Κατέβασμα παρουσίασης
1
Η βιολογία της μάθησης και της μνήμης:
Μακρόχρονη ενδυνάμωση/αποδυνάμωση Λίλα Τσάλτα & Δημήτρης Κόντης Ψυχίατρος Ιατρείο Μελέτης Γνωστικής Λειτουργίας στην Ψύχωση Ψυχιατρικό Νοσοκομείο Αττικής

2
Τι λέμε μνήμη? Η ικανότητα να καταγράφουμε, να αποθηκεύουμε και να ανακαλούμε πληροφορίες Είναι ένας πολύπλοκος συνδυασμός υποσυστημάτων μνήμης

3
Κριτήρια κατηγοριοποίησης και είδη μνήμης 1
Κριτήριο Είδη μνήμης Διάρκεια αποθήκευσης πληροφορίας (Baddeley & Hitch 1974) Αισθητική (< ¼ sec)-cinema! Βραχύχρονη ή μνήμη εργασίας (secs) -γράψε το νούμερο τηλεφώνου! Μακρόχρονη (λεπτά-χρόνια) Είδος πληροφορίας (Tulving 1972) Σημασιολογική (semantic) -Ισημερινός/Κίτο Επεισοδική-αυτοβιογραφική (episodic) -τι κάνατε το Πάσχα? Διαδικαστική (procedural)-ποδήλατο Υλικό πληροφορίας Λεκτική Οπτική (ακουστική, οσφρητική, απτική, γευστική)
 Αισθητική (< ¼ sec)-cinema! Βραχύχρονη ή μνήμη εργασίας (secs) -γράψε το νούμερο τηλεφώνου! Μακρόχρονη (λεπτά-χρόνια) Είδος πληροφορίας. (Tulving 1972) Σημασιολογική (semantic) -Ισημερινός/Κίτο. Επεισοδική-αυτοβιογραφική (episodic) -τι κάνατε το Πάσχα Διαδικαστική (procedural)-ποδήλατο. Υλικό πληροφορίας. Λεκτική. Οπτική. (ακουστική, οσφρητική, απτική, γευστική)")
4
Κριτήρια κατηγοριοποίησης και είδη μνήμης 2
Κριτήριο Είδη μνήμης Αναγκαιότητα-ικανότητα συνειδητής ανάκλησης Δηλωτική-έκδηλη (explicit) Άδηλη (implicit) Αφορά παρελθόν ή μέλλον? Αναδρομική (retrospective) Προοπτική (prospective)-να πληρώσεις το λογαριασμό! Αμνησία γεγονότων πριν ή μετά τη βλάβη Οπισθόδρομη αμνησία (retrograde) Προχωρητική (anterograde) Wilson 2009
