Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
ΥΠΟΔΟΧΕΙΣ G-ΠΡΩΤΕΪΝΩΝ (GPCRs)
Καραβασίλης Θεόδωρος Καραγεώργος Χρήστος Καρατζιά Δέσποινα ΙΑΤΡΙΚΗ ΣΧΟΛΗ Α.Π.Θ. 11/12/2013

2
ΠΕΡΙΕΧΟΜΕΝΑ Εισαγωγή Ρόλοι των GPCRs στη Φυσιολογία
Μηχανισμοί G πρωτεΐνες Δομή Τρόποι Λειτουργίας Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) Η οδός του κυκλικού AMP Η οδός των φωσφολιπιδίων ινοσιτόλης Μηχανισμοί τερματισμού σηματοδότησης Ρύθμιση των GPCRs Βιβλιογραφία
 Η οδός του κυκλικού AMP. Η οδός των φωσφολιπιδίων ινοσιτόλης. Μηχανισμοί τερματισμού σηματοδότησης. Ρύθμιση των GPCRs. Βιβλιογραφία.")
3
ΕΙΣΑΓΩΓΗ (I) GPCRs Μεγαλύτερη πρωτεϊνική οικογένεια υποδοχέων
Μεγαλύτερη πρωτεϊνική οικογένεια υποδοχέων Πρόσδεση με σηματοδοτικά μόρια επάγει την ενεργοποίηση πορειών μεταγωγής σήματος Αποκαλούνται 7-διαμεμβρανικοί υποδοχείς (7 ΤΜ receptors) ή επτά-ελικοειδείς υποδοχείς Απαντώνται μόνο στα ευκαρυωτικά κύτταρα και όχι στα βακτήρια
 ή επτά-ελικοειδείς υποδοχείς. Απαντώνται μόνο στα ευκαρυωτικά κύτταρα και όχι στα βακτήρια.")
4
ΕΙΣΑΓΩΓΗ (II) Πορείες μεταγωγής σήματος
Οδός κυκλικού ΑΜΡ Οδός φωσφολιπιδίων ινοσιτόλης Ταξινόμηση σε 6 κατηγορίες με βάση την ομολογία αλληλουχίας και την λειτουργική ομοιότητα Μοιράζονται κοινή δομή και μηχανισμό μεταγωγής σήματος Υπάρχουν και κάποιοι υποδοχείς με άγνωστη λειτουργία

5
Ρόλοι των GPCRs στη Φυσιολογία (Ι)
1.Όραση Όλα τα φωτοευαίσθητα όργανα βασίζονται σε συστήματα φωτοϋποδοχέων, τα οποία χρησιμοποιούν μια οικογένεια πρωτεϊνών που λέγονται οψίνες. Χαρακτηριστικό παράδειγμα: ροδοψίνη (σύμπλοκο οψίνης και ρετινάλης) Η μετάβαση από την ανενεργή στην ενεργή μορφή της ρετινάλης έχει ως αποτέλεσμα αρχικά την ενεργοποίηση μιας G-πρωτεΐνης, της τρανσδουσίνης, και μέσω μιας πολύπλοκης ενδοκυττάριας σηματοδοτικής ακολουθίας, το φως που αρχικά προσέπεσε στον αμφιβληστροειδή χιτώνα του οφθαλμού μετατρέπεται τελικά σε μια νευρική ώση.
 Η μετάβαση από την ανενεργή στην ενεργή μορφή της ρετινάλης έχει ως αποτέλεσμα αρχικά την ενεργοποίηση μιας G-πρωτεΐνης, της τρανσδουσίνης, και μέσω μιας πολύπλοκης ενδοκυττάριας σηματοδοτικής ακολουθίας, το φως που αρχικά προσέπεσε στον αμφιβληστροειδή χιτώνα του οφθαλμού μετατρέπεται τελικά σε μια νευρική ώση.")
6
Ρόλοι των GPCRs στη Φυσιολογία (ΙΙ)
2.Γεύση Οι GPCRs στα κύτταρα που σχετίζονται με την αίσθηση της γεύσης διαμεσολαβούν για την απελευθέρωση της γκαστδουσίνης (G-πρωτεΐνη). Η έκκριση της συγκεκριμένης πρωτεΐνης προκαλείται όταν τα προαναφερθέντα κύτταρα έρθουν σε επαφή με πικρές και γλυκιές γευστικά ουσίες, και αυτή μεταβιβάζει στη συνέχεια τα ανάλογα σήματα στον οργανισμό μέσω συγκεκριμένων σηματοδοτικών οδών.
. Η έκκριση της συγκεκριμένης πρωτεΐνης προκαλείται όταν τα προαναφερθέντα κύτταρα έρθουν σε επαφή με πικρές και γλυκιές γευστικά ουσίες, και αυτή μεταβιβάζει στη συνέχεια τα ανάλογα σήματα στον οργανισμό μέσω συγκεκριμένων σηματοδοτικών οδών.")
7
Ρόλοι των GPCRs στη Φυσιολογία (ΙΙI)
3.Όσφρηση Οι υποδοχείς του οσφρητικού επιθηλίου δεσμεύουν οσμές και φερομόνες. Στα θηλαστικά, η πρόσδεση αυτών των συνδετών στους υποδοχείς ενεργοποιεί την αδενυλική κυκλάση (η οποία συνθέτει cAMP) μέσω μιας G-πρωτεΐνης που ονομάζεται Golf
 μέσω μιας G-πρωτεΐνης που ονομάζεται Golf.")
8
Ρόλοι των GPCRs στη Φυσιολογία (ΙV)
4. Ρύθμιση συμπεριφοράς και διάθεσης Οι GPCRs στους εγκεφάλους των θηλαστικών δεσμεύουν αρκετούς διαφορετικούς νευροδιαβιβαστές, οι οποίοι είναι ενδογενείς χημικές ουσίες που μεταβιβάζουν σήματα από έναν νευρώνα σε κύτταρα-στόχους μέσω των συνάψεων.

