Κατέβασμα παρουσίασης
1
Εισαγωγή στα Αντιβιοτικά -Γενικά -Μηχανισμοί δράσης -Μηχανισμοί αντοχής

2
ΑΝΤΙΒΙΟΤΙΚΑ ή ΧΗΜΕΙΟΘΕΡΑΠΕΥΤΙΚΑ ή ΑΝΤΙΜΙΚΡΟΒΙΑΚΟΙ ΠΑΡΑΓΟΝΤΕΣ ή ΑΝΤΙΒΑΚΤΗΡΙΑΚΟΙ ΠΑΡΑΓΟΝΤΕΣ Ποιός είναι ο πλέον σωστός όρος ;

3
Αρχές 20ου αιώνα: Συστηματική προσπάθεια ανάπτυξης αντιμικροβιακών φαρμάκων THERAPIA MAGNA STERILISANS “THERAPIA MAGNA STERILISANS” Τέλη 20ου αιώνα: Απαισιοδοξία για το μέλλον των αντιβιοτικών “The end of the miracle drugs….Bugs have won…etc.”

4
Τι είναι αντιβιοτικό ; ή Ποιά “ουσία” δικαιούται αυτό το όνομα ;

5
ΑΙΤΗΜΑΤΑ FLEMING -Εκλεκτική τοξικότητα -Προσέγγιση θέσης λοίμωξης -Σταθερότητα στα βιολογικά υγρά

6
ΤΑ ΔΙΑΘΕΣΙΜΑ ΑΝΤΙΒΙΟΤΙΚΑ ΔΕΝ ΕΙΝΑΙ ΙΔΑΝΙΚΑ Η βασική ιδιότητα που πρέπει να έχει ένα αντιβιοτικό είναι η εκλεκτική τοξικότητα. -Ωστόσο, αντιβιοτικό με απολύτως εκλεκτική τοξικότητα δεν υπάρχει. -Μόνο τα β-λακταμικά αντιβιοτικά εκδηλώνουν σαφή εκλεκτική τοξικότητα. Τα υπόλοιπα εμφανίζουν «επαρκή» εκλεκτική τοξικότητα

7
Δράση των αντιβιοτικών Βακτηριοκτόνα, βακτηριοστατικά ή απλώς αντιβακτηριακά; Συνήθως βακτηριοκτόνα β-Λακτάμες, Αμινογλυκοσίδες, Κινολόνες, Γλυκοπεπτίδια Συνήθως βακτηριοστατικά Τετρακυκλίνες, Σουλφοναμίδες, Τριμεθοπρίμη, Μακρολίδες

8
Δεν υπάρχει σαφής διαχωριστική γραμμή Ενα αντιβιοτικό μπορεί να είναι κατά περίπτωση βακτηριοκτόνο ή βακτηριοστατικό Αιτίες -Ο ακριβής μηχανισμός δράσης για πολλά αντιβιοτικά δεν είναι γνωστός -Υπάρχει συχνά αναντιστοιχία μεταξύ in vitro και in vivo δράσης

9
ΠΑΛΑΙΑ ΚΑΙ “ΝΕΑ” ΑΝΤΙΒΙΟΤΙΚΑ ΟΜΑΔΑΠΡΟΓΟΝΟΣΣΥΓΧΡΟΝΟ ΑΝΤΙΒΙΟΤΙΚΟ -------------------------------------------------------------------------------------- β-ΛακτάμεςΠενικιλλίνη (1929)Μεροπενέμη ΣουλφοναμίδεςΣουλφοναμίδη (1932)Σουλφαμεθοξαζόλη Αμινογλυκ.Στρεπτομυκίνη (1942)Αμικασίνη ΤετρακυκλίνηΑουρεομυκίνη (1948)Μινοκυκλίνη ΜακρολίδεςΕρυθρομυκίνη (1952)Αζιθρομυκίνη ΓλυκοπεπτίδιαΒανκομυκίνη (1956)Τεϊκοπλανίνη ΚινολόνεςΝαλιδιξικό οξύ (1958)Τροβαφλοξασίνη ---------------------------------------------------------------------------------------
Μεροπενέμη ΣουλφοναμίδεςΣουλφοναμίδη (1932)Σουλφαμεθοξαζόλη Αμινογλυκ.Στρεπτομυκίνη (1942)Αμικασίνη ΤετρακυκλίνηΑουρεομυκίνη (1948)Μινοκυκλίνη ΜακρολίδεςΕρυθρομυκίνη (1952)Αζιθρομυκίνη ΓλυκοπεπτίδιαΒανκομυκίνη (1956)Τεϊκοπλανίνη ΚινολόνεςΝαλιδιξικό οξύ (1958)Τροβαφλοξασίνη")
10
Nέα αντιβιοτικά Οξαζολιδινόνες (Linezolid) Στρεπτογραμίνες (Quinopristin/Dalfopristin) Κετολίδες (Telithromycin) Γλυκυλο-γλυκίνες (Tigecyclin) Λιποπεπτίδια Λιπογλυκοπεπτίδια Εβερνινομυκίνες -Επισήμανση Ολα τα νέα αντιβιοτικά είναι δραστικά κυρίως κατά των Gram-θετικών
 Στρεπτογραμίνες (Quinopristin/Dalfopristin) Κετολίδες (Telithromycin) Γλυκυλο-γλυκίνες (Tigecyclin) Λιποπεπτίδια Λιπογλυκοπεπτίδια Εβερνινομυκίνες -Επισήμανση Ολα τα νέα αντιβιοτικά είναι δραστικά κυρίως κατά των Gram-θετικών")
11
Πως επιλέγονται οι στόχοι των αντιβιοτικών έτσι ώστε αυτά να εκδηλώνουν εκλεκτική τοξικότητα ?

12
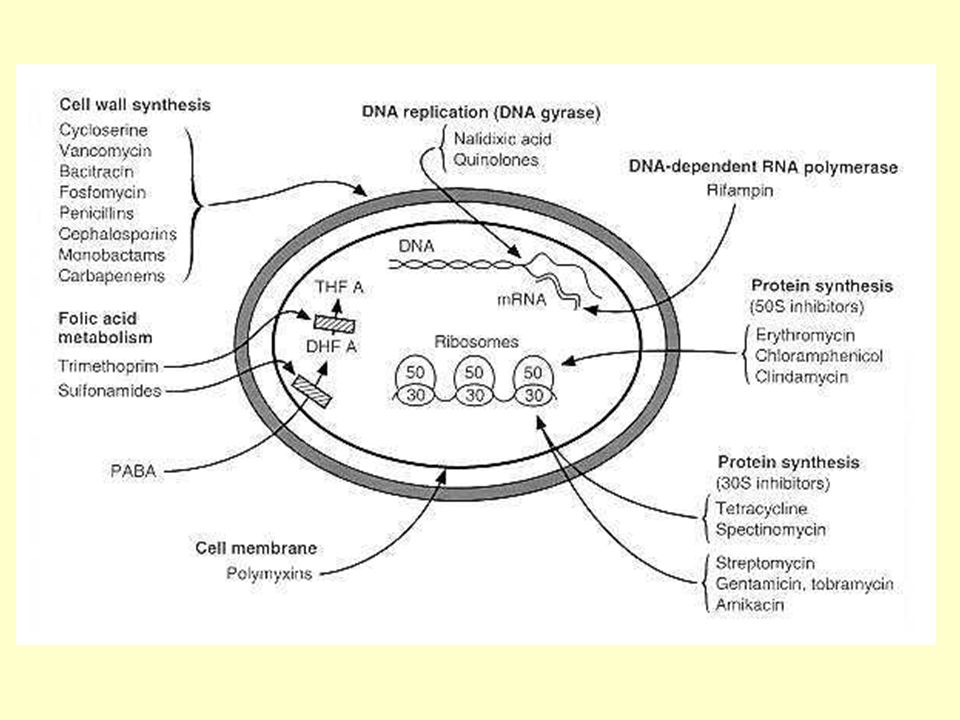
ΣΤΟΧΟΙ ΤΩΝ ΑΝΤΙΒΙΟΤΙΚΩΝ -Κυτταρικό τοίχωμα [β-Λακτάμες, Γλυκοπεπτίδια] -Πρωτεϊνοσύνθεση [Αμινογλυκοσίδες, Μακρολίδες, Τετρακυκλινες, Λινκοσαμίδες, Χλωραμφαινικόλη] -Σύνθεση DNA [Κινολόνες] -Σύνθεση φυλικού [Σουλφοναμίδες, Τριμεθοπρίμη] -RNA πολυμεράση [Ριφαμπικίνη] -Κυτταροπλασματική μεμβράνη [Κολιστίνη]
![ΣΤΟΧΟΙ ΤΩΝ ΑΝΤΙΒΙΟΤΙΚΩΝ -Κυτταρικό τοίχωμα [β-Λακτάμες, Γλυκοπεπτίδια] -Πρωτεϊνοσύνθεση [Αμινογλυκοσίδες, Μακρολίδες, Τετρακυκλινες, Λινκοσαμίδες, Χλωραμφαινικόλη] -Σύνθεση DNA [Κινολόνες] -Σύνθεση φυλικού [Σουλφοναμίδες, Τριμεθοπρίμη] -RNA πολυμεράση [Ριφαμπικίνη] -Κυτταροπλασματική μεμβράνη [Κολιστίνη]](http://images.slideplayer.gr/46/11667517/slides/slide_12.jpg "ΣΤΟΧΟΙ ΤΩΝ ΑΝΤΙΒΙΟΤΙΚΩΝ -Κυτταρικό τοίχωμα [β-Λακτάμες, Γλυκοπεπτίδια] -Πρωτεϊνοσύνθεση [Αμινογλυκοσίδες, Μακρολίδες, Τετρακυκλινες, Λινκοσαμίδες, Χλωραμφαινικόλη] -Σύνθεση DNA [Κινολόνες] -Σύνθεση φυλικού [Σουλφοναμίδες, Τριμεθοπρίμη] -RNA πολυμεράση [Ριφαμπικίνη] -Κυτταροπλασματική μεμβράνη [Κολιστίνη]")
14
Θέματα προς συζήτηση α) Γιατί οι στόχοι των αντιβιοτικών είναι περιορισμένοι ? Δεν θα μπορούσαν να κατασκευαστούν αντιβιοτικά που θα είχαν ως στόχους π.χ. την αναπνευστική αλυσίδα ή τη γλυκόλυση ? β) Σε σχέση με τα αντιβιοτικά, τα αντιικά και τα αντιπαρασιτικά είναι πολύ λιγότερα. Γιατί ?
 Σε σχέση με τα αντιβιοτικά, τα αντιικά και τα αντιπαρασιτικά είναι πολύ λιγότερα. Γιατί .")
15
Το τοίχωμα των Gram-θετικών

16
Το τοίχωμα των Gram-αρνητικών

17
Δομή της πεπτιδογλυκάνης

18
Τόπος δράσης των β-λακταμών, γλυκοπεπτιδίων, D-κυκλοσερίνης, βακιτρακίνης κ’ φωσφομυκίνης

19
β-ΛΑΚΤΑΜΙΚΑ ΑΝΤΙΒΙΟΤΙΚΑ -Αναστολή της δράσης των τρανσπεπτιδασών (PBPs) -To βακτήριο, ενώ συνθέτει αλυσίδες πεπτιδογλυκάνης, δεν είναι ικανό να τις διασταυρώσει.
 -To βακτήριο, ενώ συνθέτει αλυσίδες πεπτιδογλυκάνης, δεν είναι ικανό να τις διασταυρώσει.")
20
H στερεοχημική ομοιότητα D-Ala-D-Ala και β-λακταμικού δακτύλίου

21
Penicillium chrysogenum, P. notatum Cephalosporium acremonium Streptomyces clavuligerus Streptomyces cattleya Gluconobacter, Chromobacterium