 Άδηλη (implicit) Αφορά παρελθόν ή μέλλον Αναδρομική (retrospective) Προοπτική (prospective)-να πληρώσεις το λογαριασμό! Αμνησία γεγονότων πριν ή μετά τη βλάβη. Οπισθόδρομη αμνησία (retrograde) Προχωρητική (anterograde) Wilson")
6
Ταξινόμηση μνήμης ΜΝΗΜΗ ΔΗΛΩΤΙΚΗ ΑΔΗΛΗ Αντανακλα-στικές οδοί ΓΕΓΟΝΟΤΑ
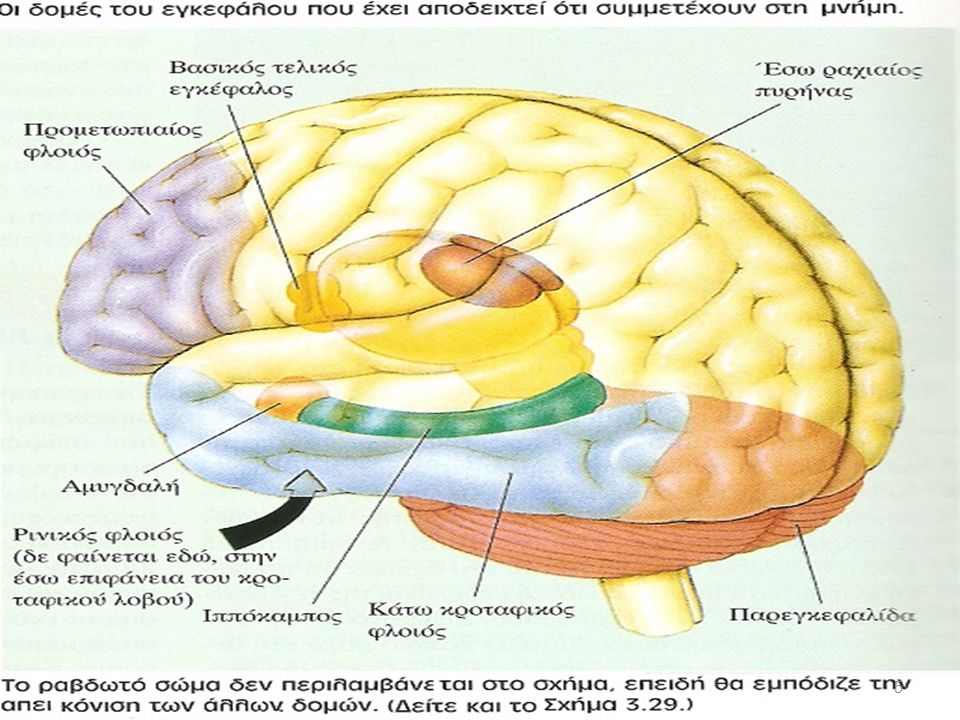
PRIMING: διευκόλυνση της ικανότητας ανίχνευσης ή ταυτοποίησης ερεθισμάτων εξαιτίας της προηγούμενης εμπειρίας με αυτά τα ερεθίσματα Ταξινόμηση μνήμης ΜΝΗΜΗ ΔΗΛΩΤΙΚΗ ΑΔΗΛΗ Αντανακλα-στικές οδοί ΓΕΓΟΝΟΤΑ (FACTS) ΔΡΩΜΕΝΑ (EVENTS) ΔΕΞΙΟΤΗΤΕΣ ΣΥΝΗΘΕΙΕΣ ΕΞΟΙΚΕΙΩΣΗ ΕΥΑΙΣΘΗΤΟΠΟΙΗΣΗ ΣΥΝΤΕΛΕΣΤΙΚΗ ΜΑΘΗΣΗ PRIMING Έσω κροταφικός λοβός Συναισθηματικές απαντήσεις Σκελετικοί μύες Ραβδωτό Νεοφλοιός Αμυγδαλή Παρεγκεφαλίδα Kandel 2000
 ΔΡΩΜΕΝΑ. (EVENTS) ΔΕΞΙΟΤΗΤΕΣ ΣΥΝΗΘΕΙΕΣ. ΕΞΟΙΚΕΙΩΣΗ ΕΥΑΙΣΘΗΤΟΠΟΙΗΣΗ. ΣΥΝΤΕΛΕΣΤΙΚΗ ΜΑΘΗΣΗ. PRIMING. Έσω κροταφικός λοβός. Συναισθηματικές απαντήσεις. Σκελετικοί μύες. Ραβδωτό. Νεοφλοιός. Αμυγδαλή. Παρεγκεφαλίδα. Kandel")
7
Nόμος του Hebb (1949) Neurons that fire together, wire together! Όταν ένας νευρώνας Α είναι αρκετά κοντά για να διεγείρει έναν νευρώνα Β και συμμετέχει κατ’ επανάληψη στην ενεργοποίηση του Β, η συναπτική σύνδεση μεταξύ των δύο νευρώνων ενισχύεται και ο νευρώνας Β γίνεται πιο ευαίσθητος σε ερεθίσματα από το νευρώνα Α

8
Νευροβιολογία Μνήμης: Timing και σύναψη
Με βάση το νόμο του Hebb, δύο ζητήματα που μας απασχολούν στη νευροβιολογία της μνήμης είναι: Το timing Η σύναψη

