Κατέβασμα παρουσίασης
1
METABOLISMUL GLICOGENULUI

2
METABOLISMUL GLICOGENULUI
GLICOGENOGENEZA GLICOGENOLIZA

3
METABOLISMUL GLICOGENULUI
Sursa constanta de G= conditie esentiala a vietii G= sursa pt tesutul cerebral, eritrocite mature G= sursa pt m. striat 3 surse de G: 1. aport exogen de G si precursori glucogeni (amidon, mono-, dizaharide) inconstant 2. glicogenoliza 3.GNG asigura sinteza sustinuta de G, dar V de raspuns la hipoglicemie este redusă
 inconstant. 2. glicogenoliza. 3.GNG asigura sinteza sustinuta de G, dar V de raspuns la hipoglicemie este redusă.")
4
METABOLISMUL GLICOGENULUI
Glicogenoliza= potential rapid de mobilizare a G din glicogen la nivel hepatic şi renal In timpul efortului fizic: glicogenoliza musculara In conditiile epuizarii glicogenului: GNG din AA rezulatati din catabolizarea proteinelor

5
METABOLISMUL GLICOGENULUI
Principalele rezerve de glicogen: in muschiul striat (400g glicogen= 1-2% din masa reala M) : sinteza ATP in contractie musculara ficat (100g glicogen=10%Fi): rol in mentinerea glicemiei mici cantitati in toate celulele
 : sinteza. ATP in contractie musculara. ficat (100g glicogen=10%Fi): rol in mentinerea. glicemiei. mici cantitati. in toate celulele.")
6
STRUCTURA GLICOGENULUI

7
STRUCTURA GLICOGENULUI
Granulaţii citoplasmatice discrete ce conţin enzimele necesare sintezei şi degradării glicogenului GRANULE DE GLICOGEN

8
UDP-G-PIROFOSFORILAZA
GLICOGENOGENEZA În citosol din α-D-Glc, cu consum de ATP (fosforilarea G) si UTP A. Sinteza UDP-G UDP-G-PIROFOSFORILAZA G-1-P UDP-G
 si UTP. A. Sinteza UDP-G. UDP-G-PIROFOSFORILAZA. G-1-P. UDP-G.")
9
UDP-G-PIROFOSFORILAZA
GLICOGENOGENEZA UDP-G-PIROFOSFORILAZA UDP-G

10
GLICOGENOGENEZA Reactia de activare devine ireversibila deoarece, in vivo, se formeaza fosfat prin degradarea pirofosfatului sub actiunea pirofosfatazei: PIROFOSFATAZA

11
GLICOGENOGENEZA FOSFOGLUCOMUTAZA G-1-P G-6-P G-1-P UDP-G G-1-P UDP-G

12
GLICOGENOGENEZA Sinteza glicogenului se realizeaza prin atasarea resturilor glucozil de pe UDP-glucoza pe o molecula de glicogen “amorsa” (glicogen (n-1)). Formarea glicogenului primer o realizeaza glicogenina (o proteina cu M 37,2 kDa) care functioneaza autocatalitic. In final, UTP-ul necesar activarii se reface prin reactia:
). Formarea glicogenului primer o realizeaza glicogenina (o proteina cu M 37,2 kDa) care functioneaza autocatalitic. In final, UTP-ul necesar activarii se reface prin reactia:")
13
GLICOGENOGENEZA B. Glicogen-sintaza asigură formarea legăturilor α (1→4) alungeşte catenele de G deja formate. În absenţa fragmentului de glicogen- o proteina, glicogenina AUTOCATALIZARE
 alungeşte catenele de G deja formate. În absenţa fragmentului de glicogen- o proteina, glicogenina. AUTOCATALIZARE.")
14
GLICOGENOGENEZA Glicogen sintaza este enzima ce catalizeaza transferul unor grupe glucozil activate pe o molecula de glicogen “amorsa” (primer) formand legaturi 1,4-glicozidice putand adauga maxim 10 resturi glucozil Este o enzima care se prezinta sub doua forme interconvertibile prin fosforilare–defosforilare, glicogen sintaza D, fosforilata (inactiva) si glicogen sintaza I, nefosforilata (activa);
 formand legaturi 1,4-glicozidice putand adauga maxim 10 resturi glucozil. Este o enzima care se prezinta sub doua forme interconvertibile prin fosforilare–defosforilare, glicogen sintaza D, fosforilata (inactiva) si glicogen sintaza I, nefosforilata (activa);")
15
GLICOGENOGENEZA LANT GLICOGENIC SCURT (legaturi α 1→4)
")
16
E=NUCLEOZID-DIFOSFAT-
GLICOGENOGENEZA C. Alungirea lanturilor de glicogen: transferul G din UDP-G la capătul nereducător al lanţului în constituire, cu formarea unei noi legături glicozidice α 1→4 E=NUCLEOZID-DIFOSFAT- KINAZA UDP + ATP UTP + ADP E

17
GLICOGENOGENEZA D. Formarea ramificaţiilor în molecula de glicogen
la 8 resturi glicozil cu creşterea nr de capete nereducătoare la care se ataşează resturi de glicozil noi AMILO-α (1→4)→ (1→6)- TRANSGLUCOZIDAZA
→ (1→6)- TRANSGLUCOZIDAZA.")
18
GLICOGENOGENEZA Enzima de ramificare, (amilo α1,4 –α1,6-transglucozidaza) enzima ce scindeaza un bloc de 6-7 resturi glucozil de pe un lant in crestere si le transfera pe un alt lant realizand o legatura 1,6-glicozidica. Noua ramificatie trebuie sa se gaseasca la o distanta de cel putin 4 resturi glucozil fata de ramificatia adiacenta Ecuatia glicogenogenezei este: Glicogen(n-1) + Glucoza + 2ATP Glicogen(n) + 2ADP + 2Pi
 enzima ce scindeaza un bloc de 6-7 resturi glucozil de pe un lant in crestere si le transfera pe un alt lant realizand o legatura 1,6-glicozidica. Noua ramificatie trebuie sa se gaseasca la o distanta de cel putin 4 resturi glucozil fata de ramificatia adiacenta. Ecuatia glicogenogenezei este: Glicogen(n-1) + Glucoza + 2ATP Glicogen(n) + 2ADP + 2Pi.")
19
GLICOGENOGENEZA Sinteza ramificaţiilor adiţionale: formarea de legături α 1→4

20
GLICOGENOLIZA Catabolizarea glicogenului din rezerve hepatice şi musculare. Procesul se desfasoara in ficat si in muschiul in contractie. GLICOGEN

21
GLICOGENOLIZA GLICOGEN GLICOGEN- FOSFORILAZA GLUCOZO-1- P GLUCOZO-6- P
GLUCOZA (SÂNGE) GLICOLIZA
 GLICOLIZA.")
22
GLICOGENOLIZA Degradarea glicogenului presupune scindarea legaturilor 1,4- si 1,6-glicozidice prin actiunea conjugata a doua sisteme enzimatice distincte: a) Sistemul fosforilazic care cuprinde enzimele ce intervin in activarea fosforilazei, enzima ce scindeaza legaturile 1,4-glicozidice din glicogen prin fosforoliza, reactie ce implica transferul unui rest fosfat pe un rest glucozil, cu formare de glucozo-1-fosfat; b) Enzima de deramificare - are capacitatea de a transfera o unitate triglucidica de pe un lant pe altul formand legaturi 1,6–glicozidice si de a scinda restul de la zona de ramificatie.
 Sistemul fosforilazic care cuprinde enzimele ce intervin in activarea fosforilazei, enzima ce scindeaza legaturile 1,4-glicozidice din glicogen prin fosforoliza, reactie ce implica transferul unui rest fosfat pe un rest glucozil, cu formare de glucozo-1-fosfat; b) Enzima de deramificare - are capacitatea de a transfera o unitate triglucidica de pe un lant pe altul formand legaturi 1,6–glicozidice si de a scinda restul de la zona de ramificatie.")
23
Necesită un set de enzime specifice.
GLICOGENOLIZA Necesită un set de enzime specifice.

24
GLICOGENOLIZA

25
GLICOGENOLIZA A. Scurtarea lanţurilor de glicogen
Glicogen-fosforilaza clivează secvenţial legăturile α (1→4) dintre resturile glicozil de la capetele nereducatoare ale lanţurilor de glicogen până când se ajunge ca fiecare lanţ să conţină doar 4 unităţi glicozil înainte de orice punct de ramificaţie E conţine o moleculă de piridoxal-fosfat (coenzimă) legată covalent Structura rezultată este denumită dextrina finală şi nu mai poate fi scindată de fosforilază
 dintre resturile glicozil de la capetele nereducatoare ale lanţurilor de glicogen până când se ajunge ca fiecare lanţ să conţină doar 4 unităţi glicozil înainte de orice punct de ramificaţie. E conţine o moleculă de piridoxal-fosfat (coenzimă) legată covalent. Structura rezultată este denumită dextrina finală şi nu mai poate fi scindată de fosforilază.")
26
GLICOGENOLIZA CAPATUL NEREDUCATOR GLICOGEN cu "n" molecule de G
FOSFORILAZA GLICOGEN cu "n-1" molecule de G G-1-P

27
GLICOGENOLIZA GLICOGEN GLUCOZO-1-P

28
GLICOGENOLIZA Tezaurismoze (boli de stocare). Tipul V (Sdr McARDLE):
Deficit de glicogen-fosforilaza sau miofosforilaza in m. scheletic) afectarea m.striati; VN ale enzimei hepatice fatigabilitate musculara dupa efort absenta cresterii lactatului dupa efort sustinut dezvoltare psihica normala Mb-emie si Mb-urie afectiune cronica benigna [glicogen cu structura normala]
 afectarea m.striati; VN ale enzimei hepatice. fatigabilitate musculara dupa efort. absenta cresterii lactatului dupa efort sustinut. dezvoltare psihica normala. Mb-emie si Mb-urie. afectiune cronica benigna. [glicogen cu structura normala]")
29
GLICOGENOLIZA Fosforilaza nu desface legaturile 1,6-glicozidice, actiunea sa oprindu-se la 4 resturi glucozil fata de o ramificatie 1,6 La acest nivel actioneaza cel de-al doilea sistem enzimatic enzima de deramificare care are proprietatea de a transfera o unitate triglucidica de pe un lant pe altul In zona de ramificatie ramane un singur rest glucozil care va fi scindat de aceeasi enzima ce are si actiune 1,6-glucozidazica. Se continua apoi activitatea fosforilazei pana in apropierea unei noi ramificatii.

