proces metabolic.

83
Inhibitorii enzimatici
Inhibitia enzimatica Inhibitia enzimatica a fost clasificata in moduri diferite: Inhibitie ireversibila, ce se manifesta prin alterarea de durata a structurii enzimei ca urmare a unor legaturi mai puternice. Inhibitie reversibila, fiind determinata de compusii care interactioneaza cu enzima prin interactii necovalente care poate fi: competitiva necompetitiva;

84
Inhibitorii enzimatici
Inhibitie ireversibila Se formeaza un complex EI stabil, nedisociabil E + I EI (reactie ireversibila) 1.Inhibitia se accentueaza progresiv pana la inlaturarea completa a activitatii enzimatice si nu poate fi anulata prin indepartarea I 2. se datoreaza unor modificari covalente, permanente ale gruparilor functionale necesare catalizei, transformand E in molecule inactive; 3.la inceput inhibitia este incompleta, dar creste pe masura ce apar modificarile chimice respective
 1.Inhibitia se accentueaza progresiv pana la inlaturarea completa a activitatii enzimatice si nu poate fi anulata prin indepartarea I. 2. se datoreaza unor modificari covalente, permanente ale gruparilor functionale necesare catalizei, transformand E in molecule inactive; 3.la inceput inhibitia este incompleta, dar creste pe masura ce apar modificarile chimice respective.")
85
Inhibitorii enzimatici
Inhibitie ireversibila Exemple de inhibitori ireversibili: substantele toxice, acidul iodacetic, metalele grele, compusii arsenicali Acestia interactioneaza cu radicalii aa din centru activ al E, esentiali pentru structura si functia acesteia Acidul iodacetic este inhibitor al ribonucleazei la pH=5,5; se obtine un produs alchilat al His 119 sau al His12 si o forma in care ambele histidine sunt alchilate Ionii metalelor grele ca Hg2+, Pb2+ sunt I irev pentru E ce contin sulf in centrul activ

86
Inhibitorii enzimatici

87
Inhibitorii enzimatici
Inhibitia reversibila 1.Intre E si I se formeaza un complex EI care poate regenera E E + I EI Se defineste o constanta de inhibitie, Ki ca fiind constanta de disociere a complexului EI Ki = [E] x [I]/ [EI] Inhibitia reversibila poate fi: competitiva, uncompetitiva si necompetitiva

88
Inhibitori competitivi

89
Inhibitori competitivi
I competitiv prezinta analogie structurala cu S, avand astfel afinitate pentru situsul catalitic al E Apare o competitie intre E si S pentru ocuparea situsului catalitic. Legatura prin care se leaga I este de aceeasi natura cu cea prin care se leaga S. Reactiile care au loc in acest caz sunt urmatoarele: Ks Ki

90
Inhibitori competitivi
Este o inhibitie reversibila, deoarece prin cresterea [s] se indeparteaza I competitiv de la situsul catalitic, legarea I competitiv la E fiind reversibila

91
Inhibitori competitivi
Viteza de reactie atinge Vmax ca si cum I nu ar fi prezent, daca se mareste suficient de mult [S] I competitivi cresc panta curbei, deci Km creste, scazand afinitatea E pentru S Adaugarea I competitiv nu modifica Vmax, dar creste mult Km

92
Inhibitori competitivi
Exemple de I competitivi: Succinat dehidrogenaza enzima ce transforma acidul succinic in acid fumaric, este inhibata competitiv de acizii dicarboxilici: acid malonic, acid malic, acid oxalic, asemanatori structural cu substratul acestei enzime; Sulfanilaminda isi exercita actiunea bacteriostatica prin inhibitie competitiva. Sulfanilamida este asemanatoare structural cu acidul p-aminobenzoic, pe care il poate inlocui in timpul sintezei acidului folic de catre bacterii pentru care este factor de crestere. In lipsa acidului folic bacteriile mor.