9
Donald Hebb Neurons that fire together, wire together!
Αλλαγές διαρκείας στην αποτελεσµατικότητα της συναπτικής µεταβίβασης αποτελούν τη βάση της µακροπρόθεσµης µνήµης

10
Συναπτικοί μηχανισμοί μάθησης και μνήμης

11
Συναπτικοί μηχανισμοί μάθησης και μνήμης
Τι συμβαίνει μέσα στις εγκεφαλικές δομές που εμπλέκονται στη μνήμη? Hebb –οι αλλαγές στη συναπτική αποτελεσματικότητα είναι η βάση της μακρόχρονης μνήμης Μακρόχρονη ενδυνάμωση (LTP) – οι συνάψεις γίνονται δυνατότερες λόγω του επαναλαμβανόμενου ερεθισμού
 – οι συνάψεις γίνονται δυνατότερες λόγω του επαναλαμβανόμενου ερεθισμού.")
12
Μακρόχρονη Ενδυνάμωση Long-Term Potentiation (LTP)
Bliss και Lomo (1973): υπάρχει µια ενίσχυση της συναπτικής διαβίβασης µετά από υψίσυχνο ηλεκτρικό ερεθισµό προσυναπτικών νευρώνων: μακρόχρονη ενδυνάμωση
: υπάρχει µια ενίσχυση της συναπτικής διαβίβασης µετά από υψίσυχνο ηλεκτρικό ερεθισµό προσυναπτικών νευρώνων: μακρόχρονη ενδυνάμωση.")
13
LTP and LTD (μακρόχρονη αποδυνάμωση) in vitro έναντι in vivo
Καλλιέργεια κυττάρων Περιορισμοί – μιμούνται ένα ακέραιο σύστημα απομάκρυνση φυσιολογικών εισερχόμενων ερεθισμάτων και περιβάλλοντος προσθήκη αποκλειστών όπως η πικροτοξίνη ή τετροδοτοξίνη έλλειψη φυσιολογικών εξερχόμενων ερεθισμάτων Πλεονεκτήματα – απάντηση καθαρή και ερμηνεύσιμη Μονήρες EPSP (excitatory post-synaptic potential/ IPSP inhibitory PSP) που είναι αναμφισβήτητο– υπάρχει ή δεν υπάρχει Όχι «μόλυνση» από άλλα εισερχόμενα ερεθίσματα In vivo: Ακέραιο αναισθητοποιημένο ή ελεύθερα κινούμενο ζώο Δεν ξέρουμε τον «αποτελεσματικό» ερεθισμό Μπορεί να μελετήσει το αποτέλεσμα ερεθισμών στη συμπεριφορά
 που είναι αναμφισβήτητο– υπάρχει ή δεν υπάρχει. Όχι «μόλυνση» από άλλα εισερχόμενα ερεθίσματα. In vivo: Ακέραιο αναισθητοποιημένο ή ελεύθερα κινούμενο ζώο. Δεν ξέρουμε τον «αποτελεσματικό» ερεθισμό. Μπορεί να μελετήσει το αποτέλεσμα ερεθισμών στη συμπεριφορά.")
14
Πού στον εγκέφαλο μελετάται η LTP?

15
Κύρια συστατικά του ιπποκάμπου

16
Πού στον εγκέφαλο μελετάται η LTP?
Διατιτραίνουσα οδός Οδοντωτή έλικα

17
LTP στην οδοντωτή έλικα του ιπποκάμπου
μακρόχρονη

18
LTP στην CA1 Καταγραφή EPSP Μέγεθος EPSP σε απάντηση εισόδου 1 Τέτανος μόνο στην είσοδο 1 Είσοδος 1 Είσοδος 2 Μέγεθος EPSP σε απάντηση ερεθίσματος 2 Requires only a brief tetanus, is input specific, and can last many weeks

19
Μακρόχρονη Ενδυνάμωση Long-Term Potentiation (LTP)
Η LTP είναι συμβατή με τις συναπτικές αλλαγές που υπέθεσε ο Hebb Μπορεί να διαρκέσει εβδομάδες Συμβαίνει μόνο όταν η προσυναπτική διέγερση ακολουθείται από μετασυναπτική διέγερση Η υπόθεση του Hebb για τη μάθηση Η συνύπαρξη διεγέρσεων σε προ- και μετα-συναπτικούς νευρώνες είναι προϋπόθεση της μάθησης και της μνήμης