30
GLICOGENOLIZA B.Clivarea ramificaţiilor: sub acţiunea enzimatică dublă a unei proteine bifuncţionale- Enzima de deramificare Oligo- α (1→4)→α (1→4)- glucan-transferaza clivează 3 resturi glicozil exterioare (din cele 4) ataşate fiecarei ramificaţii Aceeaşi E transferă resturile glicozil clivate la capatul nereducator al unui lant determinand alungirea corespunzatoare a acestuia; o legătură α (1→4) este scindată şi o altă legătură α (1→4) este constituită E acţionează ca o 4: 4 TRANSFERAZA
→α (1→4)- glucan-transferaza clivează 3 resturi glicozil exterioare (din cele 4) ataşate fiecarei ramificaţii. Aceeaşi E transferă resturile glicozil clivate la capatul nereducator al unui lant determinand alungirea corespunzatoare a acestuia; o legătură α (1→4) este scindată şi o altă legătură α (1→4) este constituită. E acţionează ca o 4: 4 TRANSFERAZA.")
31
GLICOGENOLIZA GLICOGEN- FOSFORILAZA G-1-P TRANSFERAZA α (1→6)-
GLUCOZIDAZA G liberă

32
GLICOGENOLIZA α (1→6)- GLUCOZIDAZA
- GLUCOZIDAZA")
33
GLICOGENOLIZA Intre actiunea fosforilazei la nivel muscular si hepatic exista diferente notabile care sunt dependente de concentratia glucozei sanguine si de actiunea unor hormoni: adrenalina – ce determina activarea fosforilazei musculare; glucagonul – care stimuleaza activarea fosforilazei hepatice.

34
GLICOGENOLIZA În acest moment, lanţul glucidic poate fi din nou clivat sub acţiunea glicogen-fosforilazei până la detaşarea următoarelor 4 unităţi glicozil C. Conversia G-1-P la G-6-P. E= FOSFOGLUCOMUTAZA G-1-P G-1,6-BP G-6-P

35
GLICOGENOLIZA În ficat, G-6-P este translocat în RE prin intervenţa G -6-P-translocazei În RE, G-6-P este convertit la G în prezenţa G-6-fosfatazei, ce intervine în etapa finală a GNG G rezultată este transportată din RE în citosol cu GLUT-7 Hepatocitele eliberează G din glicogenoliză direct în sg circ În muşchiul striat, G-6-P este supus glicolizei şi asigură energia necesară contracţiei musculare (G-6-fosfataza absentă)
")
36
GLICOGENOLIZA Degradarea lizozomală a glicogenului (1-3%) în mod continuu sub acţiunea unei enzime lizozomale: α (1→4) glucozidaza sau maltaza acidă Deficitul E determină acumularea de glicogen sub formă de vacuole lizozomale= Boala Pompe, tipul II glicemie normală cardiomegalie importantă tratament prin substitutie enzimatica forma infantila: deces prematur prin insuficienta cardiaca glicogen cu structura normala

37
GLICOGENOLIZA Tipul Ia (Boala von Gierke):deficit de G-6-fosfatază Tipul Ib: deficit de G-6-fosfat-translocaza Ri, Fi afectate steatoza hepatica, hepato-si renomegalie retard psihosomatic si pubertate intarziate hiperlactacidemie, hiperlipidemie, hiperuricemie glicogen normal ca structura Tratament: infuzii gastrice nocturne de G sau administrarea regulata de amidon de porumb neprelucrat termic

38
GLICOGENOLIZA Deficienta ereditara a unor sisteme enzimatice implicate in metabolismul glicogenului determina aparitia bolilor ereditare denumite glicogenoze, caracterizate prin tezaurizarea in diverse tesuturi a unor cantitati mari de glicogen cu structura normala sau anormala in unele tesuturi (ficat, rinichi, intestin, muschi, creier, etc), fiind insotite de hipoglicemie, acidoza lactica. Sunt noua tipuri de glicogenoze notate I-IX, de exemplu: Maladia Von Gierke (tip I) – deficit de glucozo-6-fosfataza; Maladia Pompe (tip II) – deficit de glicozidaza lizozomala. Maladia Forbes (tip III) – deficit de enzima de deramificare. Maladia Andersen (tip IV) – deficit de enzima de ramificare.
, fiind insotite de hipoglicemie, acidoza lactica. Sunt noua tipuri de glicogenoze notate I-IX, de exemplu: Maladia Von Gierke (tip I) – deficit de glucozo-6-fosfataza; Maladia Pompe (tip II) – deficit de glicozidaza lizozomala. Maladia Forbes (tip III) – deficit de enzima de deramificare. Maladia Andersen (tip IV) – deficit de enzima de ramificare.")
39
REGLAREA SINTEZEI ŞI A CATABOLIZARII GLICOGENULUI
Cele doua procese se afla sub controlul adrenalinei, glucagonului si insulinei, astfel incat ele sa nu se produca simultan Fosforilaza reprezinta punctul principal de control al glicogenolizei, iar glicogen- sintaza al glicogenogenezei.

40
REGLAREA SINTEZEI ŞI A CATABOLIZARII GLICOGENULUI
2 niveluri: glicogen-sintaza glicogen-fosforilaza Reglare: alosterica: concentratia metabolitilor, necesarul energetic al celulei hormonala: pe calea mediata de AMPc

41
REGLAREA SINTEZEI ŞI A CATABOLIZARII GLICOGENULUI
Reglarea alosterica in aport alimentar adecvat glicogen-sintaza este activata de [G-6-P] glicogen-fosforilaza este inhibata de [G-6-P] ;ATP În contractie musculara, fosforilaz-kinaza este activata de c Ca2+- calmodulina fara interventia protein-kinazei AMPc- dependenta În relaxare musculara, fosforilaz-kinaza este inactivata Ca2+-revine in reticolul sarcoplasmic

42
REGLAREA SINTEZEI ŞI A CATABOLIZARII GLICOGENULUI
[AMPc] activeaza glicogen- fosforilaza musculara (muschi striat in anoxie extrema si depletie de ATP) AMP se leaga de forma inactiva a glicogen- fosforilazei şi induce activarea acesteia
 AMP se leaga de forma inactiva a glicogen- fosforilazei şi induce activarea acesteia.")
43
REGLAREA SINTEZEI ŞI A CATABOLIZARII GLICOGENULUI
Inhibitia glicogenogenezei prin calea metabolica mediata de AMPc Glicogen- sintaza este activa in forma defosforilata sau "a" si inactiva in forma fosforilata sau "b" Fosforilarea E este determinata de : PK C,

44
REGLAREA SINTEZEI ŞI A CATABOLIZARII GLICOGENULUI
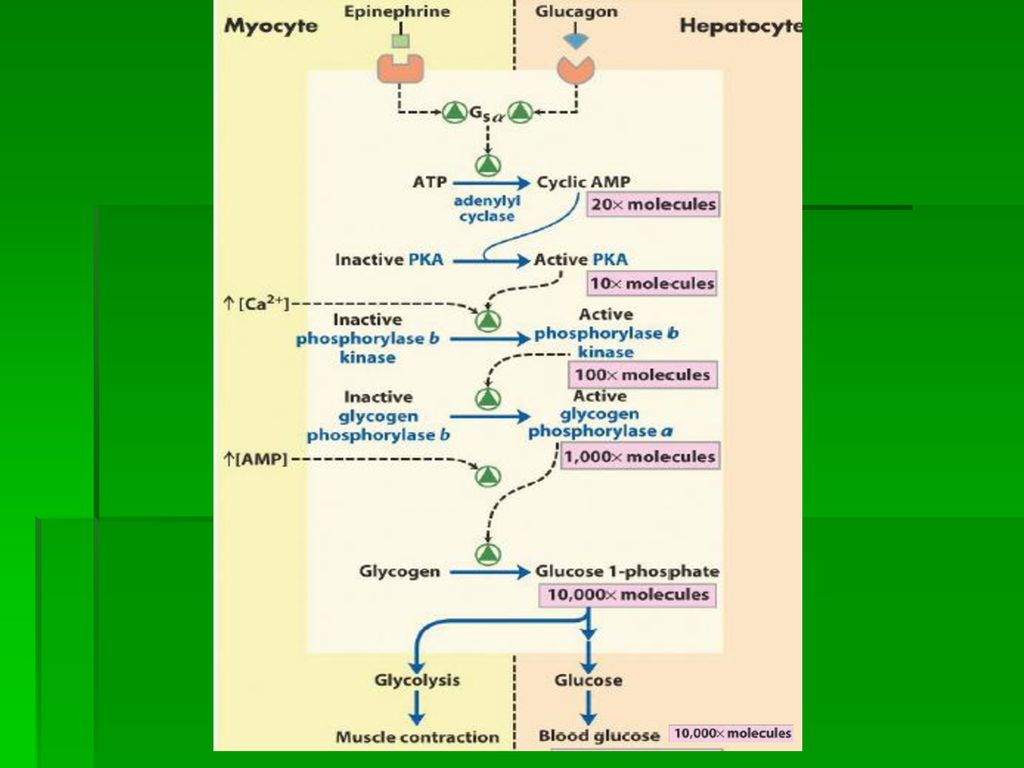
B. Activarea glicogenolizei pe calea mediata de AMPc Atasarea hormonilor (epinefrina/glucagon) la R sp membranari activeaza glicogenoliza cu cresterea glicemiei si a energiei necesare m. striat in efort Activarea protein-kinazei Activarea fosforilaz-kinazei Activarea glicogen-fosforilazei Aceasta cascada de reactii induce sinteza unui nr mare de molecule de glicogen-fosforilaza activa "a" capabila sa degradeze glicogenul
 la R sp membranari activeaza glicogenoliza cu cresterea glicemiei si a energiei necesare m. striat in efort. Activarea protein-kinazei. Activarea fosforilaz-kinazei. Activarea glicogen-fosforilazei. Aceasta cascada de reactii induce sinteza unui nr mare de molecule de glicogen-fosforilaza activa a capabila sa degradeze glicogenul.")
46
REGLAREA SINTEZEI ŞI A CATABOLIZARII GLICOGENULUI
Inhibitia glicogenezei- Glicogen-sintaza

