Κατέβασμα παρουσίασης
1
水稻根负向光性的研究 学生:钱善勤 导师:王 忠 专业:植物学

2
光是对植物调控作用最广泛、最明显的环境因子。光作为环境信号,对植物的器官发生、形态建成、向性运动等方面都有深刻的影响。
植物光受体 1) 向光素 2) 隐花色素 3) 光敏色素 光质→光受体→信号转导组分→光调节基因→光形态建成反应
 向光素. 2) 隐花色素. 3) 光敏色素. 光质→光受体→信号转导组分→光调节基因→光形态建成反应.")
3
根的负向光性早在一个多世纪之前就有报导,但它对植物生长的意义还未见报导,Okada和Shimura在对拟南芥研究后发现其根系具有负向光性。我们在透明培养缸中培养水稻的根系,发现水稻种子根、次生根及其分枝根都具有明显的负向光性。同时我们也发现稗草等植物的根也有负向光性。 在此基础上,我们以水稻为主要材料,研究了负向光性的机理及其影响因素

4
1.材料与方法 1.1 供试材料 供试的水稻(Oryza sativa.L)有扬稻6号(籼稻),粳稻941(粳稻)与“三粒寸”(籼型大粒),“大力”(粳型大粒)。 前两个品种用来研究水稻次生根的负向光性,而后面两个品种则是用来研究种子根的负向光性。
有扬稻6号(籼稻),粳稻941(粳稻)与 三粒寸 (籼型大粒), 大力 (粳型大粒)。 前两个品种用来研究水稻次生根的负向光性,而后面两个品种则是用来研究种子根的负向光性。")
5
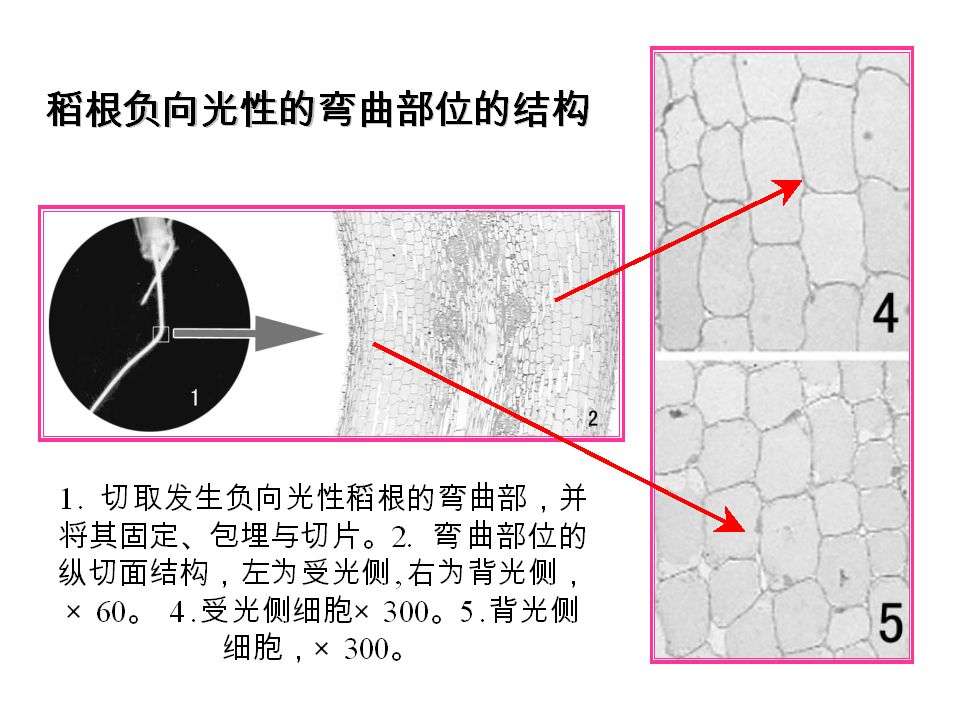
1.2实验处理的设置 (1)用锡箔包裹法及切除法研究光信号的感受部位 (2)将培养缸放到有透光玻璃的培养箱中研究温度对水稻负向光性的影响
(3)改变光源与稻根间的距离调节光强,在光源与稻根之间插入滤光片来改变光质,研究光强和光质对水稻根负向光性的影响 (4)改变培养缸中溶液成分研究化学试剂对根生长影响 (5)将稻根负向光性弯曲部位制成树脂超薄切片,并在光镜下观察根负向光性弯曲部位的显微结构 (6)采用ELISA法测定水稻根尖的生长素含量
改变光源与稻根间的距离调节光强,在光源与稻根之间插入滤光片来改变光质,研究光强和光质对水稻根负向光性的影响. (4)改变培养缸中溶液成分研究化学试剂对根生长影响. (5)将稻根负向光性弯曲部位制成树脂超薄切片,并在光镜下观察根负向光性弯曲部位的显微结构. (6)采用ELISA法测定水稻根尖的生长素含量.")
6
2 水稻根的负向光性 2.1 水稻次生根的负向光性反应
1 照光 0 h 2 照光 6 h 3 照光12 h 4 照光24 h 5 照光36 h 6 照光48 h

