Κατέβασμα παρουσίασης
1
Δρ. Ευριπίδου Πολύκαρπος C.D.A.College Limassol 2015/2016

2
Οι υποδοχείς της κυτταρικής μεμβράνης διακρίνονται σε δύο επιμερούς ομάδες. Οι υποδοχείς της πρώτης ομάδας συνδέονται με μία κινάση τυροσίνης ενώ Οι υποδοχείς της δεύτερης ομάδας συνδέονται με τις πρωτεΐνες G. Ωστόσο η δομή και των δύο ομάδων είναι παρόμοια. Ο υποδοχέας αποτελείται από τρία τμήματα: 1. Μια εξωκυττάρια περιοχή (ectodomain), η οποία παρουσιάζει υψηλή συγγένεια για την αντίστοιχη ορμόνη.
, η οποία παρουσιάζει υψηλή συγγένεια για την αντίστοιχη ορμόνη..")
3
2. Μια διαμεμβρανική περιοχή. Η δομή της ποικιλλεί: μπορεί να συνιστάται απλά σε μία γραμμική υδρόφοβη αλυσίδα αμινοξέων ή να είναι μία περισσότερο ελικοειδής δομή που διασχίζει τη μεμβράνη επτά φορές. 2. Μία ενδοκυττάρια περιοχή, η οποία αναλαμβάνει την ενδοκυττάρια διαβίβαση του σήματος. Η οδός της ενδοκυττάριας διαβίβασης του σήματος μπορεί να είναι αρκετά πολύπλοκη, με αρκετά σημεία διακλάδωσης. Η διαδοχική φωσφορυλίωση πρωτεϊνών παίζει κεντρικό ρόλο στη διαβίβαση του σήματος.

4
Τα αμινοξέα σερίνη, θρεονίνη και τυροσίνη διαθέτουν μία πολική ομάδα υδροξυλίου. Η αντικατάσταση του υδροξυλίου αυτού από μία φωσφορική ομάδα που προέρχεται από ένα μόριο ΑΤΡ ονομάζεται φωσφορυλίωση. Η φωσφορυλίωση έχει ως αποτέλεσμα τη δημιουργία ενός ομοιοπολικού δεσμού μεταξύ του αμινοξέος και του φωσφορικού ανιόντος.

5
Κατά την αντίδραση αυτή παρατηρείται μεταφορά ενέργειας, η οποία οδηγεί στην ενεργοποίηση και την αλλαγή της διαμόρφωσης της φωσφορυλιωμένης πρωτεΐνης στο χώρο. Σε πολλές περιπτώσεις η ίδια η φωσφορυλιωμένη πρωτεΐνη μπορεί με τη σειρά της να δράσει πλέον σαν πρωτεϊνική κινάση με αποτέλεσμα τη φωσφορυλίωση της επόμενης πρωτεΐνης και την ενδοκυττάρια μετάδοση του μηνύματος μέσω μίας αλληλουχίας φωσφορυλιώσεων (phosphorylation cascade).
..")
6
Η φωσφορυλίωση των πρωτεϊνών παίζει το ρόλο ενός «μοριακού διακόπτη». Οι πολικές υδροξυλικές ομάδες της σερίνης και της θρεονίνης φωσφορυλιώνονται σε πολύ μεγαλύτερο βαθμό από τις αντίστοιχες της τυροσίνης. Αυτό σημαίνει ότι η πλειονότητα των φωσφορυλιωμένων πρωτεϊνών των κυττάρων των θηλαστικών (οι οποίες αποτελούν το 10% του συνόλου των πρωτεϊνων) αφορά την φωσφοσερίνη και φωσφοθρεονίνη.
 αφορά την φωσφοσερίνη και φωσφοθρεονίνη..")
7
Μέχρι το 1980, θεωρούσαμε ότι η φωσφοσερίνη και η φωσφοθρεονίνη ήταν τα μόνα φυσικά φωσφοαμινοξέα. Η φωσφοτυροσίνη ευθύνεται για το 0,1% μόνο των φωσφορυλιωμένων πρωτεϊνών. Η φωσφορυλιωση της τυροσίνης προσδίδει ιδιαίτερα χαρακτηριστικά στη συγκεκριμένη πρωτεΐνη, λόγω του μεγάλου μήκους της πλευρικής αλύσου και της μεγάλης πυκνότητας του ηλεκτρονικού νέφους στον αρωματικό δακτύλιο της φωσφοτυροσίνης.

8
Η φωσφορυλίωση της τυροσίνης παίζει ιδιαίτερα σημαντικό ρόλο στη διαβίβαση του μηνύματος στις περιπτώσεις της ινσουλίνης και της αυξητικής ορμόνης. Η φωσφορυλίωση της τυροσίνης εκτός από την ενεργοποίησης μίας πρωτεΐνης, έχει σαν αποτέλεσμα και τη δημιουργία «θέσεων σύνδεσης» στο μόριο, οι οποίες βοηθούν στην αλληλεπίδραση μεταξύ των πρωτεϊνών.