22
* Οι β-λακτάμες είναι βακτηριοκτόνες σε μεταβολικώς ενεργά κύτταρα -Το βακτηριοκτόνο αποτέλεσμα εξαρτάται από τη συμμετοχή του κυττάρου. -Η αναστολή του τελικού σταδίου σύνθεσης επάγει την αυτόλυση της πεπτιδογλυκάνης (πρακτικά το κύτταρο αυτοκτονεί) Π.Χ. Στα E. coli και S. pneumoniae ενεργοποιούνται οι υδρολάσες της πεπτιδογλυκάνης. Στον S. aureus ενεργοποιείται το σύστημα διαχωρισμού των θυγατρικών κυττάρων.
 Π.Χ. Στα E. coli και S. pneumoniae ενεργοποιούνται οι υδρολάσες της πεπτιδογλυκάνης. Στον S. aureus ενεργοποιείται το σύστημα διαχωρισμού των θυγατρικών κυττάρων..")
23
Γλυκοπεπτίδια (Βανκομυκίνη - Τεϊκοπλανίνη) Προέρχονται από διάφορα είδη ακτινομυκήτων (Amycolaptosis, Actinoplanes κά) Η κάλυψη της ομάδας -D-Ala-D-Ala από τα γλυκοπεπτίδια δεν επιτρέπει την προσέγγιση τρανσγλυκολασών και τρανσπεπτιδασών διακόπτωντας έτσι το τελικό στάδιο διασταύρωσης των αλυσίδων πεπτιδογλυκάνης
 Προέρχονται από διάφορα είδη ακτινομυκήτων (Amycolaptosis, Actinoplanes κά) Η κάλυψη της ομάδας -D-Ala-D-Ala από τα γλυκοπεπτίδια δεν επιτρέπει την προσέγγιση τρανσγλυκολασών και τρανσπεπτιδασών διακόπτωντας έτσι το τελικό στάδιο διασταύρωσης των αλυσίδων πεπτιδογλυκάνης")
24
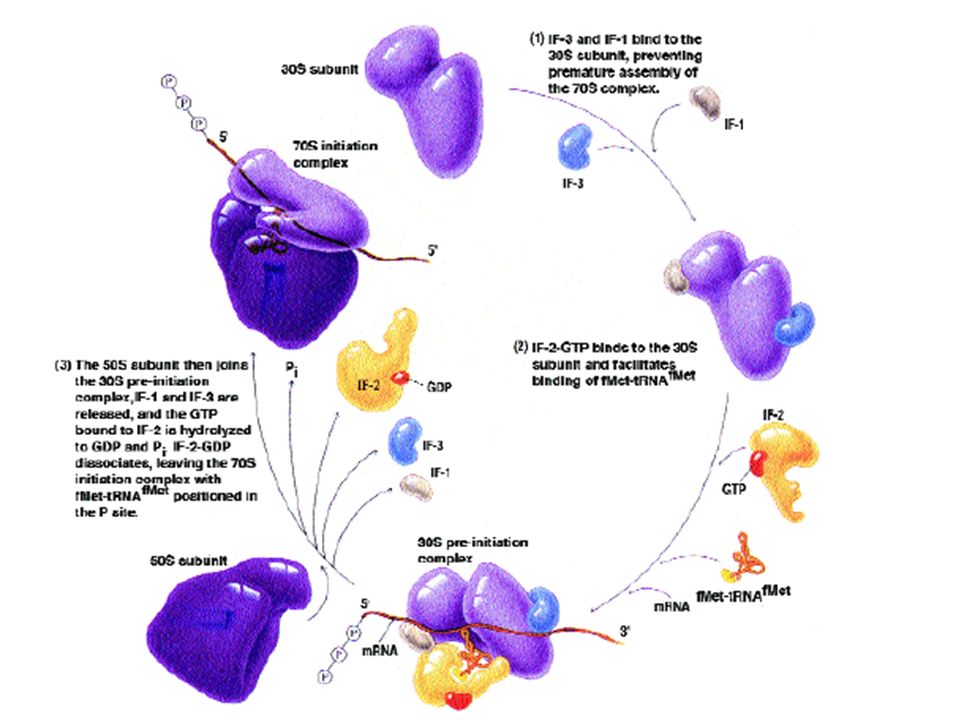
Πρωτεϊνοσύνθεση και αντιβιοτικά-αναστολείς Διαδικασία βακτηριακής πρωτεϊνοσύνθεσης : -Η μετάφραση των mRNA σε πρωτεϊνες γίνεται στα ριβοσώματα. Κάθε κύτταρο περιέχει μεγάλο αριθμό ριβοσωμάτων (π.χ. ένα κύτταρο E. coli διαθέτει περίπου 30.000 ριβοσώματα). -Το ριβόσωμα περιλαμβάνει δύο υπομονάδες, τη 30S (μικρή υπομονάδα) και τη 50S (μεγάλη υπομονάδα) που συνίστανται από rRNA και πρωτεϊνες. - Το πλήρες βακτηριακό ριβόσωμα (70S) περιλαμβάνει τις δύο υπομονάδες, το μεταφραζόμενο mRNA και tRNA συνδεδεμένο με αμινοξέα. -Η πρωτεϊνοσύνθεση γίνεται με ταχύτατο ρυθμό (300-450 πεπτιδικοί δεσμοί ανά λεπτό) σε τρία στάδια: α) έναρξη, β) επιμήκυνση της πεπτιδικής αλυσίδας και γ) τερματισμός.
. -Το ριβόσωμα περιλαμβάνει δύο υπομονάδες, τη 30S (μικρή υπομονάδα) και τη 50S (μεγάλη υπομονάδα) που συνίστανται από rRNA και πρωτεϊνες. - Το πλήρες βακτηριακό ριβόσωμα (70S) περιλαμβάνει τις δύο υπομονάδες, το μεταφραζόμενο mRNA και tRNA συνδεδεμένο με αμινοξέα. -Η πρωτεϊνοσύνθεση γίνεται με ταχύτατο ρυθμό ( πεπτιδικοί δεσμοί ανά λεπτό) σε τρία στάδια: α) έναρξη, β) επιμήκυνση της πεπτιδικής αλυσίδας και γ) τερματισμός..")
26
Σχηματική παράσταση του πλήρους βακτηριακού ριβοσώματος κατά την πρωτεϊνοσύνθεση

27
Αμινογλυκοσίδες Αμινοκυκλιτόλη (συνήθως δεοξυ-στρεπταμίνη) ενωμένη με 2 αμινο-σάκχαρα
 ενωμένη με 2 αμινο-σάκχαρα")
28
Αμινογλυκοσίδες - Είναι μεταβολικά παράγωγα ειδών Streptomyces και Micromonospora -Δεν επηρεάζουν το σχηματισμό του εναρκτήριου συμπλέγματος -Δεσμεύονται στη θέση Α της 30S υπομονάδας του ριβοσώματος: α] αναστολή μετάθεσης από Α σε Ρ β] μεταφραστικά σφάλματα -Σε μερικά Gram-αρνητικά προκαλούν βλάβες στην εξωτερική μεμβράνη -Το τελικό αποτέλεσμα είναι βακτηριοκτόνο -ΔΡΟΥΝ ΜΟΝΟ ΟΤΑΝ ΛΕΙΤΟΥΡΓΕΙ Η ΑΕΡΟΒΙΑ ΑΝΑΠΝΟΗ
![Αμινογλυκοσίδες - Είναι μεταβολικά παράγωγα ειδών Streptomyces και Micromonospora -Δεν επηρεάζουν το σχηματισμό του εναρκτήριου συμπλέγματος -Δεσμεύονται στη θέση Α της 30S υπομονάδας του ριβοσώματος: α] αναστολή μετάθεσης από Α σε Ρ β] μεταφραστικά σφάλματα -Σε μερικά Gram-αρνητικά προκαλούν βλάβες στην εξωτερική μεμβράνη -Το τελικό αποτέλεσμα είναι βακτηριοκτόνο -ΔΡΟΥΝ ΜΟΝΟ ΟΤΑΝ ΛΕΙΤΟΥΡΓΕΙ Η ΑΕΡΟΒΙΑ ΑΝΑΠΝΟΗ](http://images.slideplayer.gr/46/11667517/slides/slide_28.jpg "Αμινογλυκοσίδες - Είναι μεταβολικά παράγωγα ειδών Streptomyces και Micromonospora -Δεν επηρεάζουν το σχηματισμό του εναρκτήριου συμπλέγματος -Δεσμεύονται στη θέση Α της 30S υπομονάδας του ριβοσώματος: α] αναστολή μετάθεσης από Α σε Ρ β] μεταφραστικά σφάλματα -Σε μερικά Gram-αρνητικά προκαλούν βλάβες στην εξωτερική μεμβράνη -Το τελικό αποτέλεσμα είναι βακτηριοκτόνο -ΔΡΟΥΝ ΜΟΝΟ ΟΤΑΝ ΛΕΙΤΟΥΡΓΕΙ Η ΑΕΡΟΒΙΑ ΑΝΑΠΝΟΗ")
29
Τα αντιβιοτικά που αναστέλλουν την πρωτεϊνοσύνθεση δεν αποτελούν μιά ομοιογενή ομάδα

31
Τα αντιβιοτικά που αναφέραμε μέχρι τώρα είναι μεταβολικά προϊόντα μικροβίων του περιβάλλοντος ΚΑΝΤΕ ΥΠΟΘΕΣΕΙΣ ΓΙΑ ΤΙΣ ΑΙΤΙΕΣ

32
Τοπολογία του βακτηριακού χρωμοσώματος Η DNA γυράση καταλύει την υπερέλιξη της διπλής έλικας. Αποτελείται από 2 υπομονάδες GyrA και δύο GyrB. Η τοποϊσομεράση IV καταλύει το διαχωρισμό των χρωμοσωμάτων κατά τον αναδιπλασιασμό. Αποτελείται από 2 υπομονάδες ParC και 2 ParE.

33
Κινολόνες -Δεσμεύονται στα συμπλέγματα DNA-γυράσης και DNA- τοποϊσομεράσης IV και αναστέλλουν τη δράση των ενζύμων

34
Σουλφοναμίδη-Τριμεθοπρίμη Αναστολή της παραγωγής φυλικού

35
Γενικές επισημάνσεις -Τα αντιβιοτικά δεν ‘προκαλούν’ αντοχή. Απλώς αλλάζουν το οικοσύστημα -Η αντοχή στα αντιβιοτικά δεν είναι ένα αλλά πολλαπλά και ετερογενή βιολογικά γεγονότα (π.χ. Κινολόνες vs. Γλυκοπεπτίδια) ΑΝΤΟΧΗ ΣΤΑ ΑΝΤΙΒΙΟΤΙΚΑ
 ΑΝΤΟΧΗ ΣΤΑ ΑΝΤΙΒΙΟΤΙΚΑ.")
36
-Η αντοχή δεν είναι βιολογικό μέγεθος απόλυτο αλλά ΣΧΕΤΙΚΟ που αντανακλά την ικανότητά μας να θεραπεύουμε τις λοιμώξεις α) Φαρμακολογικός καθορισμός των ορίων β) Προσαρμογές των ορίων (π.χ. ESBL πρίν και μετά το 1997, MRSA και β-λακτάμες, Σαλμονέλλες και κινολόνες) Γενικές επισημάνσεις
 Γενικές επισημάνσεις.")
37
-Η εμφάνιση και η διασπορά της αντοχής δεν μπορούν να προβλεφθούν (π.χ. S. pyogenes-πενικιλλίνες, διασπορά CTX-M) -Η σχέση της χρήσης αντιβιοτικών και της συχνότητας αντοχής δεν είναι γραμμική (π.χ. αντοχή στη χλωραμφαινικόλη) Γενικές επισημάνσεις
 -Η σχέση της χρήσης αντιβιοτικών και της συχνότητας αντοχής δεν είναι γραμμική (π.χ. αντοχή στη χλωραμφαινικόλη) Γενικές επισημάνσεις.")
38
ΕΓΓΕΝΗΣ ΑΝΤΟΧΗ Παραδείγματα εγγενούς αντοχής -Γλυκοπεπτίδια vs Gram-αρνητικά -Μακρολίδες vs Gram-αρνητικά Αμινογλυκοσίδες vs αναερόβια -Aztreonam vs Staphylococcus aureus -Κεφοταξίμη vs Pseudomonas aeruginosa -Ημισυνθετικές πενικιλλίνες vs Klebsiella pneumoniae -Co-trimoxazole vs Pseudomonas aeruginosa -Τετρακυκλίνες vs Proteus mirabilis Στη Κλινική Μικροβιολογία ο όρος αντοχή σημαίνει ΕΠΙΚΤΗΤΗ αντοχή