93
Inhibitori competitivi
In terapia cancerului se utilizeaza analogi structurali ai bazelor purinice si pirimidinice, ce inhiba sinteza acizilor nucleici, impiedicand diviziunea celulara care este mult mai rapida la celulele tumorale = antimetaboliti De exemplu, metotrexatul este antagonist FH2 pentru dihidrofolat reductaza Biosinteza prostaglandinelor este inhibata de anumiti acizi grasi, inhibitori competitivi ai acizilor cu 20 atomi de carbon folositi ca substrat.

94
Inhibitori competitivi
Aminele cuaternare inhiba competitiv acetilcolinesteraza, E ce hidrolizeaza Ach Actiunea unor medicamente se datoreaza faptului ca sunt inhibitori competitivi Exemple de I comp

95
Inhibitori competitivi

96
Inhibiţie uncompetitiva
I nu se combina cu E libera si nu afecteaza relatia sa cu S I se combina cu complexul ES formand un complex ESI ce nu poate genera P dorit Reducerea [ES] creste afinitatea aparenta e E pt S Inhibitorii uncompetitivi sau anticompetitivi scad Km si Vmax Inhibitia uncompetitiva functioneaza cel mai bine cand [S] este mare, iar I nu tb sa fie asemanator S

97
Inhibiţie necompetitiva
Inhibitorul necompetitiv nu se leaga la acelasi situs cu substratul, nu prezinta asemanare structurala cu acesta, deci efectul acestuia nu poate fi inlaturat prin cresterea concentratiei de substrat Inhibitorul necompetitiv interactioneaza cu complexul ES sau si cu complexul ES si cu enzima. Reactiile care au loc in acest caz sunt urmatoarele: ESI nu se poate Scinda pt a Forma P

98
Inhibiţie necompetitiva
Vmax scade Km = ct

99
Inhibiţie necompetitiva
In cazul inhibitiei necompetitive, I chiar daca nu se leaga la centru activ al E actioneaza asupra acesteia deformand-o incat nu mai poate forma cES la viteza normala, iar cES format nu se mai descompune cu viteza normala pt a forma P Inhibitia nu poate fi inlaturata prin cresterea [S]

100
Inhibiţie necompetitiva
Tipuri de inhibitori necompetitivi: ioni sau molecule ce actioneaza la nivelul cofactorilor: CN-CO; pentru citocromi I intervin in lnatul resp. mitoc inhibitori ai grupelor -SH libere ale enzimei: acid iod acetic, p-clormercuribenzoat; metale grele: argint, mercur, plumb, ce actioneaza la nivelul grupelor -SH ale enzimei; agenti de chelatare: EDTA.

101
Efectul inhibitorilor asupra vitezei de reactie

102
Enzime alosterice. Efectori alosterici
Sunt proteine oligomere alcatuite din mai multe subunitati identice sau diferite, in numar par Reactiile catalizate sunt endergonice si ireversibile; imprima sensul unic al cailor metabolice din care fac parte Intervin in prima etapa a unui lant de reactii, asigurand controlul intensitatii procesului si ireversibilitatea lui Fiecare monomer poseda un centru activ; fixarea S pe una din subunitati influienteaza legarea lui pe celelalte prin fenomenul de cooperativitate

103
Enzime alosterice. Efectori alosterici
Pe langa centri activi, monomerii prezinta si centri alosterici de care se vor lega efectorii alosterici Efectorii alosterici sunt compusi cu masa moleculara mica, fara analogie cu S si care activeaza reactiile enzimatice (efectori pozitivi) sau le inhiba (efectori negativi) Efectorii alosterici sunt prezenti la locul de actiune al E variind doar concentratia lor E alosterice sunt inhibate de produsul de reactie prin retroinhibitie sau inhibitie feedback
 sau le inhiba (efectori negativi) Efectorii alosterici sunt prezenti la locul de actiune al E variind doar concentratia lor. E alosterice sunt inhibate de produsul de reactie prin retroinhibitie sau inhibitie feedback.")
104
Efectori alosterici