9
Ρόλοι των GPCRs στη Φυσιολογία (V)
5. Ρύθμιση της δραστηριότητας του ανοσοποιητικού συστήματος και της φλεγμονής Υποδοχείς χημειοκίνης δεσμεύουν συνδέτες που διαμεσολαβούν στη μεσοκυττάρια επικοινωνία μεταξύ των κυττάρων του ανοσοποιητικού συστήματος. Η πρόσδεση αυτή έχει ως αποτέλεσμα ποικίλες κυτταρικές αντιδράσεις. Υποδοχείς ισταμίνης δεσμεύουν ορισμένους φλεγμονώδεις μεσολαβητές και εμπλέκουν κατ’ αυτόν τον τρόπο συγκεκριμένα κύτταρα-στόχους στη φλεγμονώδη αντίδραση.

10
Ρόλοι των GPCRs στη Φυσιολογία (VI)
6.Αυτόνομο νευρικό σύστημα μεταφοράς Τόσο το συμπαθητικό όσο και το παρασυμπαθητικό νευρικό σύστημα ρυθμίζονται από πορείες ελεγχόμενες από GPCRs, υπεύθυνες για τον έλεγχο πολλών αυτόματων λειτουργιών του σώματος, όπως η αρτηριακή πίεση, ο καρδιακός ρυθμός και οι πεπτικές διεργασίες. 7.Ρύθμιση της κυτταρικής πυκνότητας 8.Διαμόρφωση ομοιόστασης (π.χ. ρύθμιση και διατήρηση υδατικού ισοζυγίου) 9.Εμπλέκονται στην ανάπτυξη και μετάσταση ορισμένων τύπων όγκων
 9.Εμπλέκονται στην ανάπτυξη και μετάσταση. ορισμένων τύπων όγκων.")
11
Δομή και Σχέσεις δομής-λειτουργίας των GPCRs (Ι)
7 διαμεμβρανικές α-έλικες (TM-1 to TM-7) Στον εξωκυττάριο χώρο βρίσκεται το αμινικό άκρο (N-terminus) ενώ στον ενδοκυττάριο το καρβοξυλικό άκρο (C-terminus). Οι α-έλικες συνδέονται μεταξύ τους μέσω τριών εξωκυττάριων (EL-1 to EL-3) και τριών ενδοκυττάριων (IL-1 to IL-3) βρόγχων. Η τριτοταγής δομή των GPCRs μοιάζει με «βαρέλι» στο εσωτερικό του οποίου σχηματίζεται μια κοιλότητα όπου προσδένεται ο συνδέτης. Για πιο ογκώδεις συνδέτες εντοπίζονται και κάποιες άλλες θέσεις πρόσδεσης. Τέτοιες χαρακτηριστικές θέσεις παρατηρούνται στην 3η κλάση των GPCRs.
 Στον εξωκυττάριο χώρο βρίσκεται το αμινικό άκρο (N-terminus) ενώ στον ενδοκυττάριο το καρβοξυλικό άκρο (C-terminus). Οι α-έλικες συνδέονται μεταξύ τους μέσω τριών εξωκυττάριων (EL-1 to EL-3) και τριών ενδοκυττάριων (IL-1 to IL-3) βρόγχων. Η τριτοταγής δομή των GPCRs μοιάζει με «βαρέλι» στο εσωτερικό του οποίου σχηματίζεται μια κοιλότητα όπου προσδένεται ο συνδέτης. Για πιο ογκώδεις συνδέτες εντοπίζονται και κάποιες άλλες θέσεις πρόσδεσης. Τέτοιες χαρακτηριστικές θέσεις παρατηρούνται στην 3η κλάση των GPCRs.")
14
Δομή και Σχέσεις δομής-λειτουργίας των GPCRs (ΙΙ)
Όταν ένας συνδέτης, προσδεθεί στην κατάλληλη θέση επέρχεται μεταβολή στη στερεοδιαμόρφωση του υποδοχέα. Συγκεκριμένα, έχουμε αλλαγή στους σχετικούς προσανατολισμούς των ΤΜ ελίκων η οποία οδηγεί στη δημιουργία μιας φαρδύτερης επιφάνειας στον ενδοκυττάριο χώρο και στην αποκάλυψη καταλοίπων και άλλων περιοχών των ΤΜ ελίκων, απαραίτητα για τη σηματοδότηση.

15
Δομή και Σχέσεις δομής-λειτουργίας των GPCRs (ΙΙI)
Η φωσφορυλίωση καταλοίπων σερίνης και θρεονίνης του καρβοξυλικού άκρου που ευνοεί τη σύνδεση των β-αρρεστινών (β-arr) Η παλμιτοϋλίωση τμημάτων του καρβοξυλικού άκρου που έχει σαν αποτέλεσμα τη στοχοποίηση των υποδοχέων από τις «σχεδίες» λιπιδίων (lipid rafts)
 Η παλμιτοϋλίωση τμημάτων του καρβοξυλικού άκρου που έχει σαν αποτέλεσμα τη στοχοποίηση των υποδοχέων από τις «σχεδίες» λιπιδίων (lipid rafts)")
16
Δομή και Σχέσεις δομής-λειτουργίας των GPCRs (ΙV)
Γενικά, οι GPCRs ανταποκρίνονται σε διάφορα εξωκυττάρια σήματα στα οποία μεσολαβούν ποικίλοι συνδέτες που όμως όλοι μεταδίδουν αυτό το σήμα μέσω μηχανισμών σύνδεσης με κάποια G-πρωτεΐνη. Αυτό έχει καταστεί δυνατό με βάση έναν τομέα GEF που σχηματίζεται κυρίως από το συνδυασμό των IL-2 και IL-3 και με παρακείμενα κατάλοιπα των ΤΜ ελίκων.