39
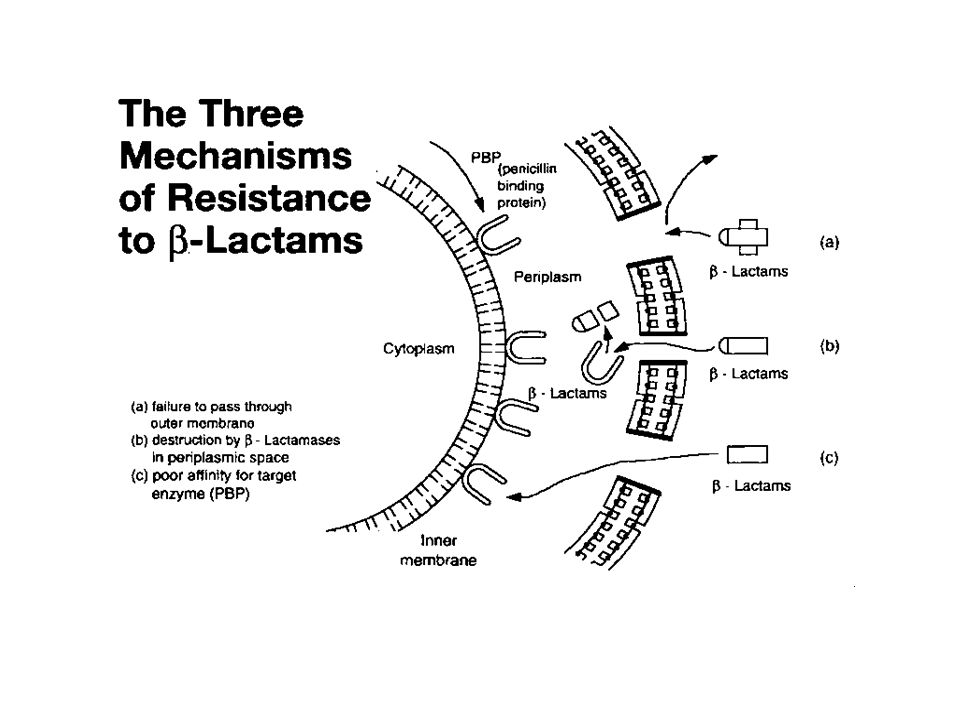
ΣΥΝΗΘΗΣ ΛΕΙΤΟΥΡΓΙΚΗ ΚΑΤΑΤΑΞΗ ΜΗΧΑΝΙΣΜΩΝ ΑΝΤΟΧΗΣ 1. Αδρανοποίηση αντιβιοτικού (κυρίως αδρανοποιητικά ένζυμα) 2. Αλλαγή του στόχου του αντιβιοτικού (πληθώρα τύπων αντοχής - αλλαγές στο γενετικό υλικό) 3. Μείωση της ενδοκυττάριας συγκέντρωσης του αντιβιοτικού (κυρίως συστήματα αποτοξίνωσης και μείωση διαπερατότητας εξ. μεμβράνης)
 2. Αλλαγή του στόχου του αντιβιοτικού (πληθώρα τύπων αντοχής - αλλαγές στο γενετικό υλικό) 3. Μείωση της ενδοκυττάριας συγκέντρωσης του αντιβιοτικού (κυρίως συστήματα αποτοξίνωσης και μείωση διαπερατότητας εξ. μεμβράνης).")
40
Αδρανοποίηση αντιβιοτικούΠαραδείγματα -β-Λακταμάσες (υδρολάσες) -Αμινογλυκοσιδο-τροποποιητικά ένζυμα (τρανσφεράσες) -Ακετυλο-τρανσφεράσες της χλωραμφαινικόλης -Εστεράσες των μακρολιδών
 -Αμινογλυκοσιδο-τροποποιητικά ένζυμα (τρανσφεράσες) -Ακετυλο-τρανσφεράσες της χλωραμφαινικόλης -Εστεράσες των μακρολιδών")
41
ΠΡΟΕΛΕΥΣΗ ΜΗΧΑΝΙΣΜΩΝ ΑΝΤΟΧΗΣ [A] ΕΤΟΙΜΟΙ ΠΡΟΣ ΧΡΗΣΗ ΜΗΧΑΝΙΣΜΟΙ [B] ΣΗΜΕΙΑΚΕΣ ΜΕΤΑΛΛΑΞΕΙΣ Γ] ΑΝΑΚΑΤΑΤΑΞΕΙΣ ΓΟΝΙΔΙΩΝ (μωσαϊκισμοί)
![ΠΡΟΕΛΕΥΣΗ ΜΗΧΑΝΙΣΜΩΝ ΑΝΤΟΧΗΣ [A] ΕΤΟΙΜΟΙ ΠΡΟΣ ΧΡΗΣΗ ΜΗΧΑΝΙΣΜΟΙ [B] ΣΗΜΕΙΑΚΕΣ ΜΕΤΑΛΛΑΞΕΙΣ Γ] ΑΝΑΚΑΤΑΤΑΞΕΙΣ ΓΟΝΙΔΙΩΝ (μωσαϊκισμοί)](http://images.slideplayer.gr/46/11667517/slides/slide_41.jpg "ΠΡΟΕΛΕΥΣΗ ΜΗΧΑΝΙΣΜΩΝ ΑΝΤΟΧΗΣ [A] ΕΤΟΙΜΟΙ ΠΡΟΣ ΧΡΗΣΗ ΜΗΧΑΝΙΣΜΟΙ [B] ΣΗΜΕΙΑΚΕΣ ΜΕΤΑΛΛΑΞΕΙΣ Γ] ΑΝΑΚΑΤΑΤΑΞΕΙΣ ΓΟΝΙΔΙΩΝ (μωσαϊκισμοί)")
42
Οι περισσότεροι μηχανισμοί αντοχής είναι αδύνατον να προκύψουν de novo (μεταλλάξεις + πίεση επιλογής) Παράδειγμα: Η αντοχή των εντεροκόκκων στη βανκομυκίνη -Το VanH ανάγει το πυροσταφυλικό σε D-γαλακτικό. -Η λιγάση VanA ενώνει το D-γαλακτικό με D-αλανίνη. -To VanX υδρολύει D-Ala-D-Ala αλλά όχι D-Ala-D-Lac. -Τα προϊόντα των γονιδίων vanS και vanR ρυθμίζουν τη παραγωγή VanH, A και X.

43
Γνωρίζοντας ότι τα περισσότερα αντιβιοτικά είναι μικροβιακοί μεταβολίτες ή ημισυνθετικά παράγωγα μεταβολιτών VAN ΠΡΟΤΕΙΝΕΤΕ ΠΙΘΑΝΕΣ ΠΗΓΕΣ ΠΡΟΕΛΕΥΣΗΣ ΤΗΣ ΑΝΤΟΧΗΣ VAN

44
-Αρκετοί μηχανισμοί αντοχήςπιθανόν προέρχονται απο είδη που παράγουν τα αντίστοιχα αντιβιοτικά (ΑΥΤΟΠΡΟΣΤΑΣΙΑ)
")
45
Υπάρχουν ‘έτοιμοι προς χρήση’ μηχανισμοί ο φυσιολογικός ρόλος των οποίων δεν σχετίζεται με τα αντιβιοτικά π.χ. -Η PBP2a του MRSA - Οι κεφαλοσπορινάσες των εντεροβακτηριακών - Τα αμινογλυκοσιδο-τροποποιητικά ένζυμα

46
Σε μερικές περιπτώσεις η αντοχή προκαλείται από πλειοτροπικές μεταλλάξεις π.χ. Μειωμένη ευαισθησία του S. aureus στη βανκομυκίνη -Περισσότερες της μιας μεταλλάξεις. -Προβλήματα στη παραγωγή πεπτιδογλυκάνης -Συσσώρευση μεταβολιτών που “παγιδεύουν” το αντιβιοτικό

47
Οι “έτοιμοι προς χρήση” μηχανισμοί μπορούν να εξελιχθούν συνδυαζόμενοι με σημειακές μεταλλάξεις Π.Χ. Η ανάπτυξη ESBL από τις κλασσικές πενικιλλινάσες ΤΕΜ και SHV

48
ΠΝΕΥΜΟΝΙΟΚΟΚΚΟΙΜΗΝΙΓΓΙΤΙΔΟΚΟΚΚΟΙ -Μετακίνηση κομματιών DNA από άλλα είδη με τη διαδικασία της ΜΕΤΑΜΟΡΦΩΣΗΣ (χωρίς φορέα) Προκύπτουν “μωσαϊκά” PBPs που είναι ανθεκτικά στις β-λακτάμες
 Προκύπτουν μωσαϊκά PBPs που είναι ανθεκτικά στις β-λακτάμες")
49
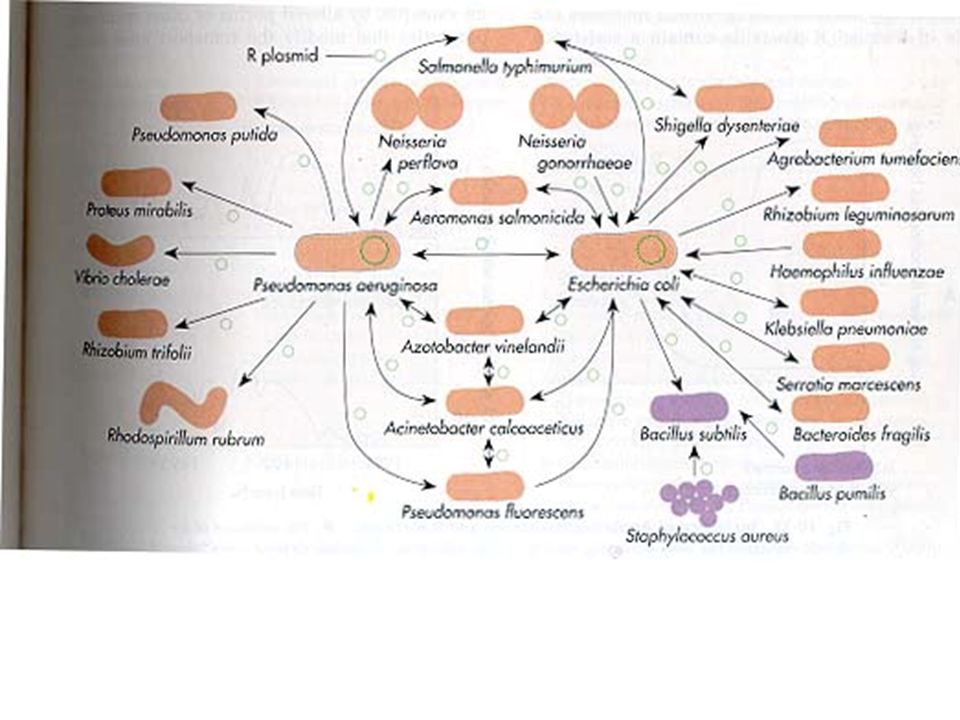
ΜΕΤΑΦΟΡΑ ΜΗΧΑΝΙΣΜΩΝ ΑΝΤΟΧΗΣ Το γενετικό υλικό που κωδικοποιεί αντοχή (γονίδια -συστήματα γονιδίων) έχει τη δυνατότητα να διασπείρεται μεταξύ διαφορετικών ειδών
 έχει τη δυνατότητα να διασπείρεται μεταξύ διαφορετικών ειδών")
51
Για ποιό λόγο γίνονται αυτές οι μετακινήσεις ? -Η μετακίνήση γενετικού υλικού που κωδικοποιεί αντοχή δεν απαιτεί παρουσία αντιβιοτικού. -Πρόκειται για αυτόματες βιολογικές διαδικασίες στα πλαίσια της διατήρησης της γενετικής ποικιλότητας. Υπάρχουν φραγμοί στη μετακίνηση ? -Τα φαινόμενα υποδεικνύουν την ύπαρξη φραγμών (π.χ. Απουσία β-λακταμασών από το S. pyogenes). -Η πειραματική προσέγγιση είναι δύσκολη.
. -Η πειραματική προσέγγιση είναι δύσκολη..")
52
Πως γίνεται η μετακίνηση των γονιδίων αντοχής ? Α -Συχνά, τα γονίδια αντοχής φέρονται από κινητά στοιχεία (π.χ. πλασμίδια, μεταθετά [transposons]) -Τα στοιχεία αυτά μεταβάλλονται αυτόματα (παίρνοντας ή χάνοντας γονίδια αντοχής) B -Σε λίγες (αλλά κλινικά σημαντικές) περιπτώσεις η μετακίνηση γονιδίων αντοχής γίνεται χωρίς τη βοήθεια φορέα
 -Τα στοιχεία αυτά μεταβάλλονται αυτόματα (παίρνοντας ή χάνοντας γονίδια αντοχής) B -Σε λίγες (αλλά κλινικά σημαντικές) περιπτώσεις η μετακίνηση γονιδίων αντοχής γίνεται χωρίς τη βοήθεια φορέα.")
53
ΠΛΑΣΜΙΔΙΑ - ΟΧΗΜΑΤΑ ΜΕΤΑΦΟΡΑΣ ΑΝΤΟΧΗΣ -Τα πλασμίδια είναι φορείς πολλών σημαντικών μηχανισμών αντοχής (β-Λακταμάσες, Τροποποιητικά αμινογλυκοσιδών, CAT, TET DHFR κλπ). -Πολλά πλασμίδια αντοχής έχουν τη δυνατότητα αυτομεταφοράς.