7
2.2 种子根的负向光性反应 间歇光照(12h) 根据水稻根在断光时垂直生长,侧光照射时背光斜向生长的特性,对生长着的水稻根给予定时的(如12h)照光和停止照光,则可看到根的生长形状呈现出锯齿状来 连续侧光照射

9
发生倾斜生长的部位是根尖 感受光的部位是根冠 对根尖遮光或切除根尖时,则不发生负向光性反应。
剥除根冠而保留根尖分生区和伸长区的根会失去背光性反应。 1.根冠的结构 2.剥除不同量根冠的稻根根尖 A. 彻底剥除根冠;B.未彻底剥除根冠;C. 对照,未剥除根冠 3.剥除根冠对水稻根尖负向性的影响

10
光质对稻根背光性的影响 用相同光强而不同光质的光照射水稻根,发现在400~720nm波段的试验范围内,蓝紫光(400nm~480nm) 对诱导根背光生长最有效,黄绿光(520nm)对根的背光生长也有效,而橙光和红光(600nm~720nm) 对诱导根背光生长无效,从而推测诱导根背光生长的光受体不是光敏素,可能是蓝光受体。 光质对稻根负向光性的影响
 对诱导根背光生长最有效,黄绿光(520nm)对根的背光生长也有效,而橙光和红光(600nm~720nm) 对诱导根背光生长无效,从而推测诱导根背光生长的光受体不是光敏素,可能是蓝光受体。 光质对稻根负向光性的影响.")
11
IAA,CTK,TIBA对扬稻6号负向光性的影响

12
IAA,CTK,TIBA对粳稻941负向光性的影响

13
EDTA对水稻根负向光性的影响

14
CaCl2对水稻根负向光性的影响

15
光照对水稻根尖生长素分布的影响 光强(100 μmol·m-2·s-1)
采用酶联免疫吸附分析(ELISA)方法测定稻根根尖生长素的含量,发现照光会引起根尖生长素发生不均等的分布,照光侧生长素含量下降,而背光侧生长素含量上升. 光强(100 μmol·m-2·s-1)
方法测定稻根根尖生长素的含量,发现照光会引起根尖生长素发生不均等的分布,照光侧生长素含量下降,而背光侧生长素含量上升. 光强(100 μmol·m-2·s-1)")
16
将稻根悬置于营养液气雾中。根尖一侧贴附含生长素的琼脂块,发现根会向贴附含生长素一侧弯曲生长
含5mg·L-1生长素的琼脂

17
3.讨论 植物的向性运动一般包括三个基本步骤: 1刺激感受, 即植物体中感受体接受环境中单方向的刺激;
2信号转导, 感受细胞把环境刺激转化成物理或化学信号; 3运动反应,生长器官接受信号后,发生不均等生长,表现出向性运动。 水稻根尖的负向光性也是光信号传递过程,应该包括这三个步骤。

18
前人的研究成果 Went (1928)用生物测定法显示生长素活性的分布比率为向光面32%,背光面68%。
Cholodny和Went以燕麦胚芽鞘为材料研究发现,在单侧蓝光作用下,IAA向胚芽鞘的背光侧移动。根据这些结果Cholodny和Went认为,植物的向光性反应是由于光照后引起生长素浓度的差异分布而引起的。 Iino以玉米为材料,对玉米胚芽鞘向光侧和背光侧的内源生长素含量变化进行了分析,证实了单侧蓝光引起生长素分布不均匀,从而使胚芽鞘产生向光弯曲。 Young等人1990年报道IAA对向重性具有重要的作用,在玉米幼根中的生长素由生长点先输入根冠。若将玉米幼根平放,在重力作用下生长素最先在根冠内不对称分布,根冠生长素浓度下侧大于上侧,不对称分布的生长素再由根冠向后运输到生长部位,引起不均等生长而导致向重性发生。