20
LTP: ένας νευρωνικός μηχανισμός Μάθησης & Μνήμης
Προκαλείται από ηλεκτρικό ερεθισμό υψηλής συχνότητας ενός προσυναπτικού νευρώνα που μιμείται τη φυσιολογική νευρωνική δραστηριότητα Τα αποτελέσματά της είναι μεγαλύτερα στις περιοχές που εμπλέκονται στη μνήμη και τη μάθηση Η μάθηση μπορεί να προκαλέσει αλλαγές που μοιάζουν με την LTP Φάρμακα που επιδρούν στη μάθηση έχουν παράλληλα αποτελέσματα στην LTP

21
LTP: ένας νευρωνικός μηχανισμός Μάθησης & Μνήμης
Επαγωγή (μάθηση) Διατήρηση (μνήμη) Έκφραση (μνημονική ανάκληση)
 Διατήρηση (μνήμη) Έκφραση (μνημονική ανάκληση)")
22
Επαγωγή LTP: Μάθηση Έχει μελετηθεί κυριότερα
Συνδέονται κυρίως οι γλουταμινεργικοί NMDA υποδοχείς Οι NMDA υποδοχείς δεν δίνουν μέγιστες απαντήσεις, παρά μόνον όταν το γλουταμικό συνδέεται και ο νευρώνας είναι ήδη εκπολωμένος Τα κανάλια Ca2+ δεν ανοίγουν εντελώς, παρά μόνον όταν υφίστανται και οι δύο συνθήκες

23
Επαγωγή LTP: Μάθηση Η είσοδος του Ca2+ συμβαίνει μόνον όταν συνυπάρχουν η πρόσδεση του γλουταμικού σε έναν NMDA υποδοχέα που είναι ήδη εκπολωμένος Η είσοδος Ca2+ μπορεί να ενεργοποιήσει πρωτεϊνικές κινάσες που επάγουν αλλαγές που προκαλούν LTP

24
Υποδοχέας NMDA ΠΡΟΣΥΝΑΠΤΙΚΟΣ ΝΕΥΡΩΝΑΣ Γλουταμικό Γλυκίνη
Σύνδεση γλουταμικού+γλυκίνης + εκπόλωση μετασυναπτικού κυττάρου (απομάκρυνση μανγνησίου) : ενεργοποίηση NMDA υποδοχέα, είσοδος Ca στο κανάλι Υποδοχέας NMDA ΠΡΟΣΥΝΑΠΤΙΚΟΣ ΝΕΥΡΩΝΑΣ Γλουταμικό Γλυκίνη Eκπόλωση μετασυναπτικού κυττάρου: απομάκρυνση μαγνησίου Θέση γλουταμικού Ca Θέση μαγνησίου (στο ιοντικό κανάλι) Θέση PCP (στο ιοντικό κανάλι) Θέση πολυαμίνης ΜΕΤΑΣΥΝΑΠΤΙΚΟΣ ΝΕΥΡΩΝΑΣ Θέση γλυκίνης Θέση ψευδαργύρου
 : ενεργοποίηση NMDA υποδοχέα, είσοδος Ca στο κανάλι. Υποδοχέας NMDA. ΠΡΟΣΥΝΑΠΤΙΚΟΣ ΝΕΥΡΩΝΑΣ. Γλουταμικό. Γλυκίνη. Eκπόλωση μετασυναπτικού κυττάρου: απομάκρυνση μαγνησίου. Θέση γλουταμικού. Ca. Θέση μαγνησίου (στο ιοντικό κανάλι) Θέση PCP (στο ιοντικό κανάλι) Θέση πολυαμίνης. ΜΕΤΑΣΥΝΑΠΤΙΚΟΣ ΝΕΥΡΩΝΑΣ. Θέση γλυκίνης. Θέση ψευδαργύρου.")
25
Μονήρες δυναμικό ενέργειας
Glu AMPA a-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate εκπόλωση Glu NMDA Δεν ανοίγει Το Mg++ αποκλείει το κανάλι Λίγο Ca++ στο μετασυναπτικό κύτταρο Ακολουθείται από περισσότερα δυναμικά ενέργειας