54
Η μεταφορά (διασπορά) γίνεται με το μηχανισμό της σύζευξης
 γίνεται με το μηχανισμό της σύζευξης")
55
Mεγάλο μέρος ενός πλασμιδίου αντοχής είναι “επίκτητο” (μεταθετά, IS, ιντεγκρόνια). Τα γονίδια αντοχής βρίσκονται στα επίκτητα κομμάτια DNA.

56
Μεταθετά (Transposons) IRi tnpAtnpRtnpM IRt mer operon ITR IS elements ITR L TrasposaseITR R Class I transposons Tet R IS10LIS10R Tn10 Cm R IS1LIS1R Tn9 Class II transposons tnpAtnpRAmp R Res Tn3 ITR In2 Tn21
 IRi tnpAtnpRtnpM IRt mer operon ITR IS elements ITR L TrasposaseITR R Class I transposons Tet R IS10LIS10R Tn10 Cm R IS1LIS1R Tn9 Class II transposons tnpAtnpRAmp R Res Tn3 ITR In2 Tn21")
57
Γενικός τύπος κίνησης μεταθετών στοιχείων

58
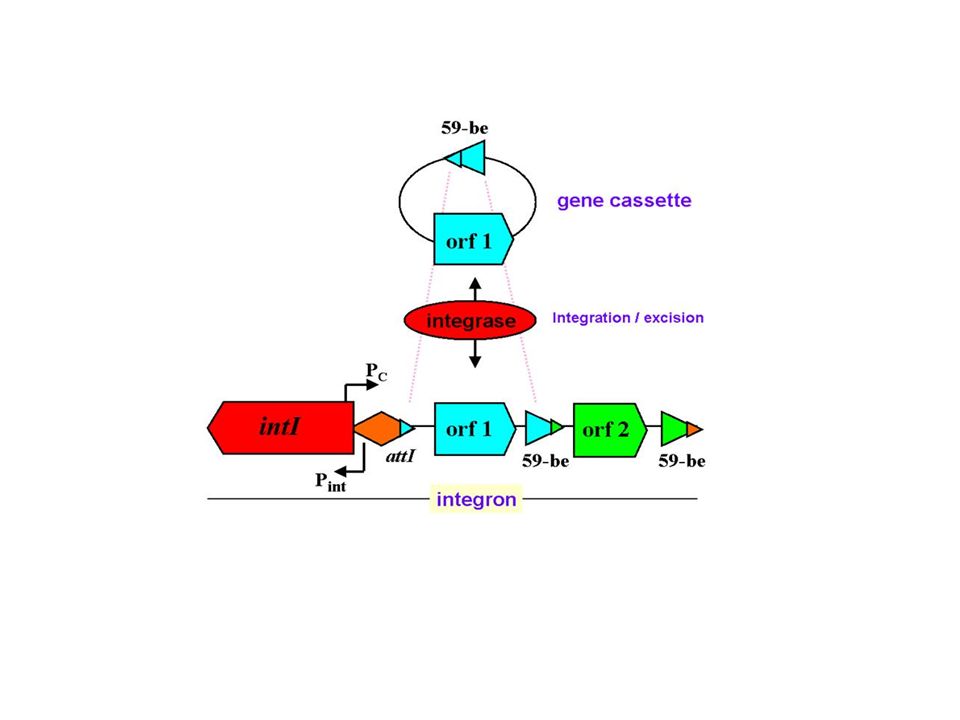
-Τα ιντεγκρόνια είναι συστήματα παγίδευσης γονιδίων αντοχής -Δεν κινούνται αλλά χρησιμοποιούν τα μεταθετά σαν οχήματα

60
In-t1/In-t2 (Tn 1935, IncFI/97) In-t2In-t1 intI1sulIintI1sulI ORF341 tnpA tnpR tnpM dfrA18 oxa1aadA1 aadBcatB3 IRi tnpA tnpR tnpM In-t2 (Tn1935, pZM3) intI1sulI oxa1 aadA1 IRi In2 (Tn21) tnpA tnpR tnpM aadA1 IRi sulIintI1 3G Τα μεταθετά και τα ιντεγκρόνια εξελίσονται
 In-t2In-t1 intI1sulIintI1sulI ORF341 tnpA tnpR tnpM dfrA18 oxa1aadA1 aadBcatB3 IRi tnpA tnpR tnpM In-t2 (Tn1935, pZM3) intI1sulI oxa1 aadA1 IRi In2 (Tn21) tnpA tnpR tnpM aadA1 IRi sulIintI1 3G Τα μεταθετά και τα ιντεγκρόνια εξελίσονται")
61
Δεξαμενή γονιδίων αντοχής Ιντεγκρόνια Γονιδιακές κασέτες Μεταθετά στοιχεία Η αδιάκοπη κίνηση του γενετικού υλικού συνδυαζόμενη με τη πίεση επιλογής οδηγεί στο σχηματισμό… Πλασμίδια αντοχής

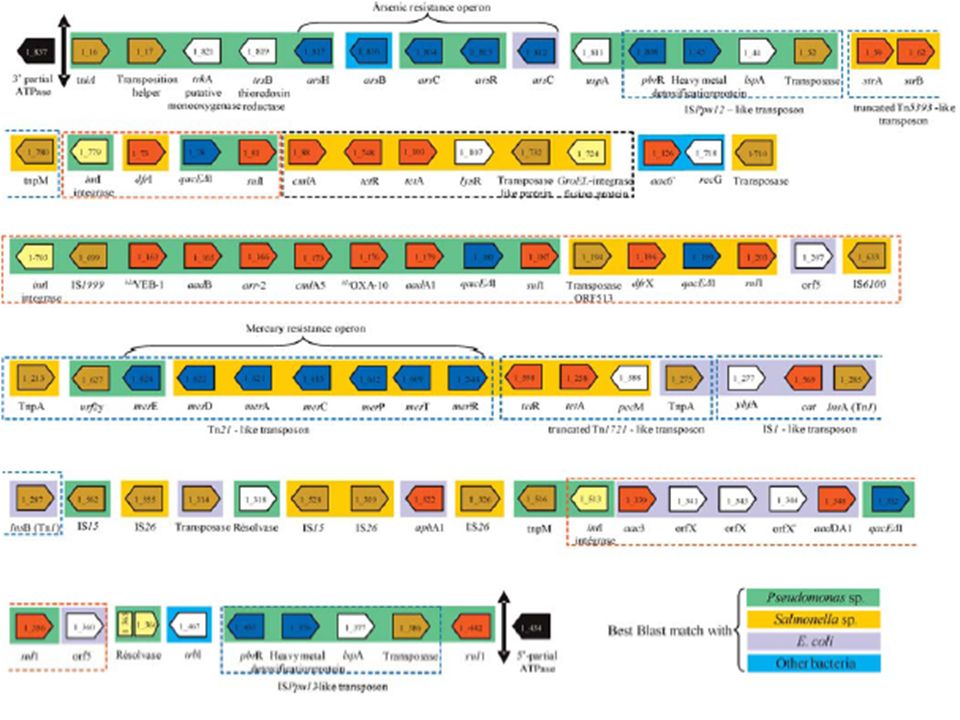
62
pSEM p2815 aacA4intIaadA1qacsulI1IS26-1orfX’orfXaacC1 shv-5ΔlacY aacA4intIaadA1qacsulI1aphAIS26IS26-1dhfrIorfOIBC-1orf5IS6100orf6 IRt orf5IS6100orf6 IRt K. pmeumoniae chromosomal region IS26 ΔygbMfucA ygbKygbJ ygbIrecF p541 VIM-1intIaadA1qacsulI1 IS26-1 dhfrIaacA7 ΔfrdBampRsugE aphAIS26 cmy13 Δorf5 IS1ΔIS26IS6100orf6 IRt IS26Δorf5 C.freundii chromosomal region blcfrdDfrd C TSD Δ5’CS ……..συγκροτημάτων πολυαντοχής……. …όπως αυτά που έχουν βρεθεί σε Gram αρνητικά μικρόβια στα Ελληνικά νοσοκομεία.

63
Αντοχή στα Gram αρνητικά FQ-R P. aeruginosa Ceftaz-R K. pneumoniae FQ-R E. coli

64
Κάντε υποθέσεις γιά τις αιτίες αύξησης της αντοχής

65
Μηχανισμοί αντοχής στα Gram αρνητικά -Βιολογία -Επιδημιολογία -Ταυτοποίηση-Διάγνωση -Αντιμετώπιση

66
Αντοχή στα β-λακταμικά αντιβιοτικά -β-Λακταμάσες -μειωμένη διαπερατότητα -αλλαγές στις ΡΒΡs Κυρίαρχος μηχανισμός στα Gram-αρνητικά είναι η παραγωγή β-λακταμασών ενώ στα Gram-θετικά οι αλλαγές στις ΡΒΡs.

68
Η πρώτη περιγραφή πλασμιδιακής β-λακταμάσης (ΤΕΜ-1) το 1965
 το 1965")
69
ΑΝΤΟΧΗ ΣΤΑ β-ΛΑΚΤΑΜΙΚΑ β-ΛΑΚΤΑΜΑΣΕΣ -Είναι υδρολάσες. Δηλαδή χρησιμοποιούν νερό για να διασπάσουν το β-λακταμικό δακτύλιο. Τα προϊόντα υδρόλυσης είναι αδρανή..