48
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
Surse de Fr: sucroza, fructe, mierea de albine siropul de porumb concentrat Patrunderea in celule nu este insulino-dependenta, iar Fr nu stimuleaza secretia de insulina Fosforilarea Fr este catalizata de: HEXOKINAZA- in toate celulele (S= alte hexoze), Km conversie min. la Fr-6-P la [G] FRUCTOKINAZA- mecanism principal de fosforilare in Fi, Ri si intestin (aici si aldolaza B) la Fr-1-P (ATP)
, Km. conversie min. la Fr-6-P la [G] FRUCTOKINAZA- mecanism principal de fosforilare in Fi, Ri si intestin (aici si aldolaza B) la Fr-1-P (ATP)")
49
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
Fr-6-P Fructoza Fr-1-P

50
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
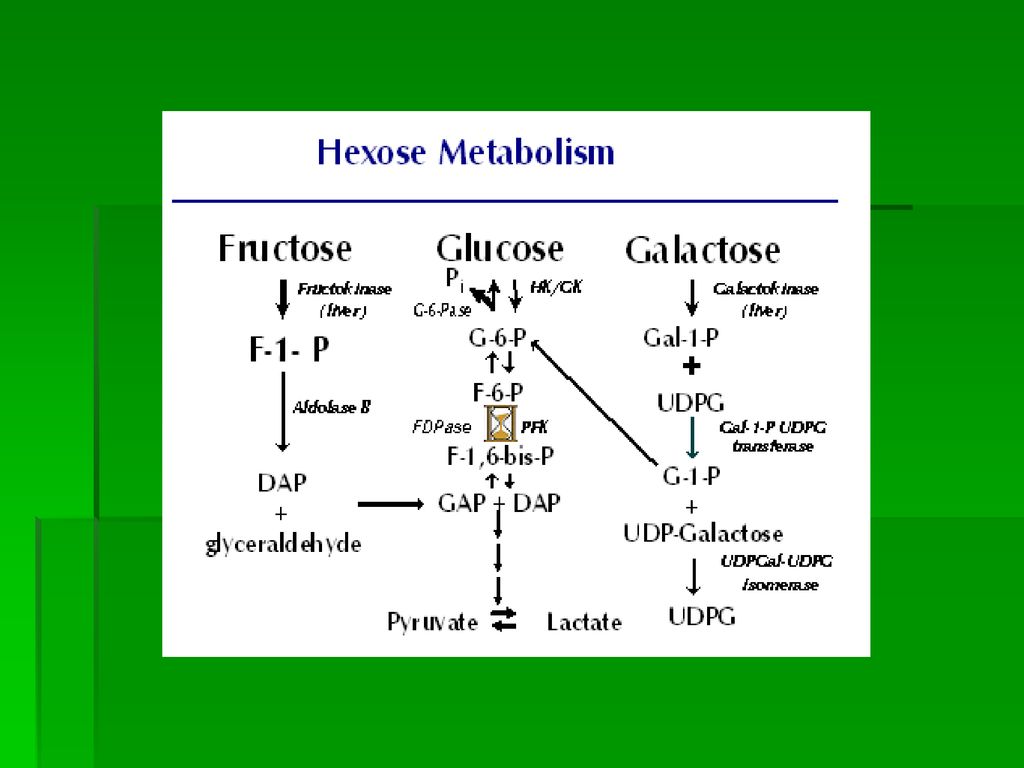
Clivarea Fr-1-P: sub acţiunea aldolazei B in DHAP si gliceraldehida aldolaza A (toate tesuturile) si aldolaza B scindeaza Fr-1,6 BP ;a DHAP si GAP numai aldolaza B scindeaza Fr-1-P DHAP intra direct in glicoliza, gliceraldehida poate fi redusa la glicerol (TAG) sau fosforilata la GAP (Py) Rata de metabolizare a Fr este mult mai mare comparativ cu G: triozele formate din Fr-1-P sunteaza PFrK, enzima limitanta de viteza a glicolizei
 si aldolaza B scindeaza Fr-1,6 BP ;a DHAP si GAP. numai aldolaza B scindeaza Fr-1-P. DHAP intra direct in glicoliza, gliceraldehida poate fi redusa la glicerol (TAG) sau fosforilata la GAP (Py) Rata de metabolizare a Fr este mult mai mare comparativ cu G: triozele formate din Fr-1-P sunteaza PFrK, enzima limitanta de viteza a glicolizei.")
51
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
FRUCTOZA-6-P FRUCTOZA DHAP ALDOLAZA B GAP

52
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
Dereglările metabolismului Fr Deficitul de fructokinaza: benign Deficitul de aldolaza B= intoleranta ereditara la Fr, IEF semnele apar dupa sistarea alaptarii Fr-1-P se acumuleaza, scade Pi si ATP, creste [AMP] In absenta Pi, AMP este catabolizat la acid uric cu hiperuricemie [ATP] hepatic determina: hipoglicemie ( - GNG), scaderea sintezei proteice cu scaderea conc plasmatice de factori de coagulare
, scaderea sintezei proteice cu scaderea conc plasmatice de factori de coagulare.")
53
INTOLERANTA EREDITARA LA FRUCTOZA

54
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
Deficitul de aldolaza B Diagnostic: Fr in urina teste genetice (RFLP ce stabileste polimorfismul electroforetic al lungimii fragmentelor de ADN rezultate prin digeastia enzimatica a ADN tinta) Tt: Fr si sucroza tb eliminate din alimentatie pt a preveni IHep si decesul
 Tt: Fr si sucroza tb eliminate din alimentatie pt a preveni IHep si decesul.")
55
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI

57
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI

58
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
[TG] crescută

59
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
HEXOKINAZA FOSFO-MANOZOIZOMERAZA Manoza- % mic din CH alimentari, epimer C-2 al G, component al GP Hexokinaza fosforileaza Man si produce Man-6-P ce va fi izomerizat rev. la Fr-6-P in prezenta fosfo-manozo-izomeraza Cea mai mare parte a Man intracel. este sintetizata din Fr sau din degradarea glucidelor structurale in prezenta hexokinazei

60
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
Conversia G in Fr via sorbitol Glucidele fosforilate nu pot traversa mb-ele biologice decat cu transportori specifici Un mecanism alternativ de metabolizare a monozaharidelor: transformarea in polioli Aldoz-reductaza este prezenta in: cristalin, retina, celule Schwann, Fi, Ri, placenta, eritocite, ovare, vezicule Sorbitol-dehidrogenaza: celule hepatice (mecanism prin care sorbitolul e transformat in intermediari ai glicolizei si GNG), ovariene, celulele liniei spermatice si ale vezicululor seminale
, ovariene, celulele liniei spermatice si ale vezicululor seminale.")
61
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
Efectul hiperglicemiei asupra metabolismului sorbitolului In celulele mentionate, [G] şi o rezerva adecvata de NADPH stimuleaza activitatea aldoz- reductazei, cu cresterea sorbitolului intracel. Sorbitolul ramane blocat in celula, fenomen accentuat cand sorbitol-DH este redusa sau absenta (retina, cristalin, rinichi, celule nervoase) cu balonizarea celulelor prin efect osmotic in DZ: cataracta, neuropatia periferica, retinopatia
 cu balonizarea celulelor prin efect osmotic. in DZ: cataracta, neuropatia periferica, retinopatia.")
62
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
ALDOZ- REDUCTAZA SORBITOL- DH

63
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
Metabolismul Galactozei Sursa principala: lactoza (hidrolizata de β-galactozidaza intestinala) din lapte si produse lactate degradarea lizozomala a CH complecsi (GP, GL) Patrunderea Gal in celule- proces insulino-independent
 din lapte si produse lactate. degradarea lizozomala a CH complecsi (GP, GL) Patrunderea Gal in celule- proces insulino-independent.")
64
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
A. Fosforilarea Gal: in prezenta galactokinazei şi a ATP, cu formare de Gal-1-P GALACTOKINAZA

65
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
B. Formarea UDP-Gal pt a patrunde in calea glicolitica in prezenta galactozo-1-fosfat-uridil-transferaza

66
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
UDP-Gal= sursa de atomi de C in glicoliza si GNG; UDP-Gal este convertita la UDP-G în prezenta UDP-hexozo-4-epimerazei UDP-G UDP-GAL

67
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI

68
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
Rolul UDP-Gal în reacţiile de biosinteza In sinteza lactozei, GP, GL, GAG Gal insuficienta (deficit de β-galactozidaza): necesarul de UDP-Gal este acoperit prin acţiunea UDP-hexozo-4-epimerazei asupra UDP-G, sintetizata eficient din G-1-P
: necesarul de UDP-Gal este acoperit prin acţiunea UDP-hexozo-4-epimerazei asupra UDP-G, sintetizata eficient din G-1-P.")
69
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
Galactozo-1-fosfat-uridil-transferaza absenta in galactozemie clasica: Gal-1-P si Gal se acumuleaza in celule Devierea metabolismului spre Galactitol, reactie catalizata de aldoz-reductaza aceeasi E ce asigura conversia G la sorbitol