17
Μηχανισμοί (Ι) Δύο είναι οι κύριοι μηχανισμοί που χαρακτηρίζουν τη λειτουργία των GPCRs: Η πρόσδεση του συνδέτη Η μεταβολή της στερεοδιαμόρφωσης Είδη συνδετών: Μόρια που διεγείρουν τις αισθήσεις Διάφορα πεπτίδια (π.χ. νευροπεπτίδιο Υ, οψίνες, γ-αμινοβουτυρικό οξύ [GABA] κ.ά.) Βιογενείς αμίνες (π.χ. ντοπαμίνη, επινεφρίνη κ.ά.) Ορμόνες (π.χ. καλσιτονίνη, νευροκινίνη κ.ά.) Συνδέτες λιπιδικής φύσεως (π.χ. προσταγλανδίνες κ.ά.) Χημειοκίνες
 Βιογενείς αμίνες (π.χ. ντοπαμίνη, επινεφρίνη κ.ά.) Ορμόνες (π.χ. καλσιτονίνη, νευροκινίνη κ.ά.) Συνδέτες λιπιδικής φύσεως (π.χ. προσταγλανδίνες κ.ά.) Χημειοκίνες.")
18
Μηχανισμοί (ΙΙ) Μεταβολή στερεοδιαμόρφωσης
Θεωρείται ότι οι υποδοχείς βρίσκονται σε μια διαμορφωτική ισορροπία μεταξύ ενεργούς και ανενεργούς βιοφυσικής κατάστασης. Υπάρχουν τρείς κατηγορίες συνδετών : Οι «αγωνιστές» (agonists), που μετατοπίζουν την ισορροπία προς την ενεργή κατάσταση , Οι «αντίστροφοι αγωνιστές» (inverse agonists), που μετατοπίζουν την ισορροπία προς την ανενεργή κατάσταση, και Οι «ουδέτεροι ανταγωνιστές» (neutral antagonists), που δεν επηρεάζουν αυτή την ισορροπία *** Παρ’ όλα αυτά δεν είναι ακόμα γνωστό που έγκειται η ακριβής διαφορά μεταξύ ενεργούς και ανενεργούς κατάστασης***
, που μετατοπίζουν την ισορροπία προς την ενεργή κατάσταση , Οι «αντίστροφοι αγωνιστές» (inverse agonists), που μετατοπίζουν την ισορροπία προς την ανενεργή κατάσταση, και. Οι «ουδέτεροι ανταγωνιστές» (neutral antagonists), που δεν επηρεάζουν αυτή την ισορροπία. *** Παρ’ όλα αυτά δεν είναι ακόμα γνωστό που έγκειται η ακριβής διαφορά μεταξύ ενεργούς και ανενεργούς κατάστασης***")
19
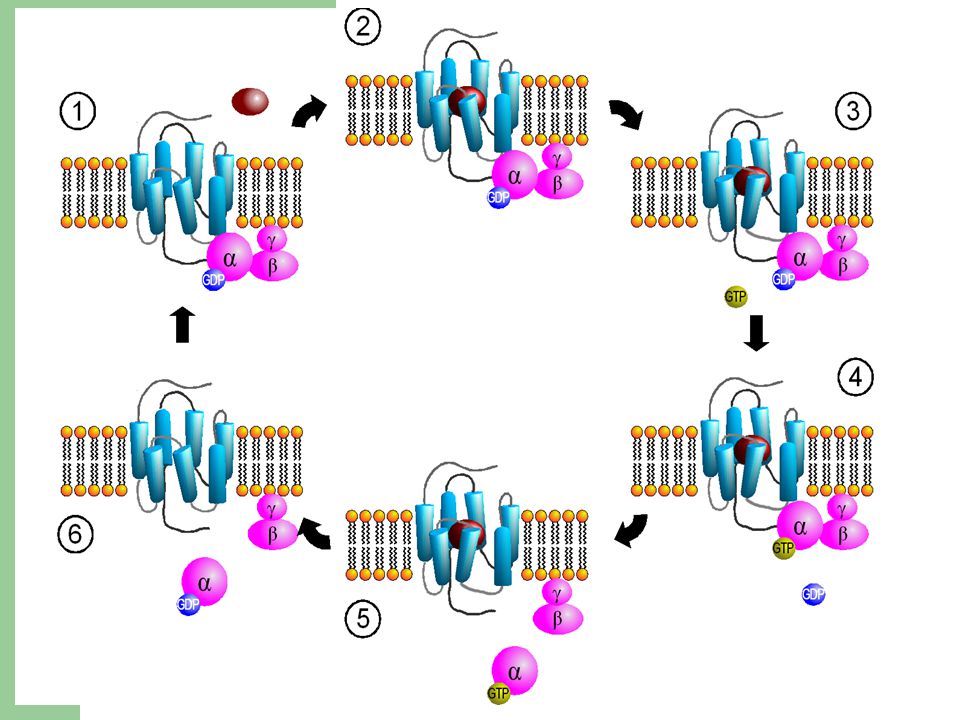
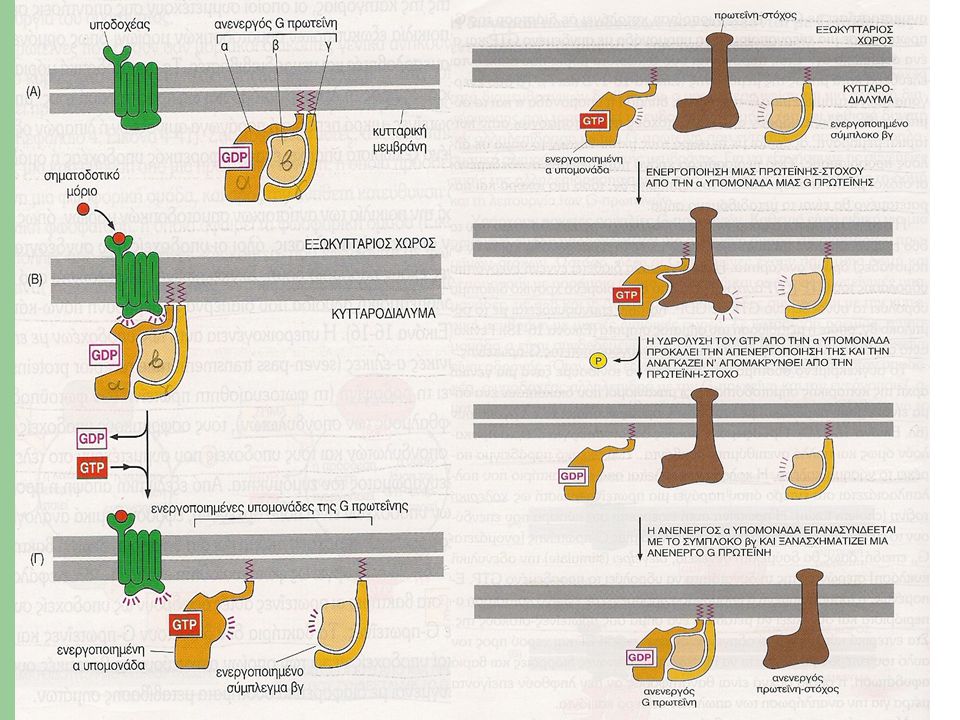
G πρωτεΐνες (I) ΔΟΜΗ Υπάρχουν ποικίλες κατηγορίες G-πρωτεϊνών, η καθεμιά από τις οποίες είναι ειδική για μια συγκεκριμένη ομάδα υποδοχέων και μια συγκεκριμένη ομάδα ενδοκυττάριων πρωτεϊνών-στόχων. Ωστόσο, όλες οι G-πρωτεΐνες έχουν παρόμοια δομή και λειτουργούν με παρόμοιο τρόπο. Απαρτίζονται από τρεις πρωτεϊνικές υπομονάδες, τις α, β και γ, γι’ αυτό ονομάζονται και ετεροτριμερείς. Οι α και γ συνδέονται με την κυτταρική μεμβράνη με μικρές ουρές λιπιδικής φύσεως. Σε ανενεργή κατάσταση, η υπομονάδα α είναι συνδεδεμένη με GDP και η G-πρωτεΐνη παραμένει αδρανής.