70
-Φυλογενετικά, είναι συγγενείς των PBPs. -O “φυσιολογικός” τους ρόλος δεν έχει προσδιοριστεί

72
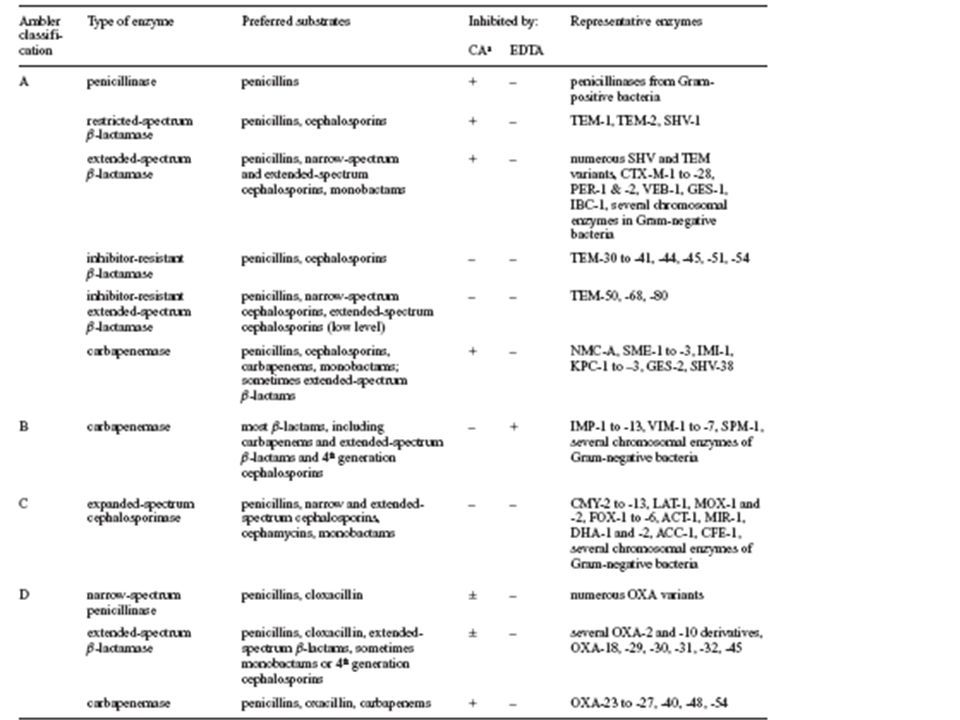
β-ΛΑΚΤΑΜΑΣΕΣ ΤΑΞΗΣ Α -Οι κοινές πλασμιδιακές πενικιλλινάσες έχουν ηλικία >500.000 ετών. -Η εντατική χρήση β-λακταμών προκάλεσε την ευρύτατη διασπορά των αντίστοιχων γονιδίων. -Η διασπορά έγινε (και γίνεται) με ποικιλία μηχανισμών -Από τις κοινές πενικιλλινάσες προέκυψαν πρόσφατα οι εκτεταμένου φάσματος β-λακταμάσες (ESBL).
 με ποικιλία μηχανισμών -Από τις κοινές πενικιλλινάσες προέκυψαν πρόσφατα οι εκτεταμένου φάσματος β-λακταμάσες (ESBL)..")
73
-Οι ΤΕΜ-1, -2 και SHV-1 β-λακταμάσες προκαλούν αντοχή στις πενικιλλίνες και στις κεφαλοσπορίνες 1ης γενιάς -Αντοχή στους συνδυασμούς πενικιλλινών/αναστολέων παρατηρείται σε περιπτώσεις υπερπαραγωγής

74
Υποκαταστάσεις αμινοξέων στις ΕSBL

75
Οι ΕSBL υποκαταστάσεις δεν έχουν άμεση σχέση με το αντιβιοτικό που υδρολύεται

76
Οι κοινές πενικιλλινάσες μετατρέπονται σε ESBL με σημειακές μεταλλάξεις που διερύνουν το ενεργό κέντρο (ετσι ώστε να “χωράει” την οξυιμινο-ομάδα των νεότερων κεφαλοσπορινών) R1 Παλαιές κεφαλοσπορίνες Νεότερες κεφαλοσπορίνες
 R1 Παλαιές κεφαλοσπορίνες Νεότερες κεφαλοσπορίνες")
77
CTX-M β-λακταμάσες -Δομικά και λειτουργικά μοιάζουν με τις κλασσικές ESBL -Προέρχονται από είδη Kluyvera (κινητοποίηση σε πλασμίδια με φορείς μεταθετά στοιχεία -Υδρολύουν κατά προτίμηση κεφοταξίμη-κεφτριαξόνη -Εχουν διασπαρεί στη κοινότητα (?!)
")
78
Σημειακές μεταλλάξεις στις CTX-M Asp240Gly, Pro167Ser Arg164His, Asp179Gly, Αrg276Ser Οι μεταλλάξεις αυτές βελτιώνουν τη δράση των CTX-M κατά της κεφταζιδίμης είτε αυξάνοντας τη συνάφεια (μικρότερα Km) είτε αυξάνοντας τη ταχύτητα υδρόλυσης
 είτε αυξάνοντας τη ταχύτητα υδρόλυσης")
79
-Χαρακτηριστικό των ESBL είναι η υπερευαισθησία στους αναστολείς (Clavulanic acid, Tazobactam). -Η ιδιότητα αυτή έχει διαγνωστική σημασία. Αναγνώριση ESBL-θετικών στελεχών

80
Ανίχνευση ESBL-θετικών στο εργαστήριο Υπάρχει ποικιλία μεθόδων (Ε-test, μέθοδοι βασισμένες σε δίσκους, αυτόματα συστήματα). Ολες είναι αποδεκτές. Προσοχή στα παρακάτω: - Οι μέθοδοι είναι προτυποποιημένες μόνο για K. pneumoniae και E. coli. -H Κ. oxytoca είναι ιδιαίτερα προβληματική. -Τα αυτόματα συστήματα που βασίζονται στη φαινοτυπική διάκριση πιθανόν επιδέχονται βελτιώσεις. -Κυκλοφορούν νέες ESBL που δεν είναι τόσο ευαίσθητες στο κλαβουλανικό όσο οι κλασσικές.

81
Θεραπεία λοιμώξεων από ESBL-θετικά μικρόβια Προτεινόμενες θεραπευτικές λύσεις: -Καρβαπενέμες, Κινολόνες, αμινογλυκοσίδες. -Εξακολουθούν να συζητούνται οι κεφαλοσπορίνες 4ης γενιάς και οι συνδυασμοί πενικιλλινών-αναστολέων.

82
ΚΑΡΒΑΠΕΝΕΜΑΣΕΣ ΤΑΞΗΣ Α Τύποι: GES/IBC, KPC, SME, NMC, IMI Υδρολυτικός μηχανισμός: Πυρηνόφιλη προσβολή του β-λακταμικού δακτυλίου της καρβαπενέμης από υδροξύλιο σερίνης. Η δομή τους Α έχει αρκετές ομοιότητες με εκείνη των συνηθισμένων πενικιλλινασών (50-60%)
.")
83
-Χαρακτηριστικό των καρβαπενεμασών της τάξης Α είναι η παρουσία γέφυρας S στο ενεργό κέντρο -Η υδρόλυση των καρβαπενεμών είναι πολύ αποτελεσματική π.χ. Sme-1 ΑμπικιλλίνηΙμιπενέμη Kcat 58 /sec185/sec Km0,5 mM0.5 mM -Το φάσμα υποστρωμάτων ποικίλει ως πρός τις νεώτερες κεφαλοσπορίνες -Αναστέλλονται από κλαβουλανικό και ταζομπακτάμη (λιγότερο από τις συνήθεις ESBL)
.")
84
-Οι καρβαπενεμάσες της τάξης Α παράγονται κυρίως από εντεροβακτηριακά bla -Τα αντίστοιχα bla γονίδια συνήθως μεταφέρονται από πλασμίδια -Η προέλευση τους είναι άγνωστη Τα γονίδια GES φέρονται από ιντεγκρόνια πολλαπλής αντοχής Τα KPC “συνδυάζονται” με αλληλουχίες από περιβαλλοντικά και φυτοπαθογόνα μικρόβια (γένη Pseudomonas και Ralstonia).
.")
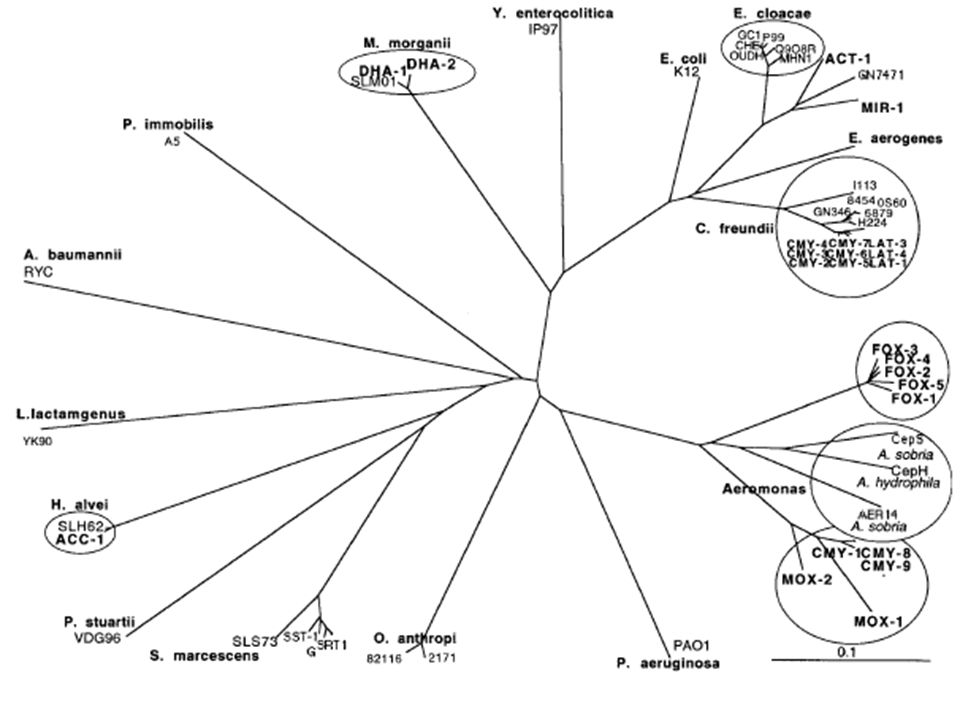
85
ΚΕΦΑΛΟΣΠΟΡΙΝΑΣΕΣ (ΤΑΞΗ C) Πολλά κλινικώς σημαντικά είδη παράγουν “κεφαλοσπορινάσες” Τα αντίστοιχα γονίδια (ampC) είναι χρωμοσωματικής προέλευσης Ο φυσιολογικός ρόλος τους δεν είναι γνωστός (πιθανόν συμμετέχουν στο μεταβολισμό της πεπτιδογλυκάνης Η παραγωγή τους είναι συνήθως επαγώγιμη Μερικές κεφαλοσπορινάσες έχουν μεταπηδήσει σε πλασμίδια
 Πολλά κλινικώς σημαντικά είδη παράγουν κεφαλοσπορινάσες Τα αντίστοιχα γονίδια (ampC) είναι χρωμοσωματικής προέλευσης Ο φυσιολογικός ρόλος τους δεν είναι γνωστός (πιθανόν συμμετέχουν στο μεταβολισμό της πεπτιδογλυκάνης Η παραγωγή τους είναι συνήθως επαγώγιμη Μερικές κεφαλοσπορινάσες έχουν μεταπηδήσει σε πλασμίδια")
87
Επαγωγή κεφαλοσπορινασών

88
Μόνιμη αποκαταστολή προκαλείται από μεταλλάξεις στο γονίδιο D Επιλογή στελεχών με αποκαταστολή Γίνεται από β-λακταμικά που α] υδρολύονται από κεφαλοσπορινάσες β] δεν είναι επαγωγείς ΔΗΛΑΔΗ ΑΠΟ ΚΕΦΑΛΟΣΠΟΡΙΝΕΣ ΤΡΙΤΗΣ ΓΕΝΙΑΣ
![Μόνιμη αποκαταστολή προκαλείται από μεταλλάξεις στο γονίδιο D Επιλογή στελεχών με αποκαταστολή Γίνεται από β-λακταμικά που α] υδρολύονται από κεφαλοσπορινάσες β] δεν είναι επαγωγείς ΔΗΛΑΔΗ ΑΠΟ ΚΕΦΑΛΟΣΠΟΡΙΝΕΣ ΤΡΙΤΗΣ ΓΕΝΙΑΣ](http://images.slideplayer.gr/46/11667517/slides/slide_88.jpg "Μόνιμη αποκαταστολή προκαλείται από μεταλλάξεις στο γονίδιο D Επιλογή στελεχών με αποκαταστολή Γίνεται από β-λακταμικά που α] υδρολύονται από κεφαλοσπορινάσες β] δεν είναι επαγωγείς ΔΗΛΑΔΗ ΑΠΟ ΚΕΦΑΛΟΣΠΟΡΙΝΕΣ ΤΡΙΤΗΣ ΓΕΝΙΑΣ")
89
ΚΙΝΗΤΟΠΟΙΗΣΗ ΚΕΦΑΛΟΣΠΟΡΙΝΑΣΩΝ

90
orf1 pCMY plasmids tnpA CMYblc SugE ISEcp1 orf6orf7 tnpA CMYblc SugE ISEcp1 IS26 ecnR C. freundii-χρωμοσωματική περιοχή ampR CMYblc SugE ecnR Fumarate operon CMY Μετάθεση κεφαλοσπορινασών με ISEcp1