70
METABOLISMUL FRUCTOZEI ŞI GALACTOZEI
Sinteza lactozei: in ap. Golgi sub actiunea lactoz-sintazei (in prezenta ambelor subunitati) la nivelul glandei mamare in lactatie. Proteina B: N-acetil-lactozamina
 la nivelul glandei mamare in lactatie. Proteina B: N-acetil-lactozamina.")
71
SINTEZA LACTOZEI Proteina B/α-LA= α Lactalbumina (+ de prolactina)
LACTOZ-SINTAZA 2 subunitati Proteina B/α-LA= α Lactalbumina (+ de prolactina) Proteina A/GT= galactozil- transferaza
 Proteina A/GT= galactozil- transferaza.")
72
CALEA PENTOZOFOSFATILOR ŞI A NADPH
Cale complementară citoplasmatică şi parţial reversibilă de oxidare a glucozei 2 reacţii oxidative ireversibile şi o serie de interconversii glucid-fosfat reversibile Fără rol energetic, cu importanţă funcţională deosebită ( în ficat, 30% din G este catabolizată astfel)
")
73
CALEA PENTOZOFOSFATILOR ŞI A NADPH
Permite inter-conversiunea ozelor: hexoze-pentoze sau pentoze, tetroze, trioze în hexoze cu degradarea ulterioară pe calea glicolizei Sursă de NADPH (nicotinamid-adenin-dinucleotid fosfat) necesar biosintezelor reductive de acizi graşi, colesterol şi de ribozo-5-fosfat necesar în biosinteza ARN, ribonucleotidelor Localizare intracelulara si tisulara. Reactiile caii pentozofosfatilor se desfasoara in citoplasma deoarece aici se desfasoara procesele la care participa NADPH,H+.
 necesar biosintezelor reductive de acizi graşi, colesterol şi de ribozo-5-fosfat necesar în biosinteza ARN, ribonucleotidelor. Localizare intracelulara si tisulara. Reactiile caii pentozofosfatilor se desfasoara in citoplasma deoarece aici se desfasoara procesele la care participa NADPH,H+.")
74
CALEA PENTOZOFOSFATILOR ŞI A NADPH
Se desfasoara in doua etape majore: conversia hexozelor la pentoze; conversia pentozelor la hexoze. NADPH este necesar hidroxilarii compusilor straini organismului prin sistemul oxidazelor cu functie mixta, refacerii glutationului redus (GSH); hidroxilarii compusilor straini organismului prin sistemul oxidazelor cu functie mixta, refacerii glutationului redus (GSH);
; hidroxilarii compusilor straini organismului prin sistemul oxidazelor cu functie mixta, refacerii glutationului redus (GSH);")
75
CALEA PENTOZOFOSFATILOR ŞI A NADPH
Reacţiile oxidative ireversibile reacţii cu formare de ribulozo-5-P, CO2, 2 NADPH pt 1 molecula de G-6-P oxidata Importanta in: Fi, GM in lactatie, tesut adipos (biosinteza AG) CSR (sinteza NADPH-dependenta a steroizilor) eritrocite (sinteza NADPH pt a mentine glutationul redus)
 CSR (sinteza NADPH-dependenta a steroizilor) eritrocite (sinteza NADPH pt a mentine glutationul redus)")
76
CALEA PENTOZOFOSFATILOR ŞI A NADPH
A. Dehidrogenarea G-6-P=cale reglatorie principala

77
CALEA PENTOZOFOSFATILOR ŞI A NADPH
NADPH= inhibitor competitiv puternic al G-6-P-DH (creste raportul NADPH/ NADP+) Cresterea necesarului de NADPH cu scaderea raportului NADPH/ NADP+ se intensifica activitatea G-6-P-DH
 Cresterea necesarului de NADPH cu scaderea raportului NADPH/ NADP+ se intensifica activitatea G-6-P-DH.")
78
CALEA PENTOZOFOSFATILOR ŞI A NADPH
B. Sinteza ribulozo-5-P 6-fosfogluconolactona este hidrolizata sub actiunea 6-fosfogluconolacton-hidrolazei 1 pentozo-fosfat (ribulozo-5-fosfat), CO2, 1 NADPH
, CO2, 1 NADPH.")
79
CALEA PENTOZOFOSFATILOR ŞI A NADPH
Reactiile neoxidative reversibile Asigura interconversia glucidelor tri-, tetra, penta, hexa si hepta-carbonice Permit conversia ribulozo-5-P in ribozo-5-P transcetolaza (transfera 2 C), transaldolaza (transfera 3C) permit conversia riboluzo-5-P in GAP si Fr-6-P (intermediari glicolitici)
, transaldolaza (transfera 3C) permit conversia riboluzo-5-P in GAP si Fr-6-P (intermediari glicolitici)")
80
CALEA PENTOZOFOSFATILOR ŞI A NADPH

81
CALEA PENTOZOFOSFATILOR ŞI A NADPH

82
CALEA PENTOZOFOSFATILOR ŞI A NADPH
Conversia pentozelor la hexoze include reactii de izomerizare, transcetolizare, transaldolizare: TRANSCETOLAZA TRANSALDOLAZA

83
CALEA PENTOZOFOSFATILOR ŞI A NADPH
Când necesarul de riboza depaseste necesarul de NADPH, reactiile neoxidative asigura biosinteza ribozo-5-P din GAP si Fr-6-P in absenta reactiilor oxidative

84
CALEA PENTOZOFOSFATILOR ŞI A NADPH
Utilizarea G-6-P fie prin glicoliza fie pe calea pentozofosfatilor depinde de necesitatile celulei pentru ATP, NADPH si ribozo-5-P Daca celula are nevoie si de NADPH,H+ si de ribozo-5-fosfat se desfasoara predominant primele 4 reactii Daca celula necesita mai mult ribozo-5-fosfat decat NADPH, H+ sunt extrase din glicoliza Fr-6-P si GAP pentru a ocoli etapele din calea pentozofosfatilor unde se sintetizeaza NADPH,H+. Se produc doar reactiile 5, 6, 7, 8 Daca este necesar mai mult NADPH,H+, ribozo-5-P este folosit pentru a produce intermediari glicolitici pentru ca G-6-P sa fie folosit in producerea de NADPH,H+.

85
UTILIZAREA NADPH NADP+ difera de NAD+ printr-o singura grupare fosfat atasata ribozei. Rol: interacţiunea NADP+ cu enzimele NADP+ - specifice rap. NADP+ / NADPH intrahep.=0.1, favorizând utilizarea NADPH ca agent reducator in biosinteza rap. NADP+ / NADPH intrahep.=1000, utilizarea NAD+ ca agent oxidant OPO3-

86
UTILIZAREA NADPH 1. Reacţiile de biosinteza reducatoare
O parte din energia moleculei de G-6-P este conservata in NADPH, utilizat in reactiile care necesita prezenta unui donor de electroni cu potential electronic ridicat 2. Reducerea peroxidului de hidrogen (ROS) A. Enzimele ce intervin in reactiile antioxidante de regenerare a glutationului redus (glutation-reductaza), superoxid-dismutaza, catalaza Detoxifierea chimica a peroxidului de hidrogen: glutation-peroxidaza seleniu-dependenta
 A. Enzimele ce intervin in reactiile antioxidante de regenerare a glutationului redus (glutation-reductaza), superoxid-dismutaza, catalaza. Detoxifierea chimica a peroxidului de hidrogen: glutation-peroxidaza seleniu-dependenta.")
87
UTILIZAREA NADPH NADPH,H+ sintetizat in calea pentozofosfatilor este folosit si la reducerea glutationului oxidat: Glutationul redus necesar descompunerii H 2O 2 in cantitate mare ar determina oxidarea Hb la metHb si stres oxidativ H2O2 provine din degradarea xenobioticelor (substante straine organismului), din unele procese biochimice (ex., degradarea bazelor purinice etc.)
, din unele procese biochimice (ex., degradarea bazelor purinice etc.)")
88
UTILIZAREA NADPH ROS au efecte deosebit de nocive la nivel celular:
● denaturarea proteinelor ● peroxidarea lipidelor nesaturate membranare ● modificarea permeabilităţii membranare ● lezarea ADN mitocondrial cu afectarea integrităţii proteinelor din lanţul respirator mitocondrial ● moartea celulară. ROS sunt implicate în numeroase procese patologice precum: neoplaziile, leziunile de reperfuzie, bolile inflamatorii şi senescenţa.

89
UTILIZAREA NADPH Glutationul este un antioxidant intracelular important. Pe lângă rolul de dezactivator al ROS, creşte imunitatea şi protecţia faţă de cataractă, previne cancerul, afectarea hepatică etc Nivelul său scade când stresul oxidativ este major în infecţii, traume, intervenţii chirurgicale, medicaţie. Deficienţa de glutation este deasemenea asociată cu aportul proteic insuficient, diabetul, infecţia HIV, sindromul de detresă respiratorie, cancerul sau fibroza pulmonară idiopatică.