20
G πρωτεΐνες (IΙ) ΤΡΟΠΟΙ ΛΕΙΤΟΥΡΓΙΑΣ
Όταν ένας εξωκυττάριος συνδέτης προσδεθεί στον υποδοχέα, προκαλεί την αλληλεπίδραση του με την G-πρωτεΐνη και εν τέλει την ενεργοποίηση της. Η πρόσδεση αυτή προκαλεί αλλαγή στη διαμόρφωση του υποδοχέα, που δρα ως παράγοντας ανταλλαγής νουκλεοτιδίων γουανίνης GEF (Guanine nucleotide Exchange Factor). Κατά αυτόν τον τρόπο, η α υπομονάδα δύναται να αποβάλλει το συνδεδεμένο σε αυτήν GDP και να το αντικαταστήσει με GTP.
. Κατά αυτόν τον τρόπο, η α υπομονάδα δύναται να αποβάλλει το συνδεδεμένο σε αυτήν GDP και να το αντικαταστήσει με GTP.")
22
G πρωτεΐνες (IΙI) H ενεργοποίηση καταλήγει σε διάσταση της G-πρωτεΐνης σε μια α υπομονάδα με συνδεδεμένο GTP (ενεργοποιημένη) και σε ένα σύμπλοκο βγ. Τα δύο ενεργοποιημένα τμήματα της G-πρωτεΐνης αλληλεπιδρούν άμεσα με στόχους που εντοπίζονται στην κυτταρική μεμβράνη. Αυτοί οι στόχοι μεταβιβάζουν με τη σειρά τους το σήμα στους επιθυμητούς προορισμούς.

23
G πρωτεΐνες (IV) Η ανεξάρτητη δράση των δύο ενεργοποιημένων τμημάτων της G-πρωτεΐνης καθορίζεται από τη συμπεριφορά της α υπομονάδας. Και αυτό, διότι η α υπομονάδα διαθέτει εγγενή ενεργότητα υδρολάσης του GTP (GTPάση) και μετά από κάποιο χρονικό διάστημα υδρολύει το συνδεδεμένο GTP σε GDP. Έπειτα επανασυνδέεται με το σύμπλοκο βγ, οπότε το σήμα παύει να μεταδίδεται.
 και μετά από κάποιο χρονικό διάστημα υδρολύει το συνδεδεμένο GTP σε GDP. Έπειτα επανασυνδέεται με το σύμπλοκο βγ, οπότε το σήμα παύει να μεταδίδεται.")
24
G πρωτεΐνες (V) Το συγκεκριμένο σύστημα λειτουργίας μας υπενθυμίζει μια βασική γενική αρχή της κυτταρικής σηματοδότησης: σημασία μηχανισμών που διακόπτουν ένα σήμα σημασία μηχανισμών που ενεργοποιούν ένα σήμα. Παραδείγματα νοσημάτων: χολέρα, κοκκύτης.

25
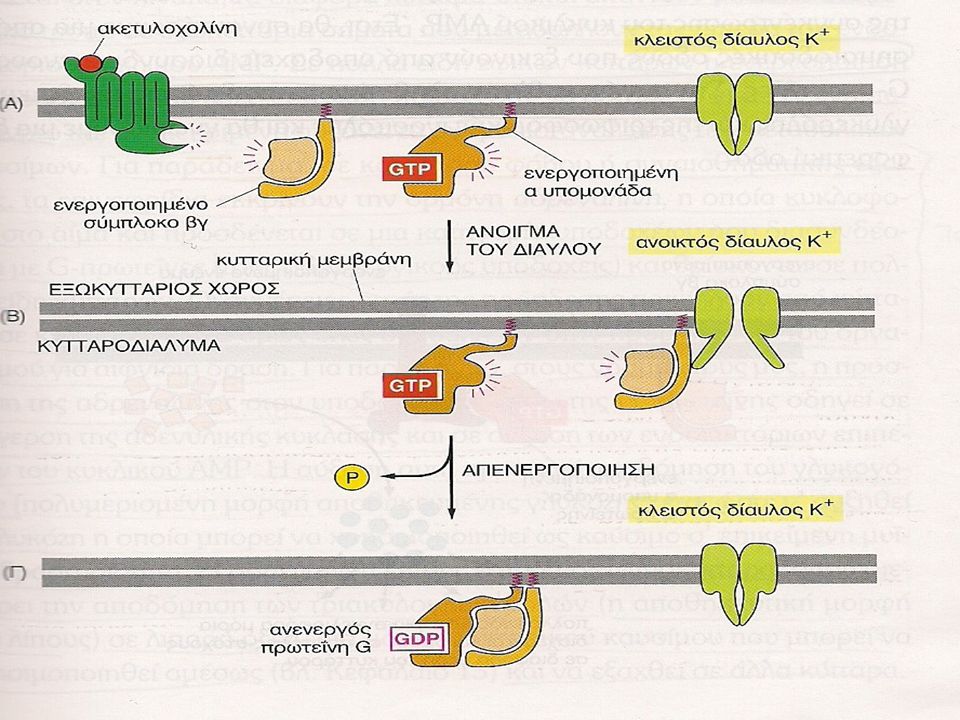
G πρωτεΐνες (VI) 2. Μερικές G-πρωτεΐνες ρυθμίζουν διαύλους ιόντων
Υπάρχουν εξωκυττάρια σηματοδοτικά μόρια που όταν προσδεθούν με έναν GPCR προκαλούν την ενεργοποίηση G-πρωτεϊνών οι οποίες έχουν στόχο διαύλους ιόντων. Χαρακτηριστικό παράδειγμα τέτοιου σηματοδοτικού μορίου αποτελεί η ακετυλοχολίνη η οποία μπορεί να ενεργοποιήσει , όταν προσδεθεί στον κατάλληλο υποδοχέα , μια G-πρωτεΐνη που προκαλεί τη διάνοιξη ενός διαύλου K+ στην κυτταρική μεμβράνη ενός μυοκαρδιακού κυττάρου.