26
Ca++ Na+ G G G G AMPA NMDA Mg

27
Ca++ Na+ G G G G AMPA NMDA Mg

28
Mg Ca++ Na+ G G G G AMPA NMDA

29
Mg Ca++ Na+ G G G G AMPA NMDA

30
Ενεργοποίηση NMDA υποδοχέα
Κανάλι Ca++ Ανοιγοκλείνει χημικά Ανοιγοκλείνει και με το δυναμικό Το Mg++ το αποκλείει εισόδου Ca++ μετασυναπτικές αλλαγές Ενδυνάμωση σύναψης: μακρόχρονη ενδυνάμωση

31
Διατήρηση και έκφραση της LTP: Αποθήκευση και Ανάκληση
Προ- και μετα-συναπτικές αλλαγές Η LTP παρατηρείται μόνο σε συνάψεις στις οποίες επάχθηκε Οι μακρόχρονες αλλαγές συνδέονται με δομικές αλλαγές (σύνθεση πρωτεϊνών) Η LTP αρχίζει στον μετασυναπτικό νευρώνα τον οποίο διεγείρει ο προσυναπτικός νευρώνας Τα αστροκύτταρα και όχι μόνο οι νευρώνες εμπλέκονται στην LTP
 Η LTP αρχίζει στον μετασυναπτικό νευρώνα τον οποίο διεγείρει ο προσυναπτικός νευρώνας. Τα αστροκύτταρα και όχι μόνο οι νευρώνες εμπλέκονται στην LTP.")
32
Αστροκύτταρα και LTP

33
Μοντέλο ρύθμισης της LTP στον ιππόκαμπο από τα αστροκύτταρα
Model of astrocyte regulation of long-term synaptic potentiation (LTP) in the hippocampus. Left panel: depiction of the experimental design used by Henneberger et al. (2010) to clamp astrocyte intracellular calcium and record dendritic field potentials at the Schaffer collateral (SC)–CA1 pyramidal cell (CA1) synapse in the hippocampus. Right panel: depiction of a model for LTP. The currently accepted model of this hippocampal synaptic LTP involves glutamate being released from the presynaptic neuron and diffusing across the synaptic cleft to activate postsynaptic receptors. N-Methyl d-aspartate receptor (NMDAR) activation is necessary for LTP induction and requires both glutamate binding and activation of the glycine-binding site. Classically, the latter is thought to be accomplished by tonic levels of glycine, although Henneberger et al. (2010) propose that d-serine, released from astrocytes, activates the glycine-binding site. In this model glutamate activates metabotropic glutamate receptors on astrocytes to increase intracellular calcium and cause d-serine release, which in turn binds the glycine-binding site of the NMDAR. By clamping internal calcium fluctuations of an astrocyte via a patch pipette, Henneberger et al. (2010) demonstrated elimination of LTP that supports this hypothesis, although another study (Agulhon et al. 2010) that uses a transgenic approach to block intracellular calcium transients finds opposite results. To more completely understand the role of astrocytes in hippocampal synaptic LTP three questions must be answered in vivo. 1) What mechanism does increased intracellular calcium act on to promote d-serine release? 2) How is d-serine released by astrocytes? 3) Is d-serine the endogenous cotransmitter involved in synaptic LTP of the hippocampus? Wenker I J Neurophysiol 2010;104: ©2010 by American Physiological Society
 in the hippocampus. Left panel: depiction of the experimental design used by Henneberger et al. (2010) to clamp astrocyte intracellular calcium and record dendritic field potentials at the Schaffer collateral (SC)–CA1 pyramidal cell (CA1) synapse in the hippocampus. Right panel: depiction of a model for LTP. The currently accepted model of this hippocampal synaptic LTP involves glutamate being released from the presynaptic neuron and diffusing across the synaptic cleft to activate postsynaptic receptors. N-Methyl d-aspartate receptor (NMDAR) activation is necessary for LTP induction and requires both glutamate binding and activation of the glycine-binding site. Classically, the latter is thought to be accomplished by tonic levels of glycine, although Henneberger et al. (2010) propose that d-serine, released from astrocytes, activates the glycine-binding site. In this model glutamate activates metabotropic glutamate receptors on astrocytes to increase intracellular calcium and cause d-serine release, which in turn binds the glycine-binding site of the NMDAR. By clamping internal calcium fluctuations of an astrocyte via a patch pipette, Henneberger et al. (2010) demonstrated elimination of LTP that supports this hypothesis, although another study (Agulhon et al. 2010) that uses a transgenic approach to block intracellular calcium transients finds opposite results. To more completely understand the role of astrocytes in hippocampal synaptic LTP three questions must be answered in vivo. 1) What mechanism does increased intracellular calcium act on to promote d-serine release 2) How is d-serine released by astrocytes 3) Is d-serine the endogenous cotransmitter involved in synaptic LTP of the hippocampus Wenker I J Neurophysiol 2010;104: ©2010 by American Physiological Society.")
34
Διατήρηση και έκφραση της LTP: Αποθήκευση και Ανάκληση
Πώς συντονίζονται οι προσυναπτικές και οι μετασυναπτικές αλλαγές? Το μονοξείδιο του αζώτου που συντίθεται στους μετασυναπτικούς νευρώνες σε απάντηση στην είσοδο Ca2+ influx μπορεί να διαχυθεί πίσω στους προσυναπτικούς νευρώνες Οι δομικές αλλαγές είναι μια τεκμηριωμένη συνέπεια της LTP