90
UTILIZAREA NADPH Sistemele neenzimatice de apărare împotriva ROS:
► vitaminele A, C şi E. Vitamina E (tocoferolul) este un antioxidant ce previne formarea lipoperoxizilor toxici, reduce colesterolemia şi previne leziunile ATS, efectul său fiind potenţat de asocierea cu vitamina A şi C. Tocoferolul este un antioxidant primar, iar radicalul liber rezultat în momentul contracarării formării radicalilor peroxilipidici reacţionează cu vitamina C, regenerând vitamina E ( ciclul redox Packer). Ascorbatul, β-carotenul şi vitamina E au capacitatea de a reduce ROS în condiţii de laborator. Consumul alimentelor bogate în antioxidanţi s-a corelat cu scăderea riscului afecţiunilor cronice sau pentru anumite cancere. Studiile clinice asupra suplimentelor alimentare cu antioxidanţi nu au stabilit un efect benefic clar, consecinţele pozitive ale consumului de fructe şi legume rezultând, mai probabil, din acţiunea cumulată a mai multor compuşi naturali.
 este un antioxidant ce previne formarea lipoperoxizilor toxici, reduce colesterolemia şi previne leziunile ATS, efectul său fiind potenţat de asocierea cu vitamina A şi C. Tocoferolul este un antioxidant primar, iar radicalul liber rezultat în momentul contracarării formării radicalilor peroxilipidici reacţionează cu vitamina C, regenerând vitamina E ( ciclul redox Packer). Ascorbatul, β-carotenul şi vitamina E au capacitatea de a reduce ROS în condiţii de laborator. Consumul alimentelor bogate în antioxidanţi s-a corelat cu scăderea riscului afecţiunilor cronice sau pentru anumite cancere. Studiile clinice asupra suplimentelor alimentare cu antioxidanţi nu au stabilit un efect benefic clar, consecinţele pozitive ale consumului de fructe şi legume rezultând, mai probabil, din acţiunea cumulată a mai multor compuşi naturali.")
91
UTILIZAREA NADPH C. Sistemul citocromilor P450 (CYP)-monooxigenazei Reprezintă o familie importantă a cărei funcţie este de a cataliza oxidarea unui număr mare de compuşi organici Numele acestei familii de enzime provine de la faptul că absoarbe lumina la o lungime de undă de 450 nm Aceste enzime sunt hemoproteine care catalizează reacţia de monooxigenare plecând de la oxigenul molecular, O2, un atom de oxigen fiind transferat substratului (RH) şi celălalt redus la apă.
 şi celălalt redus la apă.")
92
UTILIZAREA NADPH Reacţia globală catalizată de o enzimă CYP este: RH + O2 + NADPH + H+ → ROH + H2O + NADP+ unde R poate fi un compus steroidian, un medicament, un drog, un metabolit intermediar sau orice moleculă exogenă ( xenobiotică) .
 .")
93
UTILIZAREA NADPH 1.Sistemul enzimatic microzomal al CYP
Este ataşat la membranele reticulului endoplasmic neted, REN, în special hepatic, având rol extrem de important în detoxifierea xenobioticelor ( medicamente, poluanţi, pesticide, substanţe carcinogene), prin hidroxilare în prezenţă de NADPH. Efectele hidroxilării sunt: activarea/ inactivarea medicamentelor creşterea solubilităţii cu facilitarea excreţiei prin urină sau fecale obţinerea situsului de conjugare a unui compus polar (acidul glucuronic), cu creşterea solubilităţii compusului
, prin hidroxilare în prezenţă de NADPH. Efectele hidroxilării sunt: activarea/ inactivarea medicamentelor. creşterea solubilităţii cu facilitarea excreţiei prin urină sau fecale. obţinerea situsului de conjugare a unui compus polar (acidul glucuronic), cu creşterea solubilităţii compusului.")
94
UTILIZAREA NADPH Există sute de citocromi diferiţi, iniţial grupaţi în patru familii: CYP1, CYP2, CYP3, CYP4, cu subfamilii (CYP1A, CYP2D, etc.) şi izoenzime distincte (CYP3A4, CYP2D6, etc.). Fiecare tip de CYP are o funcţie specifică. Au devenit unele dintre cele mai extinse familii de proteine, fiind clasificate după algoritme noi. Exemple: ●CYP2C9 : Metabolizarea anti-inflamatoarelor non steroidiene (AINS), a ibuprofenului, a antidiabeticelor orale, a anticoagulantelor orale (acenocumarol, warfarine), a fenitoinei (anticonvulsivant) şi a antihipertensivelor (losartan). ● CYP2D6 : Metabolismul codeinei în morfină, al anumitor antiaritmice şi antidepresive, al beta-blocantelor şi al neurolepticelor.
 şi izoenzime distincte (CYP3A4, CYP2D6, etc.). Fiecare tip de CYP are o funcţie specifică. Au devenit unele dintre cele mai extinse familii de proteine, fiind clasificate după algoritme noi. Exemple: ●CYP2C9 : Metabolizarea anti-inflamatoarelor non steroidiene (AINS), a ibuprofenului, a antidiabeticelor orale, a anticoagulantelor orale (acenocumarol, warfarine), a fenitoinei (anticonvulsivant) şi a antihipertensivelor (losartan). ● CYP2D6 : Metabolismul codeinei în morfină, al anumitor antiaritmice şi antidepresive, al beta-blocantelor şi al neurolepticelor.")
95
UTILIZAREA NADPH 2.Sistemul enzimatic mitocondrial al CYP Participă la hidroxilarea steroizilor cu creşterea solubilităţii lor la nivelul placentei, ovarelor, corticosuprarenalei, CSR, sinteza acizilor biliari (în ficat) şi la hidroxilarea vitaminei D3 în forma biologic activă ( în rinichi).
 şi la hidroxilarea vitaminei D3 în forma biologic activă ( în rinichi).")
96
2. Sistemele oxigen-dependente
UTILIZAREA NADPH D. Fagocitoza produsa de leucocite 1. Mecanismele oxigen-independente utilizeaza modificarile de pH din fagolizozomi si enzimele lizozomale 2. Sistemele oxigen-dependente sistemul mieloperoxidazei (MPO) cel mai puternic dintre mecanismele bactericide intervine NADP-oxidaza din membrana leucocitara
 cel mai puternic dintre mecanismele bactericide. intervine NADP-oxidaza din membrana leucocitara.")
97
UTILIZAREA NADPH SOD, superoxiddismutaza are localizare eritrocitară, hepatică, cerebrală, fiind cea mai importantă enzimă a vieţii aerobe; este o cuproproteină care catalizează o reacţie de dismutare, prin care anionii superoxid sunt reduşi la apă oxigenată: O2 - + O2− + 2H+ H2O2 + O2 Microorganismele modificate genetic, fără SOD, îşi pierd virulenţa. Inactivarea genetică ("knockout") a SOD generează fenotipuri specifice, variind de la bacterii la şoareci, care furnizează indicii importante cu privire la toxicitatea O2 - in vivo. O2 - este toxic; aproape toate organismele dependente de O2 conţin izoforme ale enzimei. Alte proteine ce pot fi atât oxidate cât şi reduse de superoxid au activitate slabă SOD-like (hemoglobina).
 a SOD generează fenotipuri specifice, variind de la bacterii la şoareci, care furnizează indicii importante cu privire la toxicitatea O2 - in vivo. O2 - este toxic; aproape toate organismele dependente de O2 conţin izoforme ale enzimei. Alte proteine ce pot fi atât oxidate cât şi reduse de superoxid au activitate slabă SOD-like (hemoglobina).")
98
UTILIZAREA NADPH ► catalaza (CAT) descompune excesul de apă oxigenată: 2 H2O2 2 H2O + O2 Este prezentă în toate celulele aerobe ce conţin citocromi ( în peroxizomi şi mitocondrii).
.")
99
UTILIZAREA NADPH E. Sinteza oxidului nitric Arginina, O2, NADPH= substraturi pt NO-sintaza citosolica FMN, FAD, hem si tetrahidrobiopterina= au roluri de coenzima, iar NO si citrulina= produsi finali NO mentine tonusul musculaturii netede, mediaza efectul bactericid al macrofagelor, inhibitor al agregarii plachetare, neurotransmitator