27
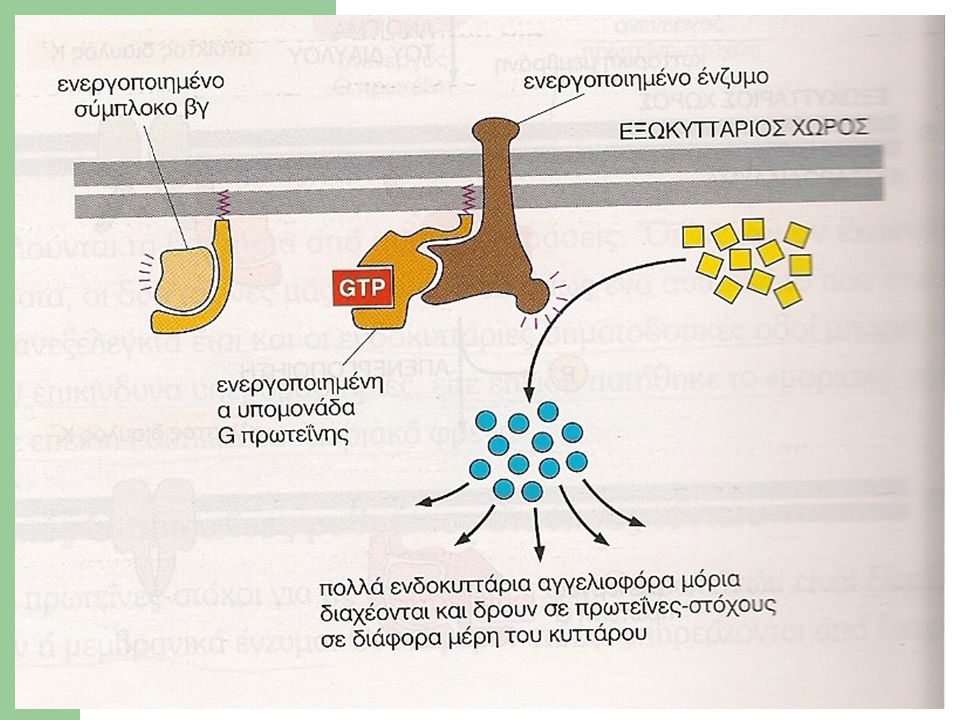
G πρωτεΐνες (VII) 3. Ορισμένες G-πρωτεΐνες ενεργοποιούν μεμβρανικά ένζυμα Τα πιο συνήθη ένζυμα-στόχοι για τις G –πρωτεΐνες είναι: Η αδενυλική κυκλάση που καταλύει την παράγωγη του cAMP (cyclic AMP) Η φωσφολιπάση C που καταλύει την σύνθεση i) τριφωσφορικής ινοσιτόλης ii) διακυλογλυκερόλης Η ενεργοποίηση των ενζύμων γίνεται από διαφορετικά είδη G πρωτεϊνών και υπό την επίδραση διαφόρων εξωκυττάριων σημάτων σχηματίζονται τα μικρά αυτά σηματοδοτικά μόρια, που αποκαλούνται δεύτεροι αγγελιοφόροι (πρώτοι είναι τα εξωκυττάρια σήματα)
 Η φωσφολιπάση C που καταλύει την σύνθεση i) τριφωσφορικής ινοσιτόλης ii) διακυλογλυκερόλης. Η ενεργοποίηση των ενζύμων γίνεται από διαφορετικά είδη G πρωτεϊνών και υπό την επίδραση διαφόρων εξωκυττάριων σημάτων σχηματίζονται τα μικρά αυτά σηματοδοτικά μόρια, που αποκαλούνται δεύτεροι αγγελιοφόροι (πρώτοι είναι τα εξωκυττάρια σήματα)")
29
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (Ι)
Η οδός του κυκλικού AMP Πολλά εξωκυττάρια σήματα επηρεάζουν την ενεργότητα της αδενυλικής κυκλάσης και επομένως μεταβάλλουν τη συγκέντρωση του cAMP. Η σύνδεση της ενεργοποιημένης α-υπομονάδας με την αδενυλική κυκλάση, ενεργοποιεί το ενζυμο με αποτέλεσμα την απότομη αύξηση της σύνθεσης του cΑΜΡ από ΑΤΡ.

30
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (ΙI)
Το κυκλικό ΑΜΡ είναι ένα υδατοδιαλυτό μόριο και προωθεί το σήμα μέσω διάχυσης από την μεμβράνη όπου συντίθεται, σε πρωτεΐνες του πυρήνα, του κυτταροπλάσματος και άλλων οργανιδίων. Τα διάφορα κύτταρα-στόχοι απαντούν με πολύ διαφορετικό τρόπο σε εξωκυττάρια σήματα που μεταβάλλουν τα επίπεδα του ενδοκυττάριου cΑΜΡ. Συνήθως η ενεργοποίηση παραγωγής του κυκλικού ΑΜΡ αυξάνει το ρυθμό κατανάλωσης των μεταβολικών ενζύμων.