35
Αποτελέσματα εισόδου Ca2+
Ενεργοποίηση προτεϊνικών κινασών Protein Kinase C (PKC) Ca2+/calmodulin-dependent protein kinase (CaMKII) Στόχοι: υποδοχείς AMPA & άλλες πρωτεϊνες σημασιοδότησης Σημαντικός ρόλος CaMKII (Ca2+/calmodulin-dependent protein kinases) Αποκλεισμός CaMKII Όχι LTP Αυτό-φωσφορυλίωση διάρκειας LTP
 Ca2+/calmodulin-dependent protein kinase (CaMKII) Στόχοι: υποδοχείς AMPA & άλλες πρωτεϊνες σημασιοδότησης. Σημαντικός ρόλος CaMKII (Ca2+/calmodulin-dependent protein kinases) Αποκλεισμός CaMKII Όχι LTP. Αυτό-φωσφορυλίωση διάρκειας LTP.")
36
LTP: Μετασυναπτικές αλλαγές
Σύνθεση υποδοχέων Περισσότερες συνάψεις Αλλαγή δομής δενδριτικών ακάνθων Σύνθεση μονοξειδίου του αζώτου

37
Μηχανισμοί για την LTP στην CA1
1. Αυξημένη αποτελεσματικότητα των υπαρχόντων μετασυναπτικών AMPA υποδοχέων (φωσφορυλίωσή τους). Η φωσφορυλίωση του ΑΜΡΑ υποδοχέα από την PKC αυξάνει την ιοντική διαπερατότητα του καναλιού 2. Είσοδος AMPA υποδοχέων στη μεμβράνη 3. Αλλαγές στη δομή της σύναψης, νέες προσεκβολές στους μετασυναπτικούς δενδρίτες, «φυτρώνουν» άξονες και σχηματίζονται πολλές συνάψεις.
. Η φωσφορυλίωση του ΑΜΡΑ υποδοχέα από την PKC αυξάνει την ιοντική διαπερατότητα του καναλιού. 2. Είσοδος AMPA υποδοχέων στη μεμβράνη. 3. Αλλαγές στη δομή της σύναψης, νέες προσεκβολές στους μετασυναπτικούς δενδρίτες, «φυτρώνουν» άξονες και σχηματίζονται πολλές συνάψεις.")
38
Πριν την LTP Αλλαγή δομής σύναψης Προσυναπτική αξονικη απόληξη
Αλλαγή δομής σύναψης Πριν την LTP Προσυναπτική αξονικη απόληξη Δενδριτική άκανθα