91
Ενσωμάτωση κεφαλοσπορινασών σε ιντεγκρόνια

92
Φαινότυποι κεφαλοσπορινασών Αντοχή ή μειωμένη ευαισθησία σε όλα τα β-λακταμικά εκτός καρβαπενεμών και 4ης γενιάς κεφαλοσπορινών Χαρακτηριστική αντοχή σε κεφοξιτίνη και πενικιλλίνες-αναστολείς (διάκριση από ESBL)
")
93
ΟΧΑ β-λακταμάσες (τάξη D) -ΟΧΑ πενικιλλινάσες (ευρεία διασπορά στα παθογόνα Gram-αρνητικά) -ΟΧΑ εκτεταμένου φάσματος (σπάνια, κυρίως σε Pseudomonas aeruginosa) -ΟΧΑ καρβαπενεμάσες (κυρίως σε Acinetobacter) Εχουν περιγραφεί 120. Οι περισσότερες (70) είναι πενικιλλινάσες. Τα γονίδια ΟΧΑ βρίσκονται συνήθως σε ιντεγκρόνια (πλασμίδια και χρομωσώματα)
 είναι πενικιλλινάσες. Τα γονίδια ΟΧΑ βρίσκονται συνήθως σε ιντεγκρόνια (πλασμίδια και χρομωσώματα).")
94
-Ο μηχανισμός υδρόλυσης είναι παρόμοιος με αυτόν στη τάξη Α (προσβολή του β-λακταμικού δακτυλίου από υδροξύλιο σερίνης). -Τα ένζυμα ΟΧΑ λειτουργούν ως διμερή -Αναστέλλονται από ιόντα χλωρίου -Τα περισσότερα είναι ανθεκτικά στο κλαβουλανικό

95
Καρβαπενεμάσες τάξης D (OXA β-λακταμάσες) Εχουν περιγραφεί 120 ποικιλίες ΟΧΑ. Από αυτές, οι 40 έχουν δράση «καρβαπενεμάσης» -Βρίσκονται κυρίως σε Acinetobacter -Δεν προέρχονται από τις OXA πενικιλλινάσες -Η πιθανή πηγή τους είναι περιβαλλοντικά μικρόβια (είδη Shewanella)
.")
96
-Οι καρβαπενεμάσες ΟΧΑ υδρολύουν πενικιλλίνες, κεφαλοσπορίνες 1ης γενιάς και καρβαπενέμες -Η ταχύτητα υδρόλυσης της ιμιπενέμης είναι χαμηλή Αυτό αντισταθμίζεται μερικώς από τη σχετικά μεγάλη συνάφεια με το υπόστρωμα ΟΧΑ-58 ΠενικιλλίνηΙμιπενέμη Kcat 5,5/sec0.1/sec Km0.05mM0.07mM -Οι καρβαπενεμάσες τύπου ΟΧΑ υποτίθεται ότι συνεισφέρουν στην αντοχή των Acinetobacter στις καρβαπενέμες. Ωστόσο, είναι αμφίβολης κλινικής σημασίας επειδή : α) δεν είναι απαραίτητες γιά την ανάπτυξη αντοχής β) συνυπάρχουν πάντα με άλλους μηχανισμούς αντοχής γ) η εξουδετέρωσή τους δεν μειώνει σημαντικά το επίπεδο αντοχής -Στα Ελληνικά νοσοκομεία έχουν περιγραφεί πολυανθεκτικά Acinetobacter με ΟΧΑ-58.
 δεν είναι απαραίτητες γιά την ανάπτυξη αντοχής β) συνυπάρχουν πάντα με άλλους μηχανισμούς αντοχής γ) η εξουδετέρωσή τους δεν μειώνει σημαντικά το επίπεδο αντοχής -Στα Ελληνικά νοσοκομεία έχουν περιγραφεί πολυανθεκτικά Acinetobacter με ΟΧΑ-58..")
97
ΜΕΤΑΛΛΟ-β-ΛΑΚΤΑΜΑΣΕΣ (τάξη Β)
")
98
ΜΕΤΑΛΛΟ-β-ΛΑΚΤΑΜΑΣΕΣ (τάξη Β) -Τρείς υποτάξεις B1 B1: χρωμοσωματικά ένζυμα από Bacillus, Bacteroides, Chryseobacterium καθώς και τα ένζυμα VIM και ΙΜP B2 B2 : χρωμοσωματικές β-λακταμάσες από Αeromonas B3 B3 : το ένζυμο L1 της Stenotrophomonas maltophilia Δεν έχουν σχέση με τα ένζυμα που συνθέτουν πεπτιδογλυκάνη Παρουσιάζουν ομοιότητες με βακτηριακές υδρολάσες Ιδιαίτερης κλινικής σημασίας είναι οι VIM και ΙΜP τύποι Τα αντίστοιχα γονίδια έχουν διασπαρεί σε πολλά Gram αρνητικά (Pseudomonas, Acinetobacter, Achromobacter και εντεροβακτήρια).
 -Τρείς υποτάξεις B1 B1: χρωμοσωματικά ένζυμα από Bacillus, Bacteroides, Chryseobacterium καθώς και τα ένζυμα VIM και ΙΜP B2 B2 : χρωμοσωματικές β-λακταμάσες από Αeromonas B3 B3 : το ένζυμο L1 της Stenotrophomonas maltophilia Δεν έχουν σχέση με τα ένζυμα που συνθέτουν πεπτιδογλυκάνη Παρουσιάζουν ομοιότητες με βακτηριακές υδρολάσες Ιδιαίτερης κλινικής σημασίας είναι οι VIM και ΙΜP τύποι Τα αντίστοιχα γονίδια έχουν διασπαρεί σε πολλά Gram αρνητικά (Pseudomonas, Acinetobacter, Achromobacter και εντεροβακτήρια).")
99
Η προέλευση των IMP και VIM β-λακταμασών είναι άγνωστη

100
-Τα ένζυμα αυτά χρειάζονται ιόντα ψευδαργύρου (α ναστέλλονται από EDTA) -Το φάσμα υποστρωμάτων περιλαμβάνει όλα τα β-λακταμικά (εκτός Αztreonam)
 -Το φάσμα υποστρωμάτων περιλαμβάνει όλα τα β-λακταμικά (εκτός Αztreonam)")
101
bla VIM-1 aacA7dhfrIaadAintIsulIqacEΔ1 bla VIM-1 aacA4intIsulIqacEΔ1repeat bla VIM-1 aac(6’)-IIcintIsulIqacEΔ1 bla VIM-2 intIsulIqacEΔ1 bla VIM-2 aacA29aintIsulIqacEΔ1 bla VIM-12 aacA7intIsulIqacEΔ1 aacA7 bla VIM-4 aac(6’)orfDaadAintIsulIqacEΔ1 bla VIM-4 intIsulIqacEΔ1 P. aeruginosa E. cloacae E. coli K. pneumoniae E. coli A. baumannii P. mirabilis P. aeruginosa K. pneumoniae Τα γονίδια VIM (και ΙΜP) φέρονται από ιντεγκρόνια Υπάρχει ποικιλία VIM ιντεγκρονίων στα μικρόβια των Ελληνικών νοσοκομείων Tsakris et al., 2000; Mavroidi et al., 2000; Pournaras et al., 2002; Pournaras et al., 2003; Miriagou et al., 2003; Giakkoupi et al., 2003a & 2003b; Scoulica et al., 2004; Galani et al., 2005; Ikonomidis et al., 2005; GenBank.
 φέρονται από ιντεγκρόνια Υπάρχει ποικιλία VIM ιντεγκρονίων στα μικρόβια των Ελληνικών νοσοκομείων Tsakris et al., 2000; Mavroidi et al., 2000; Pournaras et al., 2002; Pournaras et al., 2003; Miriagou et al., 2003; Giakkoupi et al., 2003a & 2003b; Scoulica et al., 2004; Galani et al., 2005; Ikonomidis et al., 2005; GenBank..")
102
Διαγνωστικά και θεραπευτικά ζητήματα -Θεωρητικά, τα μικρόβια που παράγουν VIM τύπου β-λακταμάσες ανιχνεύονται εύκολα (φαινότυπος + αναστολή από EDTA) -Ωστόσο, έχουν προκύψει σημαντικά διαγνωστικά προβλήματα α) αναντιστοιχίες μεθόδων - χαμηλή επαναληψιμότητα β) χαμηλά MIC στις καρβαπενέμες γ) λάθη στην ερμηνεία από τα “expert systems”
 -Ωστόσο, έχουν προκύψει σημαντικά διαγνωστικά προβλήματα α) αναντιστοιχίες μεθόδων - χαμηλή επαναληψιμότητα β) χαμηλά MIC στις καρβαπενέμες γ) λάθη στην ερμηνεία από τα expert systems")
103
ΠΡΟΒΛΗΜΑΤΑ ΣΤΗ ΘΕΡΑΠΕΥΤΙΚΗ ΑΝΤΙΜΕΤΩΠΙΣΗ -Τι ευαισθησία πρέπει να αναφέρει το εργαστήριο αν τα MICs είναι <16mg/L ??? -Ποιά αντιβιοτικά είναι αποτελεσματικά ??? -Σύμφωνα με τα in vitro δεδομένα, θα μπορούσαν να χρησιμοποιηθούν : Κολιστίνη (βακτηριαιμία ?, λοιμώξεις αναπνευστικού ?) Αztreonam Κινολόνες Αμινογλυκοσίδες -Τα κλινικά δεδομένα δεν είναι επαρκή (μέχρι τώρα)
 Αztreonam Κινολόνες Αμινογλυκοσίδες -Τα κλινικά δεδομένα δεν είναι επαρκή (μέχρι τώρα).")
104
Ενα κλινικό στέλεχος K. pneumoniae είναι ανθεκτικό στη κεφοξιτίνη. Ποιούς τύπους β-λακταμασών υποπτεύεστε ?

105
ΑΜΙΝΟΓΛΥΚΟΣΙΔO-ΤΡΟΠΟΠΟΙHTIKA ΕΝΖΥΜΑ (ATE) Yπάρχουν τρείς οικογένειες ενζύμων 1- Νουκλεοτιδυλο-τρανσφεράσες (ΑΝΤ) 2-Φωσφο-τρανσφεράσες (ΑΡΗ) 3-Ακετυλο-τρανσφεράσες (ΑΑC)
 Yπάρχουν τρείς οικογένειες ενζύμων 1- Νουκλεοτιδυλο-τρανσφεράσες (ΑΝΤ) 2-Φωσφο-τρανσφεράσες (ΑΡΗ) 3-Ακετυλο-τρανσφεράσες (ΑΑC)")
106
—N-ακετυλοτρανσφεράσες (AAC). Τροποποιούν αμινομάδες. Χρησιμοποιούν ως δότη ακετυλομάδας το ακετυλο-συνένζυμο Α. —O-φωσφοροτρανσφεράσες (APH). Τροποποιούν υδροξυλομάδες. Χρησιμοποιούν ως δότη φωσφορικού το ATP. —O-νουκλεοτιδυλοτρανσφεράσες (ANT). Τροποποιούν υδροξυλομάδες. Χρησιμοποιούν ως δότη νουκλεοτιδυλο-ομάδων το ATP.
. Τροποποιούν υδροξυλομάδες. Χρησιμοποιούν ως δότη φωσφορικού το ATP. —O-νουκλεοτιδυλοτρανσφεράσες (ANT). Τροποποιούν υδροξυλομάδες. Χρησιμοποιούν ως δότη νουκλεοτιδυλο-ομάδων το ATP..")
107
Το όνομα ενός ΑΤΕ περιλαμβάνει α) την ομάδα (ΑAC, APH ή ΑΝΤ) β) έναν αριθμό σε παρένθεση (1, 2, 3, κοκ) που δείχνει τη θέση τροποποίησης γ) έναν Λατινικό αριθμό (Ι, ΙΙ, III, κοκ) για τη διάκριση των ποικιλιών που διαφέρουν στο φάσμα υποστρωμάτων δ) ένα μικρό γράμμα (a, b, c, κοκ) γιά τη διάκριση των ενζύμων που καταλύουν την ίδια αντίδραση και παράγουν τον ίδιο φαινότυπο αλλά κωδικοποιούνται από διαφορετικά γονίδια.
 την ομάδα (ΑAC, APH ή ΑΝΤ) β) έναν αριθμό σε παρένθεση (1, 2, 3, κοκ) που δείχνει τη θέση τροποποίησης γ) έναν Λατινικό αριθμό (Ι, ΙΙ, III, κοκ) για τη διάκριση των ποικιλιών που διαφέρουν στο φάσμα υποστρωμάτων δ) ένα μικρό γράμμα (a, b, c, κοκ) γιά τη διάκριση των ενζύμων που καταλύουν την ίδια αντίδραση και παράγουν τον ίδιο φαινότυπο αλλά κωδικοποιούνται από διαφορετικά γονίδια.")
108
ΠΑΡΑΔΕΙΓΜΑΤΑ APH(3’)-I:μεταφέρει μια φωσφορική ομάδα στο 3’ υδροξύλιο των αμινογλυκοσιδών καναμυκίνη και νεομυκίνη ενώ το APH(3’)-IΙΙ καταλύει την ίδια αντίδραση για καναμυκίνη, νεομυκίνη, αμικασίνη, ισεπαμικίνη. AAC(6’)-Ia, AAC(6’)-Ib, AAC(6’)-Ic είναιακετυλοτρανσφεράσες που μετατρέπουν την 6’ αμινομάδα, παράγουν τον ίδιο φαινότυπο αντοχής (τομπραμυκίνη, νετιλμικίνη, αμικασίνη, καναμυκίνη και διβεκασίνη) αλλά κωδικοποιούνται από διαφορετικά γονίδια.
-Ia, AAC(6’)-Ib, AAC(6’)-Ic είναιακετυλοτρανσφεράσες που μετατρέπουν την 6’ αμινομάδα, παράγουν τον ίδιο φαινότυπο αντοχής (τομπραμυκίνη, νετιλμικίνη, αμικασίνη, καναμυκίνη και διβεκασίνη) αλλά κωδικοποιούνται από διαφορετικά γονίδια..")
109
-Οι μετατροπές αυτές μειώνουν τη συνάφεια μεταξύ των αμινογλυκοσιδών και της 30S υπομονάδας των ριβοσωμάτων. -