100
UTILIZAREA NADPH NADPH + H + L-ARGININA L-CITRULINA NO

101
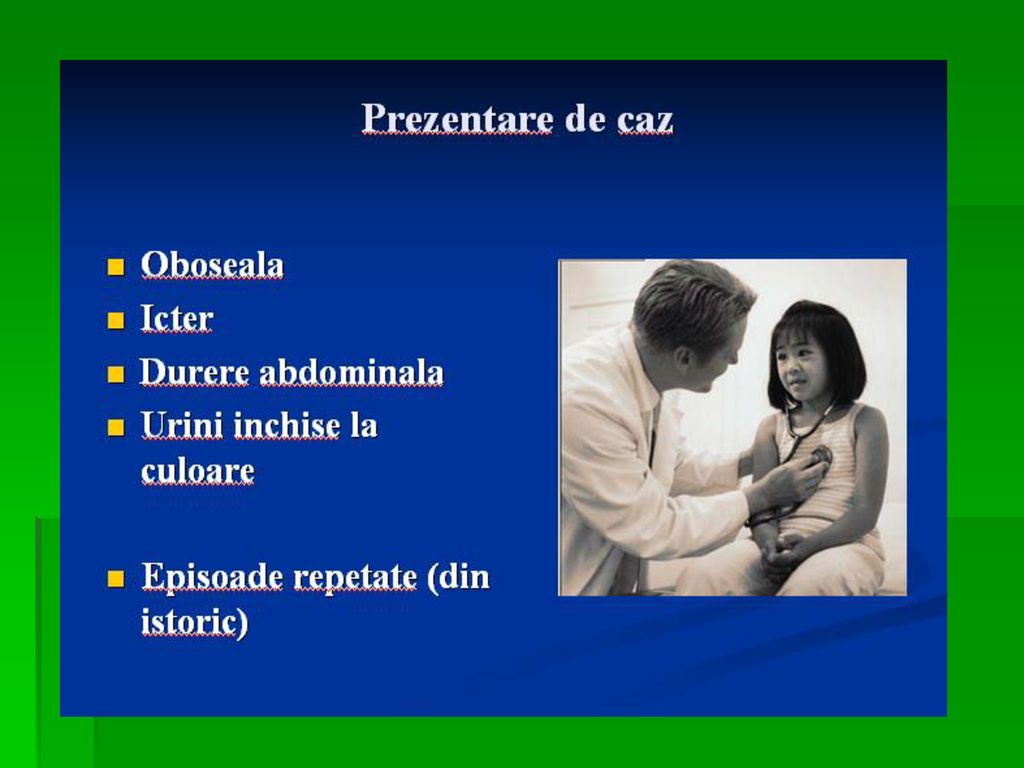
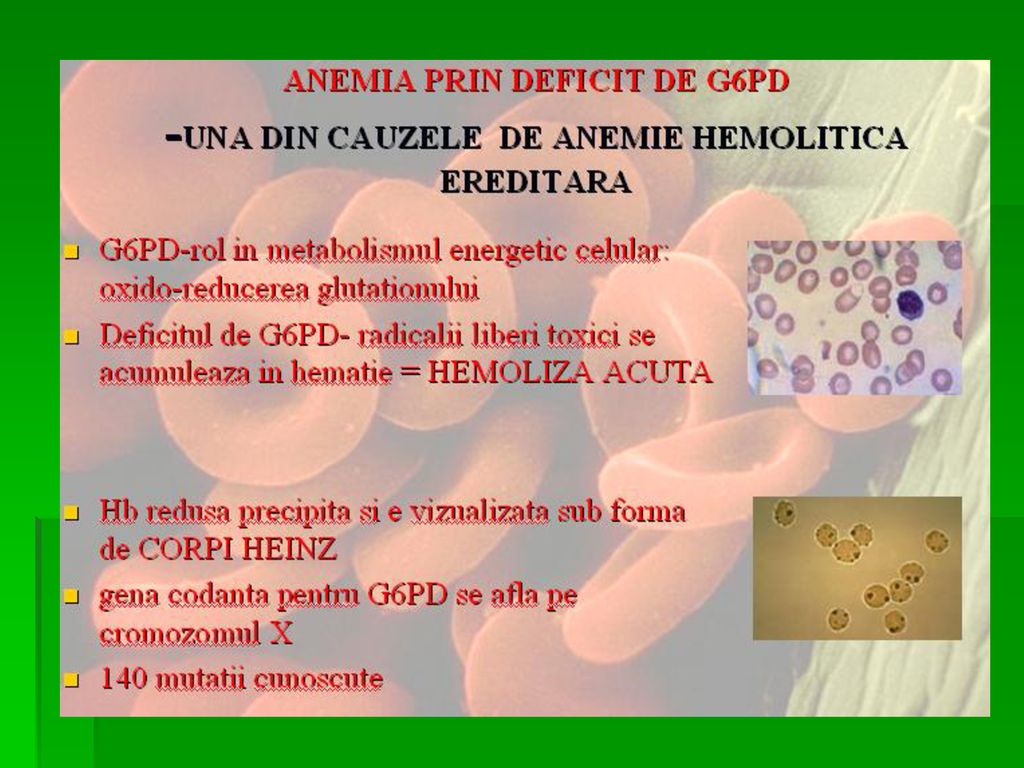
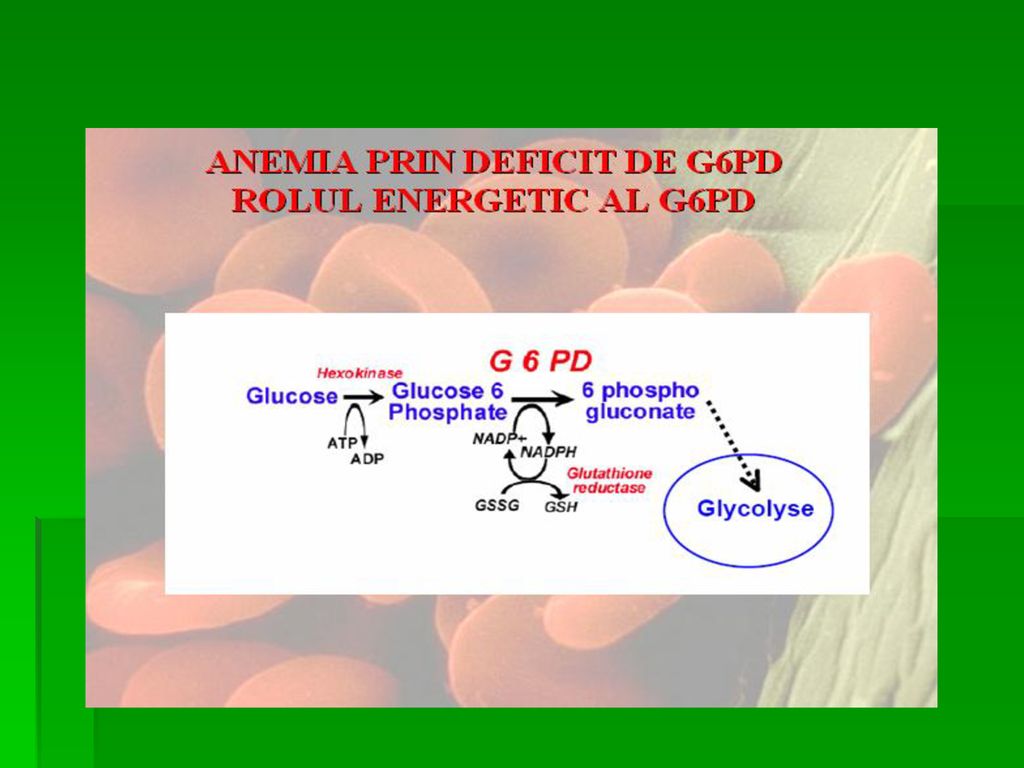
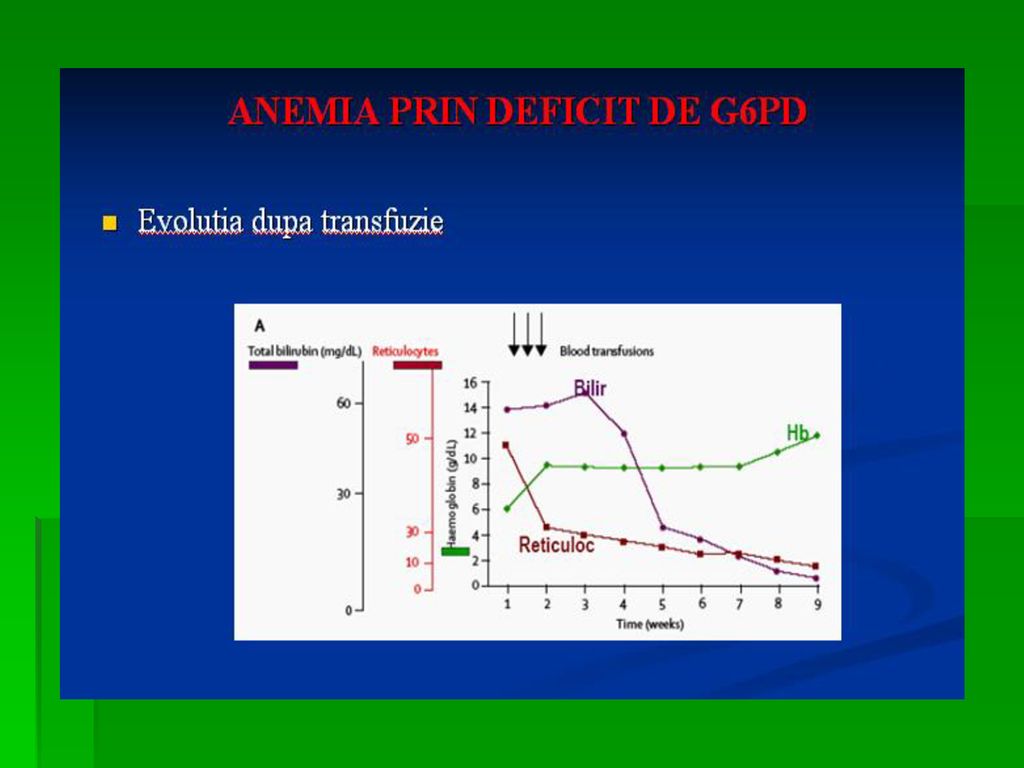
DEFICITUL DE G-6-P- DEHIDROGENAZA