31
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (ΙIΙ)
Επινεφρίδια Αδρεναλίνη Μέσω του αίματος Σύνδεση με υποδοχείς Διέγερση αδενυλικής κυκλάσης Αύξηση συγκέντρωσης cΑΜΡ

32
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (ΙV)
Γραμμωτοί μυς Λιποκύτταρα Αποδόμηση Γλυκογόνου Αποδόμηση Τριακυλογλυκερολών2 Παραγωγή Γλυκόζης Παραγωγή Λιπαρών Οξέων 1: Πολυμερισμένη μορφή αποθηκευμένης γλυκόζης 2: Αποθηκευτική μορφή του λίπους

33
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (V)
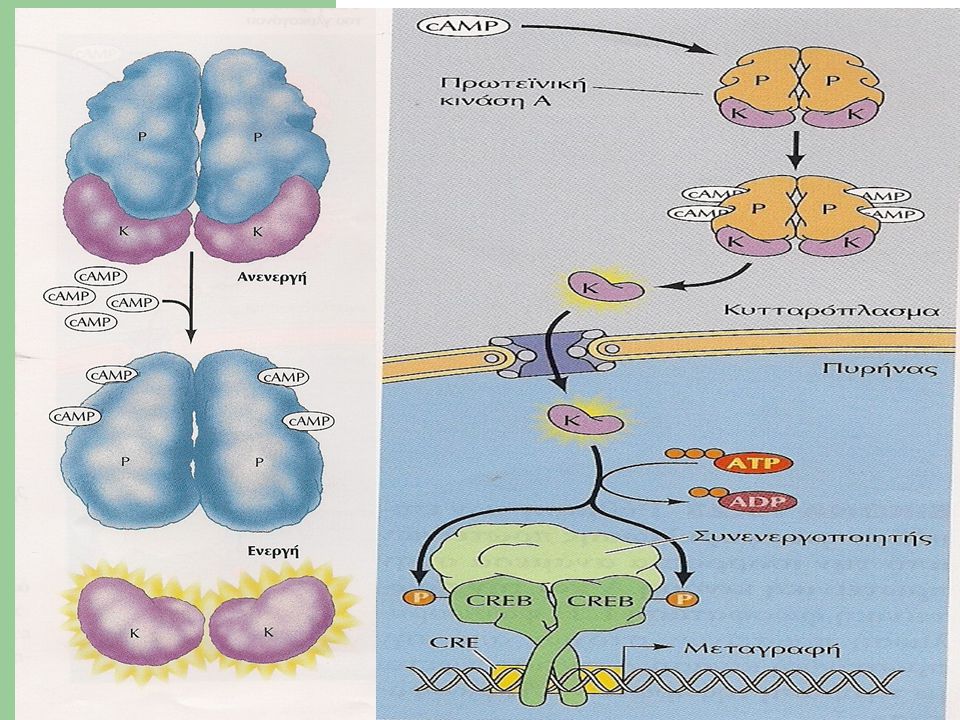
Το κυκλικό ΑΜΡ ασκεί αυτές τις δράσεις κυρίως μέσω της ενεργοποίησης της εξαρτώμενης από το κυκλικό ΑΜΡ πρωτεϊνικής κινάσης ή Α-κινάσης. Η κινάση Α φωσφορυλιώνει κατάλοιπα σερίνης ή θρεονίνης ορισμένων ενδοκυττάριων πρωτεινών , μεταβάλλοντας έτσι την δραστικότητά τους.