110
-Η καταγωγή τους δεν είναι γνωστή (Τα μικρόβια που παράγουν αμινογλυκοσίδες χρησιμοποιούν άλλα συστήματα αυτοπροστασίας) -Οι αμινογλυκοσίδες, πιθανώς, είναι ευκαιριακά και όχι φυσιολογικά υποστρώματα -Τα αντίστοιχα γονίδια αντοχής, κατά κανόνα, φέρονται από ιντεγκρόνια-μεταθετά-πλασμίδια
 -Οι αμινογλυκοσίδες, πιθανώς, είναι ευκαιριακά και όχι φυσιολογικά υποστρώματα -Τα αντίστοιχα γονίδια αντοχής, κατά κανόνα, φέρονται από ιντεγκρόνια-μεταθετά-πλασμίδια")
111
Συχνά ΑΤΕ μεταξύ των εντεροβακτηριακών AAC(3)-II kanamycin, tobramycin, gentamicin, netilmicin (KTGNt) AAC(6’)-I kanamycin, tobramycin, netilmicin, amikacin (KTNtA) ANT(2’’)-I kanamycin, tobramycin, gentamicin (KTG) APH(3’)-I kanamycin, neomycin (KNm) AAC(2’) tobramycin, gentamicin, netilmicin, neomycin (TGNtNm) AAC(3’)-I gentamicin (G) APH(3’’) streptomycin (Str) ANT(3’’) streptomycin (Str) Προσοχή Πολλά κλινικά στελέχη παράγουν περισσότερα του ενός ΑΤΕ. Η φαινοτυπική αναγνώριση είναι δύσκολη Υπερεκτίμηση της αντοχής που οφείλεται σε διαπερατότητα

112
KATANOMH ΑΤΕ ΚΑΤΑ ΓΕΝΟΣ (Νεονάκης και συν. 2005, Παν/μιο Κρήτης)
")
113
Pseudomonas aeruginosa Αμινογλυκοσιδο-τροποποιητικά ένζυμα Ενζυμο Φαινότυπος Συχνότητες Οι κυρίαρχοι ενζυμικοί τύποι ποικίλουν. Οι συχνότητες εξαρτώνται από τους κλώνους που επικρατούν

114
Κινολόνες: Χρήση και αντοχή Neuhauser MM, JAMA 2003;289:885 PA (r=0.976; p<0.001) GNB (r=0.891; p<0.001)
 GNB (r=0.891; p<0.001)")
115
Salmonella typhimurium :Εξέλιξη της αντοχής στις κινολόνες

116
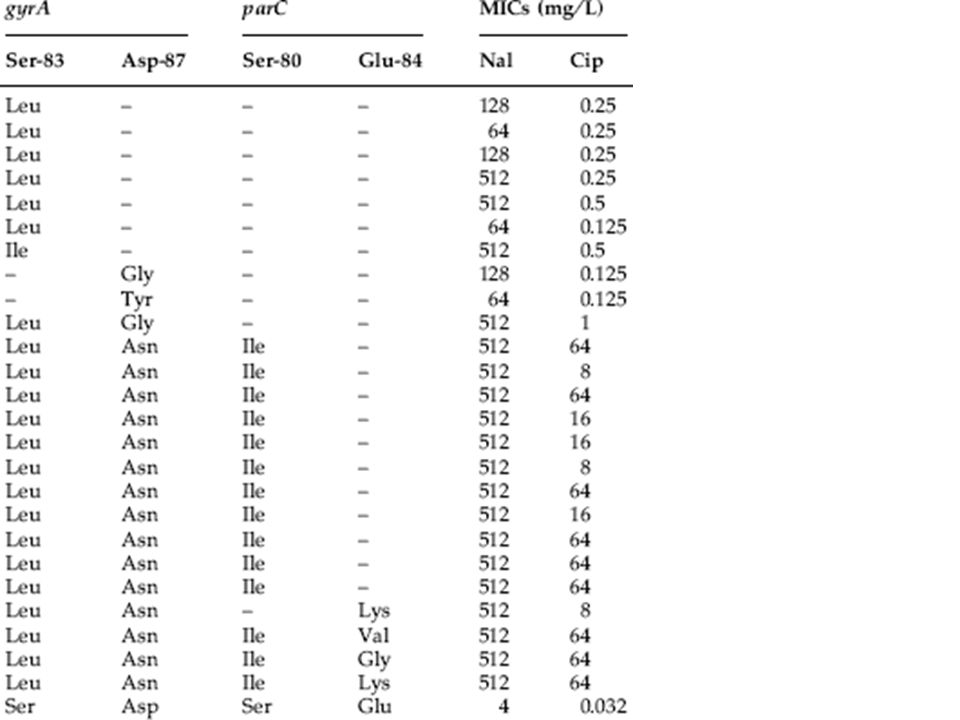
Η αντοχή στις κινολόνες οφείλεται (κυρίως) σε σημειακές μεταλλάξεις G T T G T C A T G T G A C G G G T ευαισθησία G T T G T C A T G T T A C G G G T αντοχή G T (μετάλλαξη) ευαισθησία GAC amino acid Asp αντοχή TAC amino acid Tyr Αποτέλεσμα: Asp87Tyr gyrA gene Codon 87
 σε σημειακές μεταλλάξεις G T T G T C A T G T G A C G G G T ευαισθησία G T T G T C A T G T T A C G G G T αντοχή G T (μετάλλαξη) ευαισθησία GAC amino acid Asp αντοχή TAC amino acid Tyr Αποτέλεσμα: Asp87Tyr gyrA gene Codon 87")
117
ΜΕΤΑΛΛΑΞΕΙΣ ΑΝΤΟΧΗΣ ΣΤΙΣ ΚΙΝΟΛΟΝΕΣ Κωδικόνιο Μετάλλαξη GyrA 51 Ala Val 67 Ala Ser 81 Gly Cys, Asp 82 Asp Gly 83 Ser Leu, Trp, Ala, Val 84 Ala Pro, Val 87 Asp Asn, Gly, Val, Tyr, His 106 Gln Arg, His GyrB 426 Asp Asn 447 Lys Glu

119
-Η αντοχή στις νεότερες κινολόνες προκύπτει από συρροή σημειακών μεταλλάξεων σε gyrA και parC -Συνήθως η πρώτη μετάλλαξη προκαλεί αντοχή στο ναλιδιξικό και μείωση ευαισθησίας στις νεότερες κινολόνες

120
H “ιδιαίτερη” σχέση των κινολονών με τις μεταλλάξεις Η αύξηση των θραυσμάτων DNA από τη δράση των κινολονών μπορεί να προκαλέσει επαγωγή πολυμερασών που είναι επιρρεπείς σε σφάλματα.