105
Reglarea activitatii enzimatice

106
Reglarea activitatii enzimatice

107
Reglarea activitatii enzimatice
Enzimele modulate prin fosforilare-defosforilare Trec din forma activa in inactiva si invers prin procese de fosforilare-defosforilare la nivelul gruparilor OH ale unui rest de serina, mai rar treonina sau tirozina (nu fac parte din centrul activ) Fosforilarea se realizeaza pe seama ATP, care transfera un singur rest de acid fosforic, cu participarea unor kinaze Trasnformarea inversa are loc prin hidroliza cu participarea unor fosfataze specifice
 Fosforilarea se realizeaza pe seama ATP, care transfera un singur rest de acid fosforic, cu participarea unor kinaze. Trasnformarea inversa are loc prin hidroliza cu participarea unor fosfataze specifice.")
108
Reglarea activitatii enzimatice

109
Reglarea activitatii enzimatice
Enzimele modulate prin fosforilare-defosforilare: Glicogenfosforilaza, citratliaza active in forma fosforilata si inactive in forma defosforilata Glicogensintetaza, piruvatdehidrogenaza, acetilCoAcarboxilaza active in forma defosforilata si inactive in forma fosforilata

110
Reglarea activitatii enzimatice
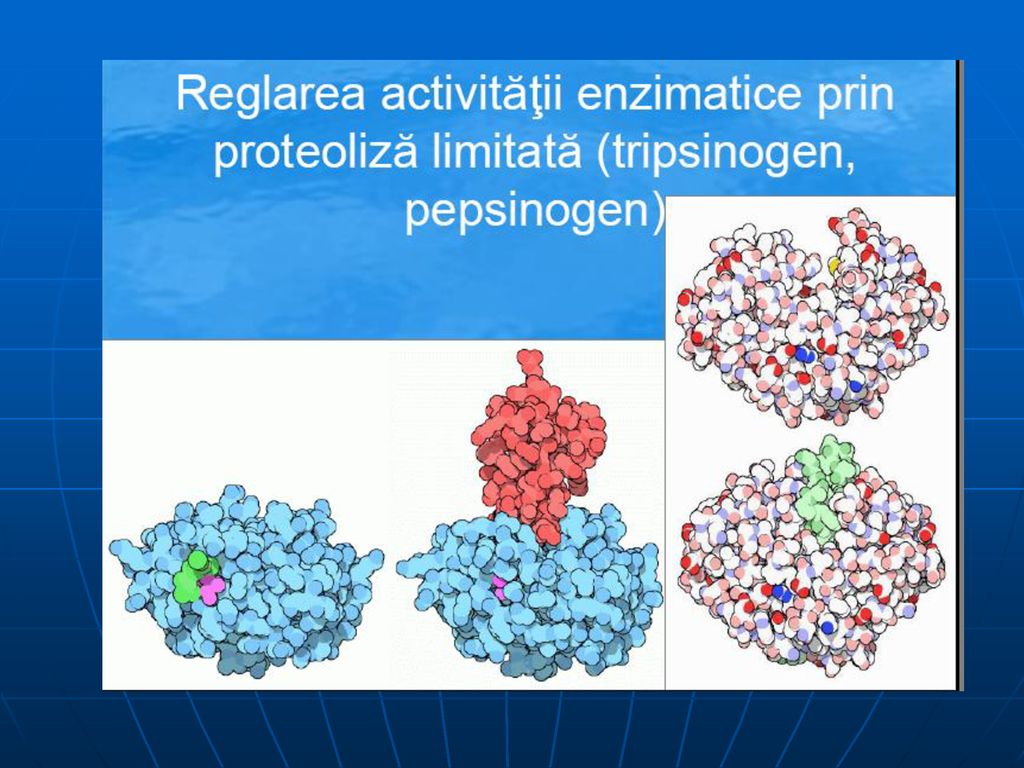
Proteoliza incompleta: pepsina, chimotripsina, tripsina, carboxipeptidazele Reglarea activitatii enzimatice prin activarea zimogenilor sau a proenzimelor la locul de actiune (stomac, intestin subtire) sub actiune aunor factori locali (pH, enzime) sau autocatalitic Transformarile sunt unidirectionale, neexistand posibilitatea refacerii zimogenului din enzimele active
 sub actiune aunor factori locali (pH, enzime) sau autocatalitic. Transformarile sunt unidirectionale, neexistand posibilitatea refacerii zimogenului din enzimele active.")
112
Reglarea activitatii enzimatice
Proteoliza incompleta. Importanta: Sinteza E la locul de actiune ar fi mult mai lenta Sintetizate in forma activa, ele si-ar putea manifesta activitatea imediat, la locul de sinteza, determinand aparitia unor boli grave Transportul lor de la locul de sinteza la locul de actiune in forma activa ar putea avea consecinte grave