102
DEFICITUL DE G-6-P- DEHIDROGENAZA
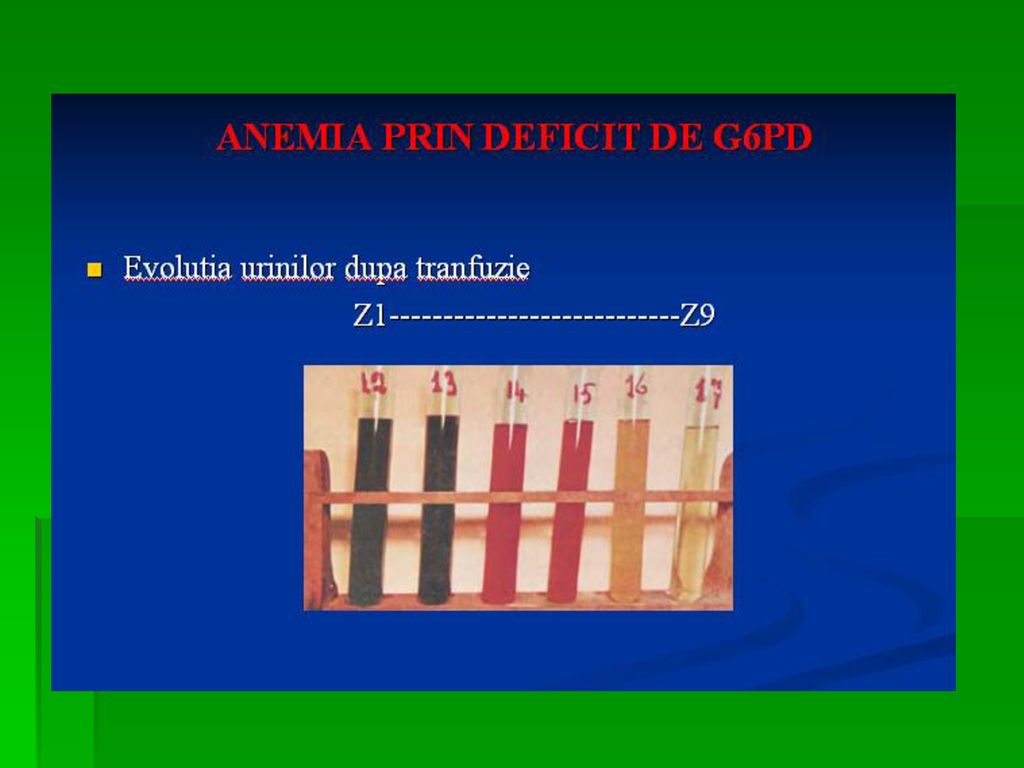
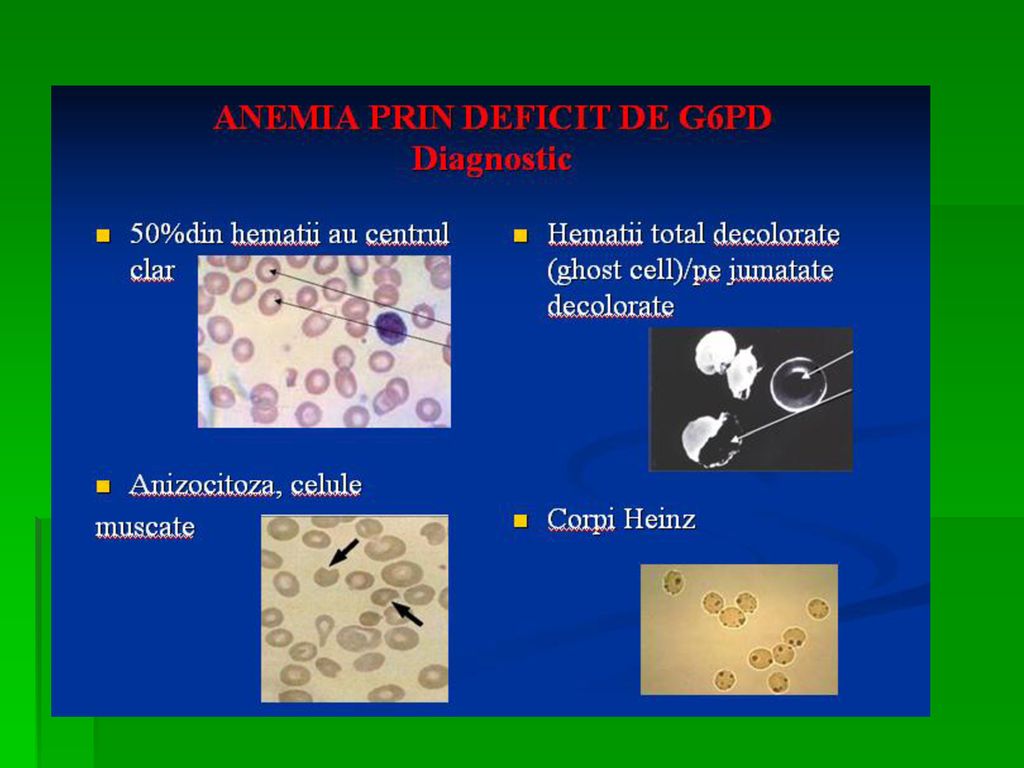
Determina anemia hemolitica. S-a observat ca unii subiecti sunt susceptibili la a manifesta anemie hemolitica acuta la administrarea unor medicamente ca antimalarice, sulfamide, antipiretice, simptomele aparand in ore, datorita deficitului de activitate a acestei enzime Incidenta acestei deficiente este crescuta in zonele geografice unde poate sa apara malaria.

121
CALEA GLUCURONICA Cale secundară de metabolizare a glucozei Acidul Glucuronic este un acid carboxilic. Structura sa este similară cu a glucozei. al 6-lea atom de C al acidului glucuronic este oxidat la acid carboxilic.

122
CALEA GLUCURONICA UDP-G-PIROFOS- FORILAZA UDP-G-DH-AZA GLUCURONOZIL
TRANSFERAZE

123
CALEA GLUCURONICA Sărurile şi esterii acidului glucuronic sunt cunoscute sub denumirea de glucuronaţi Acidul glucuronic nu trebuie confundat cu acidul gluconic, un acid carboxilic linear ce rezultă din oxidarea unui alt atom de C al glucozei Acidul glucuronic serveşte la sinteza PG şi proceselor de detoxifiere din ficat, rinichi şi intestin Glucuronidarea- formarea de legături glicozidice cu diverşi metaboliţi toxici, mărindu-le solubilitatea şi favorizând astfel eliminarea lor din organism.

124
CALEA GLUCURONICA Glucuronozide= compuşi cu polaritate crescută
Reacţia are loc sub acţiunea enzime= glucuronozil-transferaze Glucuronidarea este importantă în metabolismul xenobioticelor: medicamente, bilirubină, androgeni, estrogeni, hormoni tiroidieni, săruri biliare etc.

125
GLUCURONOZIL- TRANSFERAZE

126
CALEA GLUCURONICA REACTIA DE GLUCURONIDARE

127
CALEA GLUCURONICA REACTIA DE GLUCURONIDARE

128
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
GAG- molecule heteropolizaharidice mari ce conţin pe suprafaţa lor sarcini electrice negative Se asociaza cu proteine= proteoglicani (PG) ce contin CH (95%) Glicoproteinele (GP) contin predominant proteine si o cantitate redusa de CH
 ce contin CH (95%) Glicoproteinele (GP) contin predominant proteine si o cantitate redusa de CH.")
129
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
GAG leagă o cantitate mare de apa formand o matrice gelatinoasa ce constituie structura de baza a substantei fundamentale (impreuna cu colagenul formeaza matricea extracelulara) proprietatile lubrefiante, vascoase ale secretiilor mucoase se datoreaza partial GAG (MPZ-initial)
 proprietatile lubrefiante, vascoase ale secretiilor mucoase se datoreaza partial GAG (MPZ-initial)")
130
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Structura GAG GAG sunt polizaharide neramificate formate din unitati repetitive dizaharidice din care una este intotdeauna o ozamina (glucozamina sau galactozamina) iar cealalta un acid uronic (cu exceptia unui singur GAG-keratan-sulfatul ce contine Gal), acid glucuronic sau acid iduronic Moleculele pot sa contina grupe sulfat legate esteric la oxigen sau la azot ce impreuna cu gruparile (COOH) confera caracterul puternic electronegativ al GAG Acizii D-glucuronic si epimerul la C 5 -iduronic sunt izomeri.
 iar cealalta un acid uronic (cu exceptia unui singur GAG-keratan-sulfatul ce contine Gal), acid glucuronic sau acid iduronic. Moleculele pot sa contina grupe sulfat legate esteric la oxigen sau la azot ce impreuna cu gruparile (COOH) confera caracterul puternic electronegativ al GAG. Acizii D-glucuronic si epimerul la C 5 -iduronic sunt izomeri.")
131
GLICOZAMINOGLICANI ŞI GLICOPROTEINE

132
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Lanţurile de polizaharide se extind din cauza sarcinilor negative si sunt inconjurate de apa Prin comprimarea unei solutii de GAG, apa este "stoarsa" in exterior, iar GAG ocupa un volum mai mic La decompresiune, GAG revin la volumul initial (hidratat) ca urmare a fortei de respingere dintre sarcinile negative Elasticitatea lichidului sinovial sau a corpului vitros al globului ocular COMPRESIUNE DECOMPRESIUNE
 ca urmare a fortei de respingere dintre sarcinile negative. Elasticitatea lichidului sinovial sau a corpului vitros al globului ocular. COMPRESIUNE. DECOMPRESIUNE.")
133
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Clasificarea GAG 6 clase principale în funcţie de structura monomerilor, tipul legaturilor glicozidice, gradul si localizarea gruparilor sulfat 1. Condroitin sulfati (4-sulfat si 6-sulfat) 2. Keratan sulfati I si II 3. Acid hialuronic 4. Dermatan sulfati 5. Heparina 6. Heparan sulfatul
 2. Keratan sulfati I si II. 3. Acid hialuronic. 4. Dermatan sulfati. 5. Heparina. 6. Heparan sulfatul.")
134
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Condroitin sulfatii (4-sulfat sau 6-sulfat) se gasesc in cartilaje, oase, cornee (in limba greaca chondros=cartilaj) Sunt GAG mai mici care pot sa formeze cu acidul hialuronic si cu proteinele agregate mari in tesutul conjunctiv (de exemplu, agrecan) Contin acid glucuronic (GlcUA), N-acetilgalactozamina (GalNAc) 4-sulfatata sau 6- sulfatata legate β1,3-glicozidic. Contribuie la rezistenta la intindere a cartilajelor, tendoanelor, ligamentelor, peretilor aortei.
 se gasesc in cartilaje, oase, cornee (in limba greaca chondros=cartilaj) Sunt GAG mai mici care pot sa formeze cu acidul hialuronic si cu proteinele agregate mari in tesutul conjunctiv (de exemplu, agrecan) Contin acid glucuronic (GlcUA), N-acetilgalactozamina (GalNAc) 4-sulfatata sau 6- sulfatata legate β1,3-glicozidic. Contribuie la rezistenta la intindere a cartilajelor, tendoanelor, ligamentelor, peretilor aortei.")
135
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Condroitin -4- sulfatii şi condroitin -6- sulfatii GlcUA