121
ΑΝΤΛΙΕΣ ΑΠΟΤΟΞΙΝΩΣΗΣ -Είναι συστήματα μείωσης της ενδοκυττάριας συγκέντρωσης αντιβιοτικών (και άλλων βλαπτικών ουσιών). -Τα συστήματα αυτά είναι συντηρημένα και υπάρχουν σε όλα τα κύτταρα. -Η άντληση γίνεται με κατανάλωση ενέργειας (χρήση ΑΤΡ ή χρήση του ηλεκτροχημικού δυναμικού της μεμβράνης).
..")
122
ΑΝΤΛΙΕΣ GRAM ΑΡΝΗΤΙΚΩΝ 1] Οικογένεια SMR (small multidrug resistant) Παραδειγμα: QacEΔ1 των ιντεγκρονίων -Ο μηχανισμός είναι επίκτητος -Χρησιμοποιεί το δυναμικό της μεμβράνης -Αναγνωρίζει αντισηπτικά (βενζαλκόνιο)
![ΑΝΤΛΙΕΣ GRAM ΑΡΝΗΤΙΚΩΝ 1] Οικογένεια SMR (small multidrug resistant) Παραδειγμα: QacEΔ1 των ιντεγκρονίων -Ο μηχανισμός είναι επίκτητος -Χρησιμοποιεί το δυναμικό της μεμβράνης -Αναγνωρίζει αντισηπτικά (βενζαλκόνιο)](http://images.slideplayer.gr/46/11667517/slides/slide_122.jpg "ΑΝΤΛΙΕΣ GRAM ΑΡΝΗΤΙΚΩΝ 1] Οικογένεια SMR (small multidrug resistant) Παραδειγμα: QacEΔ1 των ιντεγκρονίων -Ο μηχανισμός είναι επίκτητος -Χρησιμοποιεί το δυναμικό της μεμβράνης -Αναγνωρίζει αντισηπτικά (βενζαλκόνιο)")
123
2] Οικογένεια MF (major facilitator) Παραδειγμα: Οι αντλίες TetA, B, C, D κλπ -Χρησιμοποιούν το δυναμικό της μεμβράνης -Αναγνωρίζουν τετρακυκλίνες -Κωδικοποιούνται από πλασμίδια (επίκτητος μηχανισμός)
![2] Οικογένεια MF (major facilitator) Παραδειγμα: Οι αντλίες TetA, B, C, D κλπ -Χρησιμοποιούν το δυναμικό της μεμβράνης -Αναγνωρίζουν τετρακυκλίνες -Κωδικοποιούνται από πλασμίδια (επίκτητος μηχανισμός)](http://images.slideplayer.gr/46/11667517/slides/slide_123.jpg "2] Οικογένεια MF (major facilitator) Παραδειγμα: Οι αντλίες TetA, B, C, D κλπ -Χρησιμοποιούν το δυναμικό της μεμβράνης -Αναγνωρίζουν τετρακυκλίνες -Κωδικοποιούνται από πλασμίδια (επίκτητος μηχανισμός)")
124
3] Οικογένεια MΑΤΕ (multidrug and toxic extrusion) Παραδειγμα: Οι αντλίες Nor -Χρησιμοποιούν το δυναμικό της μεμβράνης -Αναγνωρίζουν κυρίως κινολόνες -Είναι εγγενείς (χρωμοσωματικές) Κλινική σημασία ?
![3] Οικογένεια MΑΤΕ (multidrug and toxic extrusion) Παραδειγμα: Οι αντλίες Nor -Χρησιμοποιούν το δυναμικό της μεμβράνης -Αναγνωρίζουν κυρίως κινολόνες -Είναι εγγενείς (χρωμοσωματικές) Κλινική σημασία](http://images.slideplayer.gr/46/11667517/slides/slide_124.jpg "3] Οικογένεια MΑΤΕ (multidrug and toxic extrusion) Παραδειγμα: Οι αντλίες Nor -Χρησιμοποιούν το δυναμικό της μεμβράνης -Αναγνωρίζουν κυρίως κινολόνες -Είναι εγγενείς (χρωμοσωματικές) Κλινική σημασία")
125
4] Οικογένεια ABC (ATP binding cassette) Παραδειγμα: Η αντλία VcaM (Vibrio) -Χρησιμοποιούν ATP -Αναγνωρίζουν κυρίως κινολόνες -Είναι εγγενείς (Vibrio) ή επίκτητες (πλασμιδιακές) (P. aeruginosa) Κλινική σημασία ?
![4] Οικογένεια ABC (ATP binding cassette) Παραδειγμα: Η αντλία VcaM (Vibrio) -Χρησιμοποιούν ATP -Αναγνωρίζουν κυρίως κινολόνες -Είναι εγγενείς (Vibrio) ή επίκτητες (πλασμιδιακές) (P.](http://images.slideplayer.gr/46/11667517/slides/slide_125.jpg "aeruginosa) Κλινική σημασία .")
126
5] Οικογένεια RND (resistance, nodulation, division) Είναι οι σημαντικότερες εγγενείς αντλίες των Gram-αρνητικών Η ανάπτυξη αντοχής οφείλεται συνήθως σε μεταλλάξεις ρυθμιστικών γονιδίων
![5] Οικογένεια RND (resistance, nodulation, division) Είναι οι σημαντικότερες εγγενείς αντλίες των Gram-αρνητικών Η ανάπτυξη αντοχής οφείλεται συνήθως σε μεταλλάξεις ρυθμιστικών γονιδίων](http://images.slideplayer.gr/46/11667517/slides/slide_126.jpg "5] Οικογένεια RND (resistance, nodulation, division) Είναι οι σημαντικότερες εγγενείς αντλίες των Gram-αρνητικών Η ανάπτυξη αντοχής οφείλεται συνήθως σε μεταλλάξεις ρυθμιστικών γονιδίων")
128
Pseudomonas aeruginosa Αντλιές αποτοξίνωσης (efflux pumps) γιά αμινογλυκοσίδες MexXY -Ανήκουν στη κατηγορία RND. Η σημαντικότερη είναι η MexXY MexXY -H MexXY σχηματίζεται από τρείς πρωτεϊνες α) MexY - πρωτεϊνη άνλησης στη κυτταροπλασματική μεμβράνη β) MexX - πρωτεϊνη / περιπλασμικός σύνδεσμος γ) ΟprM πρωτεϊνη /πόρος εξωτερικής μεμβράνης MexXY -Η αντλία MexXY ειδικεύεται σε αμινογλυκοσίδες, μακρολίδες, τετρακυκλίνες, γλυκυλο-γλυκίνες και χλωραμφαινικόλη -Η αντοχή οφείλεται σε αυξημενη μεταγραφή του οπερονίου mexXY (σημειακές μεταλλάξεις ή επαγωγή).
 MexY - πρωτεϊνη άνλησης στη κυτταροπλασματική μεμβράνη β) MexX - πρωτεϊνη / περιπλασμικός σύνδεσμος γ) ΟprM πρωτεϊνη /πόρος εξωτερικής μεμβράνης MexXY -Η αντλία MexXY ειδικεύεται σε αμινογλυκοσίδες, μακρολίδες, τετρακυκλίνες, γλυκυλο-γλυκίνες και χλωραμφαινικόλη -Η αντοχή οφείλεται σε αυξημενη μεταγραφή του οπερονίου mexXY (σημειακές μεταλλάξεις ή επαγωγή)..")
129
Πορίνες και αντοχή στα αντιβιοτικά -Ολα τα Gram-αρνητικά διαθέτουν πόρους στην εξωτερική μεβράνη (ΕΜ) -Οι πόροι σχηματίζονται από πρωτεϊνες (πορίνες) που “διασχίζουν” την ΕΜ -Οι πόροι επιτρέπουν τη διέλευση υδρόφιλων μορίων -Οι φυσικοχημικές ιδιότητες του πόρου καθορίζουν το φάσμα των μορίων που μπορούν να περάσουν στο περιπλασμικό χώρο (όγκος, σχήμα, φορτία)
 -Οι πόροι σχηματίζονται από πρωτεϊνες (πορίνες) που διασχίζουν την ΕΜ -Οι πόροι επιτρέπουν τη διέλευση υδρόφιλων μορίων -Οι φυσικοχημικές ιδιότητες του πόρου καθορίζουν το φάσμα των μορίων που μπορούν να περάσουν στο περιπλασμικό χώρο (όγκος, σχήμα, φορτία)")
130
Η κύρια πορίνη (OmpF) των εντεροβακτηριακών Μείωση παραγωγής της OmpF προκαλεί μείωση της ευαισθησίας στα αντιβιοτικά
 των εντεροβακτηριακών Μείωση παραγωγής της OmpF προκαλεί μείωση της ευαισθησίας στα αντιβιοτικά")
131
-Περισσότερο επηρεάζονται τα β-λακταμικά -Οι αλλαγές αυτές συνεργούν με τις β-λακταμάσες

132
Συγκρότηματα πολλαπλής αντοχής (Multi-Resistant Islands) -Πρόκειται γιά μεγάλα τμήματα DNA (μέχρι 100 kb) σε πλασμίδια ή χρωμοσώματα -Περιλαμβάνουν πολλαπλά γονίδια αντοχής καθώς και αλληλουχίες μεταθετών και ιντεγκρονίων
 -Πρόκειται γιά μεγάλα τμήματα DNA (μέχρι 100 kb) σε πλασμίδια ή χρωμοσώματα -Περιλαμβάνουν πολλαπλά γονίδια αντοχής καθώς και αλληλουχίες μεταθετών και ιντεγκρονίων")
134
E. coli plasmid pNLT-1 ΔintI1 aadB IS26 aacC1 VEB-1 sulI orfG aadB cmlA5 arr-2 oxa-10 IS26 aadA1 intI1aacA7 IS26 dfrAI aadA1 sulI VIM-1 cmy13 IS26 ampR aphA7 E. coli plasmid p541 IS26 qacEΔ1 qacI qacEΔ1 In-53 In-e541

135
Συσσώρευση γονιδίων αντοχής σε σύστημα IS-ιντεγκρονίου IS26-1 Replicon A IS26/int IS26/Δ5’CS IS26-2 IS26-3 IS26 Chromosomal region Resistance genes Homologous recombination IS26-1 Replicon A IS26/int intI IRt IS26/Δ5’CS IS26-2 IS26-3 IS26/In4 Integration IRt

136
ΔmucBΔorf1ΔIS26dhfrA1aadA1aacA4intIaadA1qacsulI1IS1ΔIS26IS26intIorfX(B)orfX(A)aacC1fucASHV5ΔlacY aacA4intIaadA1qacsulI1IS26orfX’’orfX’aacC1SHV5ΔlacY IS26aacA4intIaadA1qacsulI1ΔIS26IS26orfXaacC1fucASHV5ΔlacYIS26Δorf1orf6orf5 pACM1 Inc L/M pSEM Inc L/M p1658/97 Inc F aacA4intIaadA1qacsulI1aphA1IS26 dhfrIorfOIBC-1 p2815 Inc L/M VIM-1intIaadA1qacsulI1IS26dhfrIaacA7ΔfrdBampRsugEaphA7IS26 cmy13 p541 Inc N orf5IS6100orf6 IRt Δ5’CS Δorf5IS1IS26IS6100orf6 IRt 57bp ΔIRL of IS26 57bp ΔIRL of IS26 orf5IS6100orf6 IRt IS26Δorf5 K. pmeumoniae chromosomal region K. pmeumoniae chromosomal region K. pmeumoniae chromosomal region C.freundii chromosomal region orf5IS6100orf6 IRt IS26 Tn4352 In-t3 In-e541 In111 IS26-chromosomal regions or TransposonIn4-like regions

137
IS26-1 Replicon A IS26/int intI IRt IS26/Δ5’CS IS26-2 IS26-3 Replicon B Transposition through IS26-3 Fusion Replicon A/B IS26-2 IS26-3’ intI IS26-1 IS26/Δ5’CS IS26-3 Homologous recombination between IS26-2 and IS26-3 in recA + cells RepA::IS26 IS26 IS26-3’ RepB::IS26/int intI IS26-2/3 IS26/Δ5’CS IS26-1 IS26-In4 Μεταφορά νησίδας πολλαπλής αντοχής IS26/In4 from repA to repB

138
Δευτερεύοντες* μηχανισμοί αντοχής * Μηχανισμοί που α) προκαλούν χαμηλού επιπέδου (υποκλινική) αντοχή ή β) είναι σπάνιοι ή γ) και τα δύο ή γ) και τα δύο
 προκαλούν χαμηλού επιπέδου (υποκλινική) αντοχή ή β) είναι σπάνιοι ή γ) και τα δύο ή γ) και τα δύο")
139
Προσαρμοστική αντοχή στις αμινογλυκοσίδες Pseudomonas aeruginosa και Pseudomonas aeruginosa -Η αντοχή εμφανίζεται λίγες ώρες μετά από έκθεση σε αμινογλυκοσίδη και «εξαφανίζεται» μετά 10-12 ώρες από την απομάκρυνση του φαρμάκου -Συμβαίνει in vitro και in vivo -Αφορά όλες τις αμινογλυκοσίδες αλλά δεν επηρεάζει άλλα αντιψευδο- μοναδικά αντιβιοτικά ΠΙΘΑΝΗ ΕΞΗΓΗΣΗ Οι αμινογλυκοσίδες προκαλούν αποδιοργάνωση του τοιχώματος και μεταφραστικά σφάλματα Πιθανόν τα παραπάνω να μειώνουν παροδικά την αερόβια αναπνοή Μείωση του δυναμικού της μεμβράνης Χαμηλός ρυθμός εισόδου του αντιβιοτικού Παροδική αντοχή

140
Αλλαγές στην εξωτερική μεμβράνη κ’ αντοχή στις αμινογλυκοσίδες (Pseudomonas aeruginosa) ΓΕΝΙΚΟΣ ΜΗΧΑΝΙΣΜΟΣ : Υποβάθμιση του ρόλου των ιόντων μαγνησίου στη κυτταρική επιφάνεια.
 ΓΕΝΙΚΟΣ ΜΗΧΑΝΙΣΜΟΣ : Υποβάθμιση του ρόλου των ιόντων μαγνησίου στη κυτταρική επιφάνεια.")
141
-Μεταλλάξεις στο οπερόνιο pho P-Q Αύξηση αμινοαραβινόζης στο λιπίδιο Α Μερική αντικατάσταση Mg++ Μείωση των θέσεων δέσμευσης αμινογλυκοσιδών -Μεταλλάξεις που αυξάνουν τη ποσότητα της OmpH H OmpH είναι κατιονική πρωτεϊνη της εξωτερικής μεμβράνης. Τα επιφανειακά θετικά φορτία της πιθανόν αντικαθιστούν τις γέφυρες Mg++ Μείωση των θέσεων δέσμευσης αμινογλυκοσιδών

142
Πλασμιδιακή αντοχή στις κινολόνες -Οφείλεται στις πρωτεϊνες Qnr οι οποίες “προστατεύουν” τη γυράση -Προκαλούν μικρή μείωση τις ευαισθησίας

143
Τα γονίδια qnr βρίσκονται σε σύνθετα ιντεγκρόνια... …που φέρονται από πλασμίδια πολυαντοχής των εντεροβακτηριακών Τα γονίδια qnr πιθανόν προέρχονται από θαλάσσιους μικροοργανισμούς (Shewanella algae)
.")
144
Μεθυλάσες της μικρής υπομονάδας του ριβοσώματος

145
-Παρεμόδιση σύνδεσης όλων των αμινογλυκοσιδών -Υψηλού επιπέδου αντοχή σε όλες τις αμινογλυκοσίδες -Τα γονίδια των μεθυλασών φέρονται από κινητά στοιχεία -Πιθανόν προέρχονται από Ακτινομύκητες που παράγουν αμινογλυκοσίδες.

146
ΑΝΤΙΜΕΤΩΠΙΣΗ ΤΗΣ ΑΝΤΟΧΗΣ -Νέα αντιβιοτικά ? -Μείωση της χρήσης αντιβιοτικών ? -Κυκλική χρήση αντιβιοτικών ? -Μείωση της διασποράς ?