39
Μετά την LTP Αλλαγή δομής σύναψης Λιγότερη Φοδρίνη (πρωτεϊνη)
Αλλαγή δομής σύναψης Μετά την LTP Προσυναπτική αξονικη απόληξη Λιγότερη Φοδρίνη (πρωτεϊνη) Μικρότερη αντίσταση Δενδριτική άκανθα
 Μικρότερη αντίσταση. Δενδριτική άκανθα.")
40
Μονοξείδιο του αζώτου- NO
Οπισθόδρομος αγγελιοφόρος ↑ δηλητηριώδες αέριο Υψηλή λιποδιαλυτότητα αποθήκευση? Σύνθεση επί ζήτησης Ca++ NO synthase NO Αυξάνει τη σύνθεση νευροδιαβιβαστών στον προσυναπτικό νευρώνα Μεγαλύτερη απελευθέρωση κατά τη διάρκεια του δυναμικού ενέργειας

41
NO cGMP Glu Ca++ G G G G NO NOS Ca++

42
Συνειρμική LTP και Μάθηση?

43
Source “B” “A” “C” Συνειρμική LTP Επαναλαμβανόμενος ερεθισμός Α και Β

44
Μεταβλητότητα της LTP Η περισσότερη έρευνα έχει εστιαστεί στην LTP στον ιππόκαμπο που μεσολαβείται από τον NMDA υποδοχέα, αλλά η LTP ρυθμίζεται από διαφορετικούς μηχανισμούς και σε άλλες περιοχές του εγκεφάλου Παρά τις ανακαλύψεις η LTP και η βιολογική βάση της μνήμης παραμένουν ένα μυστήριο

45
Μακρόχρονη αποδυνάμωση: Long term Depression (LTD)
Υπάρχει επίσης και η μακρόχρονη αποδυνάμωση (LTD -long-term depression) Mακρόχρονη μείωση στη συναπτική δύναμη
 Mακρόχρονη μείωση στη συναπτική δύναμη.")
46
Τύποι Μακρόχρονης Αποδυνάμωσης (LTD)
Linden & Connor, 1995

47
Ανατομία Παρεγκεφαλίδας
ΔΙΕΓΕΡΤΙΚΕΣ Παράλληλες ίνες Αναρριχώμενες ίνες INHIBITORY Κύτταρα Purkinje Άστεροειδή κύτταρα Καλαθιοφόρα κύτταρα