136
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
2. Keratan sulfatii I si II (in limba greaca keras=corn). Keratan sulfatul I este legat de asparagina si se gaseste in cornee iar keratan sulfatul II formeaza legatura O-glicozidica cu treonina si este intalnit in tesutul conjunctiv Se mai gasesc in structurile cornoase: coarne, par, copite, unghii, gheare. Contin N-acetilglucozamina (GlcNAc) si Gal legate 1,3-glicozidic. Nu au acizi uronici.
. Keratan sulfatul I este legat de asparagina si se gaseste in cornee iar keratan sulfatul II formeaza legatura O-glicozidica cu treonina si este intalnit in tesutul conjunctiv. Se mai gasesc in structurile cornoase: coarne, par, copite, unghii, gheare. Contin N-acetilglucozamina (GlcNAc) si Gal legate 1,3-glicozidic. Nu au acizi uronici.")
137
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
2. Keratan sulfatii I si II

138
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
3. Acidul hialuronic (din limba greaca, hyalos = sticla) se gaseste in lichidul sinovial, umoare vitroasa, tesutul conjunctiv lax. Contine aproximativ resturi de acid glucuronic si N-acetilglucozamina legate β1,3-glicozidic (cel mai lung GAG) Singurul GAG care nu contine grupe sulfat si nu se leaga de proteina. Formeaza solutii clare, vascoase ce lubrefiaza articulatiile si dau aspectul gelatinos umorii vitroase. Contribuie la elasticitatea si rezistenta la intindere a tendoanelor alaturi de late componente ale matricei extracelulare Formeaza arhitecturi moleculare “gigant” prin asamblare cu proteoglicani continand cu precadere condroitin sulfati.
 se gaseste in lichidul sinovial, umoare vitroasa, tesutul conjunctiv lax. Contine aproximativ resturi de acid glucuronic si N-acetilglucozamina legate β1,3-glicozidic (cel mai lung GAG) Singurul GAG care nu contine grupe sulfat si nu se leaga de proteina. Formeaza solutii clare, vascoase ce lubrefiaza articulatiile si dau aspectul gelatinos umorii vitroase. Contribuie la elasticitatea si rezistenta la intindere a tendoanelor alaturi de late componente ale matricei extracelulare. Formeaza arhitecturi moleculare gigant prin asamblare cu proteoglicani continand cu precadere condroitin sulfati.")
139
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
3. Acidul hialuronic Legaturile glicozidice ale acidului hialuronic pot fi hidrolizate de catre hialuronidaza bacteriana, ceea ce duce la alterarea capacitatii de filtru selectiv a substantei fundamentale si la expunerea tesuturilor la invazia bacteriana. Hialuronidaza se mai gaseste in sperma cu rolul de a permite penetrarea ovulului. GlcUA

140
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
4. Dermatan sulfat Distribuit în tegument, vase sangvine si valvele cardiace IdUA

141
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
5. Heparina (in limba greaca hepar=ficat) contine glucozamina N-sulfatata si mai putin acetilata, acid glucuronic si acid iduronic (predominant) Se gaseste in mastocite, ficat, plaman, piele. Este cel mai electronegativ GAG Are proprietati anticoagulante folosindu-se in forma pura pentru recoltarea probelor de sange.
 contine glucozamina N-sulfatata si mai putin acetilata, acid glucuronic si acid iduronic (predominant) Se gaseste in mastocite, ficat, plaman, piele. Este cel mai electronegativ GAG. Are proprietati anticoagulante folosindu-se in forma pura pentru recoltarea probelor de sange.")
142
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
6. Heparan sulfatul contine glucozamina mai putin N-sulfatata si, mai ales, acid glucuronic Se gaseste in fibroblasti, peretii vaselor. Este component al membranelor bazale, sinapselor.

143
GLICOZAMINOGLICANI ŞI GLICOPROTEINE

144
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Sinteza GAG in sistemul Golgi Alungirea lanturilor polizaharidice: adaugarea secventiala si alternativa de glucide acide si amino-glucide cedate de derivatii UDP ai acestora Sinteza GAG- similara sintezei de glicogen cu diferenta ca sunt excretati in celula

145
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
A. Sinteza amino-glucidelor sunt constituienti principali ai GAG, GP, GL, oligozaharidelor, AB 1. N-acetil-glucozamina (GlcNAc); GalNAc Fr-6-P= monozaharid precursor (pt acidul sialic-NANA sau acidul neuraminic) Glu Glucoz- amino- 6-P Gln Glucoz- amino- 1-P UTP UDP- GLUCOZAMINA G-6-P Fr-6-P GAG G Acid sialic Gangliozide GP CMP- NANA GAG, GP
; GalNAc. Fr-6-P= monozaharid precursor (pt acidul sialic-NANA sau acidul neuraminic) Glu. Glucoz- amino- 6-P. Gln. Glucoz- amino- 1-P. UTP. UDP- GLUCOZAMINA. G-6-P. Fr-6-P. GAG. G. Acid sialic. Gangliozide. GP. CMP- NANA. GAG, GP.")
146
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
B. Sinteza glucidelor acide: acidul D-glucuronic, L-iduronic C. Sinteza miezului proteic la suprafata RER dupa care patrunde in interiorul acestuia unde va fi glicozilat in prezenta transferazelor sist. Golgi D. Sinteza lantului de CH E. Atasarea gruparilor sulfat are loc dupa ce monozaharidul a fost incorporat in structura lantului in formare in prezenta sulfotransferazelor Sursa: 3'-fosfoadenozil-5'-fosfosulfat (PAPS)
")
147
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Sulfatarea defectuoasa a lantului de GAG produce condrodistrofii Degradarea GAG: in lizozomi (pH=5) in prezenta hidrolazelor acide A. Fagocitoza GAG extracelulari B. Degradarea lizozomala a GAG: lanturile polizaharidice sunt clivate sub actiunea endoglicozidazelor, formandu-se oligozaharide
 in prezenta hidrolazelor acide. A. Fagocitoza GAG extracelulari. B. Degradarea lizozomala a GAG: lanturile polizaharidice sunt clivate sub actiunea endoglicozidazelor, formandu-se oligozaharide.")
148
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Mucopolizaharidozele (MPZ) - afectiuni ereditare cu evolutie clinica progresiva acumulare de GAG in tesuturi cu deformari scheletale si retard mintal, deces precoce in forma homozigota cauzate de deficitul hidrolazelor lizozomale (se pot masura) implicate in degradarea heparan sulfatului si dermatan sulfatului a prezenta oligozaharidelor in urina Tt: transplant de MO si sange placentar macrofagele transplantate sintetizeaza sulfataza necesara degradarii GAG
 - afectiuni ereditare cu evolutie clinica progresiva. acumulare de GAG in tesuturi cu deformari scheletale si retard mintal, deces precoce in forma homozigota. cauzate de deficitul hidrolazelor lizozomale (se pot masura) implicate in degradarea heparan sulfatului si dermatan sulfatului a. prezenta oligozaharidelor in urina. Tt: transplant de MO si sange placentar. macrofagele transplantate sintetizeaza sulfataza necesara degradarii GAG.")
149
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
MPZ se pot asocia cu o lipidoza sau glico-protein-oligozaharidoza (enzimele lizozomale necesare catabolizarii GAG pasticipa si la degradarea GL sau a GP) MPZ I (Hurler): deficit de α-iduronidaza MPZ II (Hunter): deficit de iduronat-sulfataza MPZ III (Sanfilippo): subtipurile A, B, C MPZ IV (Sly): deficit de β-glucuronidaza
 MPZ I (Hurler): deficit de α-iduronidaza. MPZ II (Hunter): deficit de iduronat-sulfataza. MPZ III (Sanfilippo): subtipurile A, B, C. MPZ IV (Sly): deficit de β-glucuronidaza.")
150
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Glicoproteinele sunt proteine conjugate ce contin resturi de oligozaharide (2-10 resturi glucidice) nu neaparat electronegative, legate covalent de lantul polipeptidic catene liniare de CH; IgG contin <4% CH, GP gastrica umana (mucina) > 80% CH GP membranare participa la: recunoasterea de catre hormoni, virusuri antigenitatea de S matricea extracelulara, mucinele g-int si urogenitale
 nu neaparat electronegative, legate covalent de lantul polipeptidic. catene liniare de CH; IgG contin <4% CH, GP gastrica umana (mucina) > 80% CH. GP membranare participa la: recunoasterea de catre hormoni, virusuri. antigenitatea de S. matricea extracelulara, mucinele g-int si urogenitale.")
151
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Structura GP: oligozaharide atasate N- (Asn) si O-(OH din Ser sau treonina)glicozidic de proteine In colagen: leg. O-glicozidica intre Gal/Glc si OH din hidroxilizina Oligozaharidele O-linkate: pe S eritrocitelor (determinantii de grup sangvin) Oligozaharidele N-linkate: complexe (GlcNAc, L-fucoza si NANA) si cu un continut ridicat de Man
 si O-(OH din Ser sau treonina)glicozidic de. proteine. In colagen: leg. O-glicozidica intre Gal/Glc si OH din hidroxilizina. Oligozaharidele O-linkate: pe S eritrocitelor (determinantii de grup sangvin) Oligozaharidele N-linkate: complexe (GlcNAc, L-fucoza si NANA) si cu un continut ridicat de Man.")
152
GLICOZAMINOGLICANI ŞI GLICOPROTEINE

153
GLICOZAMINOGLICANI ŞI GLICOPROTEINE
Degradarea lizozomala a GP este similara cu cea a GAG: enzime hidrolitice lizozomale Deficientele enzimatice ereditare conduc la oligozaharidoze caracterizate prin acumularea intralizozomala de GP partial digerate