34
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (VI)
Σε διαφορετικά ειδή κυττάρων ,διαφορετικές πρωτεΐνες υφίστανται φωσφορυλίωση,γι’ αυτό και οι επιδράσεις του κυκλικού ΑΜΡ ποικίλουν από κύτταρο σε κύτταρο. Οι επιδράσεις του cΑΜΡ μπορεί να εξελίσσονται : Σε λίγα δευτερόλεπτα (μεταβολισμός γλυκογόνου) Σε αρκετά λεπτά ή ώρες (μεταβολή έκφρασης γονιδίων)
 Σε αρκετά λεπτά ή ώρες (μεταβολή έκφρασης γονιδίων)")
36
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (VII)
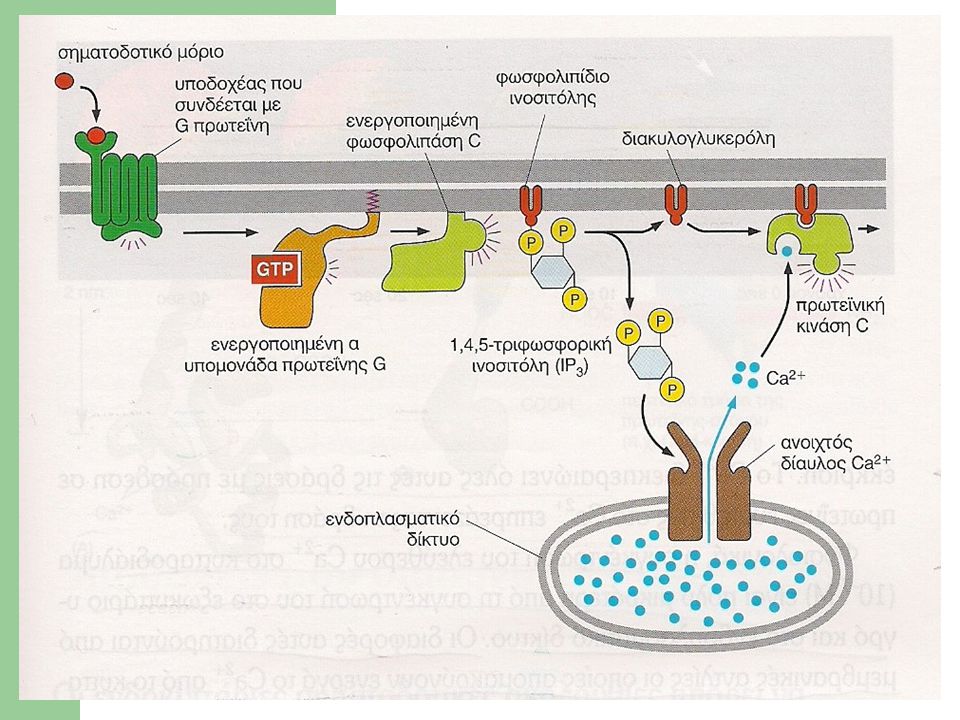
2. Η οδός των φωσφολιπιδίων ινοσιτόλης Δύο κύρια μονοπάτια ενδοκυτταρικής σηματοδότησης βασίζονται σε δεύτερους αγγελιοφόρους που παράγονται από το μεμβρανικό φωσφολιπίδιο 4,5-διφωσφορική φωσφατιδυλοϊνοσιτόλη (PIP2). Διάφορες ορμόνες και αυξητικοί παράγοντες επάγουν την υδρόλυση της PIP2 μέσω της δράσης της φωσφολιπάσης C (PLC). Η PLC αποκόπτει την υδρόφιλη φωσφοσακχαριτική κεφαλή του φωσφολιπιδίου και παράγει το φωσφορικό σάκχαρο 1,4,5-τριφωσφορική ινοσιτόλη (IP3), ενώ παράλληλα αφήνει τη λιπιδική ουρά, τη διακυλογλυκερόλη (DAG), ενσωματωμένη στην κυτταρική μεμβράνη.
. Διάφορες ορμόνες και αυξητικοί παράγοντες επάγουν την υδρόλυση της PIP2 μέσω της δράσης της φωσφολιπάσης C (PLC). Η PLC αποκόπτει την υδρόφιλη φωσφοσακχαριτική κεφαλή του φωσφολιπιδίου και παράγει το φωσφορικό σάκχαρο. 1,4,5-τριφωσφορική ινοσιτόλη (IP3), ενώ παράλληλα αφήνει τη λιπιδική ουρά, τη διακυλογλυκερόλη (DAG), ενσωματωμένη στην κυτταρική μεμβράνη.")
37
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (VIIΙ)
Η DAG και η IP3 διεγείρουν διαφορετικές ενδοκυτταρικές οδούς σηματοδότησης Η IP3 εγκαταλείπει την κυτταρική μεμβράνη και διαχέεται στο κυτταρικό διάλυμα. Όταν φτάσει στο ενδοπλασματικό δίκτυο, προσδένεται σε διαύλους Ca2+ της μεμβράνης του και προκαλεί το άνοιγμά τους. Τα ιόντα εκρέουν γρήγορα προς το κυτταρικό διάλυμα μέσω των ανοικτών διαύλων. Η συγκέντρωση των ιόντων Ca2+ στο κυτταρικό διάλυμα αυξάνει απότομα, επηρεάζοντας την ενεργότητα ποικίλων πρωτεϊνών-στόχων.

39
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (IΧ)
Η DAG παραμένει συνδεδεμένη με την κυτταροπλασματική μεμβράνη και ενεργοποιεί πρωτεϊνικές κινάσες σερίνης/θρεονίνης (οικογένεια πρωτεϊνικών κινασών C), με σημαντικό ρόλο στη ρύθμιση της κυτταρικής ανάπτυξης και διαφοροποίησης. Οι κινάσες ενεργοποιούνται μόνο με την πρόσδεση σε αυτές ιόντων Ca2+ και επακόλουθα φωσφορυλιώνουν μια ομάδα ενδοκυττάριων πρωτεϊνών που διαφέρουν ανάλογα με το είδος του κυττάρου. Ιόντα Ca2+: αύξηση της συγκέντρωσης του ελεύθερου Ca2+ στο κυτταρόπλασμα δίνει απάντηση σε πολλά διαφορετικά σήματα.
, με σημαντικό ρόλο στη ρύθμιση της κυτταρικής ανάπτυξης και διαφοροποίησης. Οι κινάσες ενεργοποιούνται μόνο με την πρόσδεση σε αυτές ιόντων Ca2+ και επακόλουθα φωσφορυλιώνουν μια ομάδα ενδοκυττάριων πρωτεϊνών που διαφέρουν ανάλογα με το είδος του κυττάρου. Ιόντα Ca2+: αύξηση της συγκέντρωσης του ελεύθερου Ca2+ στο κυτταρόπλασμα δίνει απάντηση σε πολλά διαφορετικά σήματα.")
40
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (Χ)
Στο κυτταρόπλασμα, οι επιδράσεις του Ca2+ είναι κυρίως έμμεσες και πραγματοποιούνται από ποικίλες πρωτεΐνες μεταβίβασης. Καλμοδουλίνη: Εντοπίζεται στο κυτταρικό διάλυμα όλων των ευκαρυωτικών κυττάρων. Όταν συνδεθεί με Ca2+, μεταβάλλεται η διαμόρφωση της, οπότε αποκτά την ικανότητα να προσδένεται σε διάφορες πρωτεΐνες-στόχους του κυττάρου και να τροποποιεί τη δράση τους. CaM-κινάσες: Ενεργοποιούνται από την πρόσδεση του συμπλόκου καλμοδουλίνης-Ca2+. Σε ενεργό κατάσταση, επηρεάζουν κυτταρικές διεργασίες με το μηχανισμό της φωσφορυλίωσης άλλων επιλεγμένων πρωτεϊνών.

41
Οδοί ενδοκυτταρικής μεταγωγής σήματος (Signal Pathways) (ΧΙ)
Τύποι CaM-κινασών: Ένας τύπος υπάρχει σε υψηλά επίπεδα στο νευρικό σύστημα, όπου ρυθμίζει τη σύνθεση και την απελευθέρωση νευροδιαβιβαστών. Ορισμένες ρυθμίζουν τη γονιδιακή έκφραση, φωσφορυλιώνοντας μεταγραφικούς παράγοντες. Ορισμένες ρυθμίζουν τη δράση της αδενυλικής κυκλάσης και της φωσφοδιεστεράσης.

42
Μηχανισμοί τερματισμού σηματοδότησης (I)
Οι G-πρωτεΐνες έχουν τη δυνατότητα να τερματίσουν οι ίδιες τη δράση τους λόγω της εγγενούς δράσης GTP-άσης που διαθέτουν. Όμως η διαδικασία αυτή είναι ιδιαίτερα αργή και απαιτούνται περίπου 50 sec για να απενεργοποιηθεί μια G-πρωτεΐνη αν δεν δράσει και κάποιος άλλος παράγοντας.