19
本试验的结果表明,稻根发生负向光性的光感受部位是根冠,根冠中接受光信号的受体可能是蓝光受体,而且生长素是水稻根负向光性反应中重要的因子。
根冠是光感受部位的实验依据是:(1) 对根冠遮光而对根尖其它部分照光时根不表现出负向光性;(2) 剥除根冠而保留根尖其它部分时根会失去负向光性;(3) 剥除根冠而保留根冠的原始细胞时稻根在新根冠长出时会恢复负向光性。Takahashi等人也曾指出植物顶端(茎尖或根尖) 是接受光后能向最适宜方向生长的部位。
 对根冠遮光而对根尖其它部分照光时根不表现出负向光性;(2) 剥除根冠而保留根尖其它部分时根会失去负向光性;(3) 剥除根冠而保留根冠的原始细胞时稻根在新根冠长出时会恢复负向光性。Takahashi等人也曾指出植物顶端(茎尖或根尖) 是接受光后能向最适宜方向生长的部位。")
20
光受体信号为蓝光/UV-A受体的依据是: 根的生长和负向光性反应与红光或远红光无关,而显著地受蓝紫光诱导.
前人的研究己表明引起植物茎的向光性弯曲的光为蓝光和紫外光,它们的光受体为蓝光/UV-A 受体,蓝光/UV-A受体的生色团的吸收区在近紫外和蓝紫光区,有的生色团也吸收绿光。现在已有研究证明向光性反应的主要光受体是向光素。

21
而生长素在水稻根负向光性反应的重要作用的则可以从试验中所采用的IAA,CTK,TIBA三种溶液的处理,可以初步推断出生长素在水稻根负向光性反应中的作用。此外在我们对水稻根采用ELISA 测定的结果也提供了有力的证据. 我们采用ELISA法测定水稻根向光侧与背光侧的生长素含量,结果显示,照光前两侧的生长素含量相当,而照光后,背光侧生长素含量比向光侧高出2-4倍,但两侧生长素总量则基本上保持不变。 通过我们的实验结果可知,水稻根在单侧光照的刺激下,细胞中的生长素从受光侧向背光侧运输,造成两侧的生长素差异,而使得根系负向光性弯曲。

22
生长素在细胞间运输是直接由生长素载体承担的。研究表明生长素的胞间运输是由细胞中生长素输出载体和输入载体的分布决定的。在拟南芥中已经鉴定一种载体蛋白PIN1。
通过细胞免疫化学的方法可研究PIN1蛋白在细胞中的分布和运动。用抑制剂布雷菲德菌素A(BFA)处理,不仅阻碍了PIN1蛋白的运动,破坏了它们在质膜上的不对称分布,并使得它们在细胞内膜上积累起来。当将BFA去除后,PIN1蛋白又会迅速恢复其正常分布。同样,BFA处理抑制了组织培养细胞中生长素的输出,也降低了幼苗的伸长作用及其向重性弯曲运动。这些结果表明了质膜上PIN1蛋白的分布情况会影响生长素的输出。
处理,不仅阻碍了PIN1蛋白的运动,破坏了它们在质膜上的不对称分布,并使得它们在细胞内膜上积累起来。当将BFA去除后,PIN1蛋白又会迅速恢复其正常分布。同样,BFA处理抑制了组织培养细胞中生长素的输出,也降低了幼苗的伸长作用及其向重性弯曲运动。这些结果表明了质膜上PIN1蛋白的分布情况会影响生长素的输出。")
23
产生负向光性产生的可能机理 A.根冠中的光受体接受光信号诱发光信号转导。
B.引起根冠和分生区之间的IAA重新分配,使向光侧IAA浓度低于背光侧(另一种可能是光照后使向光侧IAA大量氧化学系分解)。 C.IAA浓度高时抑制根细胞的伸长,浓度低时促进根 细胞的伸长。从而引起向光侧细胞长度大于背光侧细胞长度。
。 C.IAA浓度高时抑制根细胞的伸长,浓度低时促进根 细胞的伸长。从而引起向光侧细胞长度大于背光侧细胞长度。")
24
我们猜测向光性产生的机制有可能是水稻根冠细胞细胞膜上的光受体接受光信号后,进一步激发下游的信号转导,通过调控生长素输出载体的分布使生长素产生极性运输,从而使生长素浓度在向光侧和背光侧产生差异,由于生长素对细胞生长的调控作用,使向光侧和背光侧的细胞发生不均等生长而产生向光性或负向光性运动。当然这其中也不排除在光照条件下生长素和生长抑制物共同作用来调控植物的向光性反应。

25
谢谢!





 03088007 陈夏琴.>")
 班 周岑. Question 许多弱酸或弱碱,当 CaKa 或 CbKb 小于 10 -8 时,就不能用酸碱滴定法直接滴定 解决 : 增强酸碱强度 方法 : 可采用非水溶剂(包括有机溶剂或不 含水的无机溶剂)>")

 2 、研究串联谐振现象及电路参数对谐振特性 的影响。>")





 §6-2 集成模数转换器(ADC) §6-3 应用举例>")