114
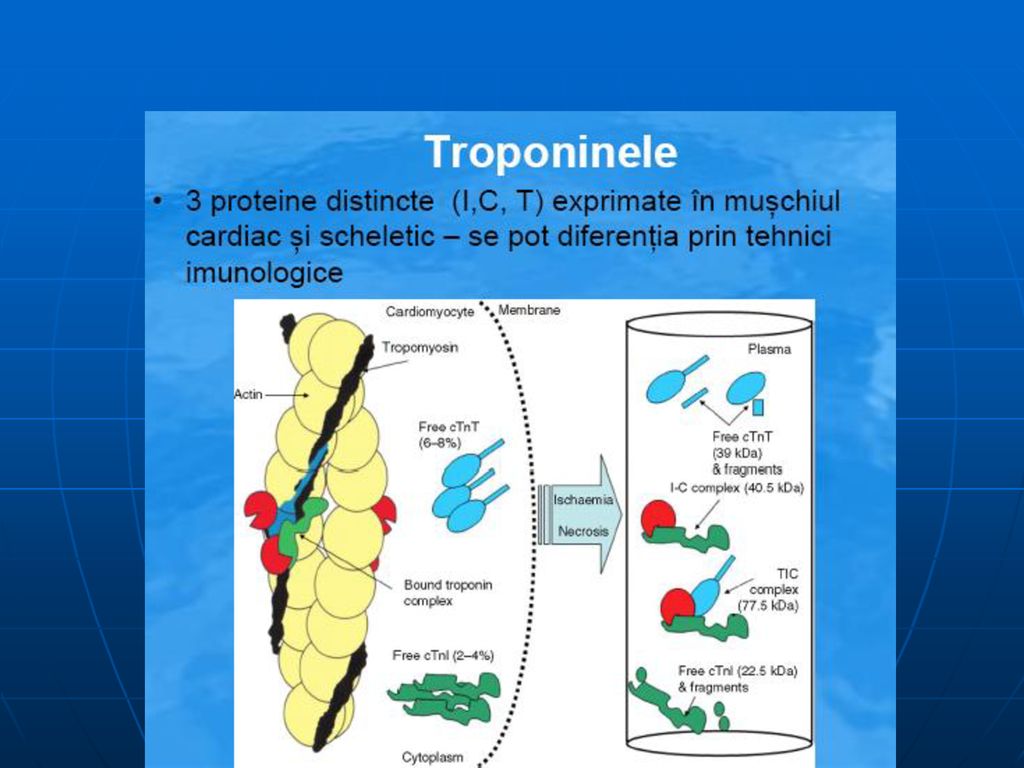
Izoenzime

115
Izoenzime

117
Izoformele lactat dehidrogenazei

123
Izoenzime

124
Izoenzime

125
Izoenzime

126
Izoenzime

























>")