43
Μηχανισμοί τερματισμού σηματοδότησης (II)
Πρωτείνες RGS: Regulators of G-protein Signaling Όταν μια πρωτεΐνη RGS συνδεθει με την Ga υπομονάδα επιταχύνει έως και 1500 φόρες το ρυθμό που γίνεται η υδρόλυση του GTP σε GDP. Μια επιπλέον λειτουργία των RGS είναι ότι διαθέτουν την ικανότητα να δράσουν και σαν ένα είδος co-GEF παράγοντα, αφού μπορούν να αυξήσουν το ρυθμό ανταλλαγής GTP-GDP στους GPCRs.

44
Ρύθμιση των GPCRs (Ι) Oι GPCRs απευαισθητοποιούνται όταν εκτίθενται στο συνδέτη τους για μεγάλη χρονική διάρκεια. Υπάρχουν δύο γνωστές μορφές απευαισθητοποίησης: Η ομόλογη, όπου μειώνεται η ικανότητα ανταπόκρισης του ενεργοποιημένου υποδοχέα σε εξωκυττάρια σήματα (GPCR is downregulated). Η ετερόλογη, όπου ένας ενεργοποιημένος υποδοχέας μειώνει αυτήν την ικανότητα ανταπόκρισης ενός άλλου υποδοχέα. ***Κλειδί αυτών είναι η φωσφορυλίωση της ενδοκυττάριας περιοχής του υποδοχέα από πρωτεϊνικές κινάσες.***
. Η ετερόλογη, όπου ένας ενεργοποιημένος υποδοχέας μειώνει αυτήν την ικανότητα ανταπόκρισης ενός άλλου υποδοχέα. ***Κλειδί αυτών είναι η φωσφορυλίωση της ενδοκυττάριας περιοχής του υποδοχέα από πρωτεϊνικές κινάσες.***")
45
Ρύθμιση των GPCRs (ΙI) Φωσφορυλίωση από την Α-κινάση ή ΡΚΑ
Σύμφωνα με έναν μηχανισμό ανάδρασης η Α-κινάση έχει τη δυνατότητα να φωσφορυλιώνει τους υποδοχείς προκαλώντας μ’αυτόν τον τρόπο ομόλογη ,σε κάποιες περιπτώσεις και ετερόλογη, απευαισθητοποίηση του υποδοχέα.

46
Ρύθμιση των GPCRs (ΙII)
Φωσφορυλίωση από τις GRKs GRKs (G-protein-coupled Receptor Kinases) Φωσφορυλιώνουν μόνο ενεργούς υποδοχείς. Είναι ρυθμιστές κλειδιά για τη σηματοδότηση. Επτά είδη τέτοιων κινασών. Η ενεργότητά τους ρυθμίζεται από τις διάφορες αλληλεπιδράσεις τους.
 Φωσφορυλιώνουν μόνο ενεργούς υποδοχείς. Είναι ρυθμιστές κλειδιά για τη σηματοδότηση. Επτά είδη τέτοιων κινασών. Η ενεργότητά τους ρυθμίζεται από τις διάφορες αλληλεπιδράσεις τους.")
47
Ρύθμιση των GPCRs (ΙV) Συνέπειες φωσφορυλίωσης GPCRs:
Η Μετατόπιση: Ο υποδοχέας φέρεται στο εσωτερικό του κυττάρου όπου αποφωσφορυλιώνεται και στη συνέχεια επιστρέφει στην κυτταρική μεμβράνη. Εναλλακτικά, ο υποδοχέας μπορεί να υποβληθεί σε λυσοζυμική αποδόμηση ή να παραμείνει στο εσωτερικό. Η σύνδεση αρρεστινών: Ο φωσφορυλιωμένος υποδοχέας μπορεί να συνδεθεί με μόρια αρρεστίνης που τον θέτουν προσωρινά εκτός λειτουργίας.

48
Ρύθμιση των GPCRs (V) Άλλοι μηχανισμοί ρύθμισης
 Άλλοι μηχανισμοί ρύθμισης")
49
Βιβλιογραφία Βασικές Αρχές Κυτταρικής Βιολογίας , Alberts, Bray, Hopkin, Johnson, Lewis, Raff, Roberts, Walter, Kεφ.16 , ‘Κυτταρική Επικοινωνία’ , σελ.: Το Κύτταρο, Μια Μοριακή Προσέγγιση , Geoffrey M. Cooper, Robert E. Hausman, Τόμος ΙΙ, Κεφ.15 , ‘Κυτταρική Σηματοδότηση’ , σελ.: Gilman AG (1987). "G proteins: transducers of receptor-generated signals". Annu. Rev. Biochem. 56: 615–49. Millar RP, Newton CL (January 2010). "The year in G protein-coupled receptor research". Mol. Endocrinol. 24 (1): 261–74.
. G proteins: transducers of receptor-generated signals . Annu. Rev. Biochem. 56: 615–49. Millar RP, Newton CL (January 2010). The year in G protein-coupled receptor research . Mol. Endocrinol. 24 (1): 261–74.")
50
Σας ευχαριστούμε για την προσοχή σας
ΤΕΛΟΣ

Παρόμοιες παρουσιάσεις













 Δομή τύπου LIFO: Last In - First Out (τελευταία εισαγωγή – πρώτη εξαγωγή) Περιορισμένος.>")





![1 Νέα Θεωρία Μεγέθυνσης Ενδογενής μεγέθυνση. 2 Συνάρτηση παραγωγής προϊόντος Υ t = Y(K, L, A) Y t = [(1-α k )·K t ] α · [(1-α L )·A t ·L t ] 1-α 0](/11/2949896/big_thumb.jpg "1 Νέα Θεωρία Μεγέθυνσης Ενδογενής μεγέθυνση. 2 Συνάρτηση παραγωγής προϊόντος Υ t = Y(K, L, A) Y t = [(1-α k )·K t ] α · [(1-α L )·A t ·L t ] 1-α 0>")