48
Μακρόχρονη Αποδυνάμωση στην Παρεγκεφαλίδα
Ερεθισμός παράλληλων ινών Καταγραφή απάντησης των κυττάρων Punkinje στον ερεθισμό των παράλληλων ινών Απάντηση κυττάρων Punkinje Χρόνος από το ζευγάρωμα αναρριχώμενων και παράλληλων ινών (min) Ερεθισμός αναρριχώμενων ινών Μετά το ζευγάρωμα αναπτύσσεται LTD της απάντησης στον ερεθισμό των παράλληλων ινών
 Ερεθισμός αναρριχώμενων ινών. Μετά το ζευγάρωμα αναπτύσσεται LTD της απάντησης. στον ερεθισμό των παράλληλων ινών.")
49
Μηχανισμός LTD στην Παρεγκεφαλίδα
X = no LTP Οι υποδοχείς AMPA εσωτερικεύονται: Μετασυναπτικό αποτέλεσμα CF ενεργοποιεί το κύτταρο Purkinje, η είσοδος Na+ εκπολώνει τον δενδρίτη, κι ενεργοποιούναι τα κανάλια Ca2+. PF ενεργοποίηση (glutamate) επίσης αυξάνει την είσοδο Na+, μέσω υποδοχέων AMPA. Το γλουταμικό επίσης ενεργοποιεί υποδοχείς mGluR. Αυτό δημιουργεί DAG που ενεργοποιεί PKC. Η PKC φωσφορυλιώνει πρωτεϊνες οδηγώντας σε μείωση του αριθμού των AMPA υποδοχέων στην προσυναπτική μεμβράνη.
 επίσης αυξάνει την είσοδο Na+, μέσω υποδοχέων AMPA. Το γλουταμικό επίσης ενεργοποιεί υποδοχείς mGluR. Αυτό δημιουργεί DAG που ενεργοποιεί PKC. Η PKC φωσφορυλιώνει πρωτεϊνες οδηγώντας σε μείωση του αριθμού των AMPA υποδοχέων στην προσυναπτική μεμβράνη.")
50
Σημασία αλλαγών στη συναπτική αποτελεσματικότητα
Η LTP (και LTD) έχουν ιδιαίτερο ενδιαφέρον γιατί η μνήμη και η μάθηση πιστεύεται ότι συνδέονται με μακρόχρονες αλλαγές στη συναπτική αποτελεσματικότητα. Έχει βρεθεί ένας αριθμός συσχετίσεων μεταξύ της χωροταξικής μάθησης σε ακέραια ζώα και LTP σε τομές ιπποκάμπου (π.χ. και οι δυο αποκλείονται από ανταγωνιστές NMDA or mGlu υποδοχέων) Η LTP στην αμυγδαλή σχετίζεται στενά με εξαρτημένο φόβο • αρουραίοι που εκπαιδεύονται να συνδέουν ένα δυσάρεστο ερέθισμα στο πόδι με έναν ήχο εμφανίζουν αυξημένο αντανακλαστικό αιφνιδιασμού • τα κύτταρα στην αμυγδαλή δείχνουν LTP αύξηση στις συναπτικές τους απαντήσεις σε ακουστικά ερεθίσματα. • και τα δυο αποκλείονται από ανταγωνιστές NMDA υποδοχέων.
 έχουν ιδιαίτερο ενδιαφέρον γιατί η μνήμη και η μάθηση πιστεύεται ότι συνδέονται με μακρόχρονες αλλαγές στη συναπτική αποτελεσματικότητα. Έχει βρεθεί ένας αριθμός συσχετίσεων μεταξύ της χωροταξικής μάθησης σε ακέραια ζώα και LTP σε τομές ιπποκάμπου (π.χ. και οι δυο αποκλείονται από ανταγωνιστές NMDA or mGlu υποδοχέων) Η LTP στην αμυγδαλή σχετίζεται στενά με εξαρτημένο φόβο. • αρουραίοι που εκπαιδεύονται να συνδέουν ένα δυσάρεστο ερέθισμα στο πόδι με έναν ήχο εμφανίζουν αυξημένο αντανακλαστικό αιφνιδιασμού. • τα κύτταρα στην αμυγδαλή δείχνουν LTP αύξηση στις συναπτικές τους απαντήσεις σε ακουστικά ερεθίσματα. • και τα δυο αποκλείονται από ανταγωνιστές NMDA υποδοχέων.")
51
Συμπεράσματα Η έρευνα της νευροβιολογίας της μνήμης και της μάθησης έχει εστιάσει στο φαινόμενο της μακρόχρονης ενδυνάμωσης στις νευρωνικές συνάψεις σε διάφορες περιοχές του εγκεφάλου, ιδίως στον ιππόκαμπο Η μακρόχρονη ενδυνάμωση ακολουθεί το νόμο του Hebb και συνοδεύεται από αλλαγές στη δομή, στον αριθμό και την αποτελεσματικότητα των συνάψεων Η μακρόχρονη ενδυνάμωση συνδέεται με τη λειτουργία των NMDA υποδοχέων του γλουταμικού

52
Συμπεράσματα Αντίστροφο φαινόμενο που επίσης συνδέεται με τη μνήμη και τη μάθηση είναι η μακρόχρονη αποδυνάμωση Παρά τις ανακαλύψεις η LTP και η νευρωνική βάση της μνήμης παραμένουν ένα μυστήριο




 : Επιδείνωση κοινωνικών δεικτών Πληθυσμός σε κίνδυνο φτώχειας ή κοινωνικό αποκλεισμό.>")
