9
Εχουν βρεθεί περιοχές πρωτεϊνών (domains), γνωστές ως SH2 και SH3, οι οποίες είναι συμπληρωματικές προς τη φωσφοτυροσίνη. Οι περιοχές αυτές έχουν μήκος 100 περίπου αμινοξέων και περιλαμβάνουν μία στρατηγικά τοποθετημένη αργινίνη (Arg-175), που μπορεί να συνδεθεί με τη φωσφοτυροσίνη αλλά όχι με τη φωσφοσερίνη ή τη φωσφοθρεονίνη. Η σχετικά επιμηκής αλυσίδα της φωσφοτυροσίνης επιτρέπει τη «σύνδεση» (docking) μίας πρωτεΐνης με τη σχισμή που σχηματίζει η SH2/SH3 καταλυτική περιοχή των φωσφατασών τυροσίνης.
, που μπορεί να συνδεθεί με τη φωσφοτυροσίνη αλλά όχι με τη φωσφοσερίνη ή τη φωσφοθρεονίνη. Η σχετικά επιμηκής αλυσίδα της φωσφοτυροσίνης επιτρέπει τη «σύνδεση» (docking) μίας πρωτεΐνης με τη σχισμή που σχηματίζει η SH2/SH3 καταλυτική περιοχή των φωσφατασών τυροσίνης..")
10
Οι SH2/SH3 περιοχές διαθέτουν μία χαρακτηριστική μοριακή τοπολογία και συναντώνται σε μία ποικιλία κυτταροπλασματικών σηματοδοτικών πρωτεϊνών. Χρησιμεύουν για τη σύνδεση των ανενεργών, διαλυτών κινασών τυροσίνης με ορμονικούς υποδοχείς της κυτταρικής μεμβράνης οι οποίοι έχουν ενεργοποιηθεί από την αντίστοιχη ορμόνη. Σε άλλες περιπτώσεις, ο ρόλος των SH2/SH3 περιοχών είναι περισσότερο παθητικός, π.χ. Διευκολύνοντας τη σύνδεση των πρωτεϊνών που συμμετέχουν σε μία αλληλουχία φωσφορυλιώσεων (phosphorylation cascade).
..")
11
Οι υποδοχείς της κυτταρικής μεμβράνης διακρίνονται σε δύο επιμερούς ομάδες. Οι υποδοχείς της πρώτης ομάδας συνδέονται με μία κινάση τυροσίνης και βασίζονται στη δράση της τελευταίας για την περαιτέρω μεταβίβαση του σήματος. Οι υποδοχείς της δεύτερης ομάδας συνδέονται με τις πρωτεΐνες G. Οι πρωτεΐνες G επιστρατεύουν ενδοκυττάριους δεύτερους αγγελιαφόρους, οι οποίοι συνήθως χρησιμοποιούν κινάσες σερίνης/θρεονίνης.

12
Οι υποδοχείς αυτοί αποτελούνται από μία σχετικά απλή διαμεμβρανική περιοχή, ενω η ενδομεμβρανική περιοχή του μορίου μπορεί να διαθέτει ιδιότητες κινάσης τυροσίνης ή να επιστρατεύσει ενδοκυττάριες κινάσες τυροσίνης μετά τη διέγερση του υποδοχέα από την αντίστοιχη ορμόνη.

13
Οι κυριότεροι υποδοχείς της υποομάδας αυτής, από ενδοκρινολογική άποψη, είναι οι υποδοχείς της ινσουλίνης. Οι υποδοχείς αυτοί συνδέονται γενικότερα με μόρια όπως ο επιδερμιδικός αυξητικός παράγοντας (EGF) και ο αυξητικός παράγοντας των ινοβλαστών (FGF), οι οποίοι προάγουν την αύξηση και τον πολλαπλασιασμό των κυττάρων, γι’αυτό και συχνά αναφέρονται και ως «υποδοχείς των αυξητικών παραγόντων».
 και ο αυξητικός παράγοντας των ινοβλαστών (FGF), οι οποίοι προάγουν την αύξηση και τον πολλαπλασιασμό των κυττάρων, γι’αυτό και συχνά αναφέρονται και ως «υποδοχείς των αυξητικών παραγόντων»..")
14
Στη υποομάδα αυτή ανήκουν και οι υποδοχείς του ινσουλινόμορφου αυξητικού παράγοντα 1 (IGF 1). Στην περίπτωση των EGF και FGF, η σύνδεση των παραγόντων προκαλεί το διμερισμό δύο παρακείμενων μονομερών υποδοχέων. Το φαινόμενο του διμερισμού ενός υποδοχέα μετά τη σύνδεσή του με τον αντιστοιχο διεγέρτη μας είναι αρκετά γνωστό. Ο διμερισμός οδηγεί στην ενεργοποίηση μίας κινάσης τυροσίνης, η οποία εμπεριέχεται στη ενδοκυττάρια περιοχή του υποδοχέα.

15
Σε αντίθεση με τα παραπάνω, οι υποδοχείς της ινσουλίνης και του IGF 1 σχηματίζουν ήδη διμερή, ακόμα και όταν δεν είναι συνδεδεμένοι με τα αντίστοιχα μόρια. Η προσκόλληση ενός μορίου ινσουλίνης ή IGF 1 στην περιοχή σύνδεσης που σχηματίζουν οι δύο α-υπομονάδες, οδηγεί στην ενεργοποίηση της ενδογενούς κινάσης τυροσίνης της ενδοκυττάριας περιοχής του υποδοχέα.

16
Η πρώτη συνέπεια της σύνδεσης της ινσουλίνης με τον υποδοχέα της είναι η αυτοφωσφορυλίωση των β- υπομονάδων του ίδιου του υποδοχέα. Αυτό οδηγεί σε μία πολύπλοκη σειρά πρωτεϊνικών αλληλεπιδράσεων, η οποία περιλαμβάνει περισσότερα από 50 ένζυμα. Η ενδογενής κινάση τυροσίνης του υποδοχέα στη συνέχεια φωσφορυλιώνει ένα σηματνικό υπόστρωμα, γνωστό ως Insulin Receptor Substrate 1 ή 2 (IRS 1 ή 2, υπόστρωμα του υποδοχέα της ινσουλίνης).
..")
17
Η φωσφορυλίωση αυτή φαίνεται ότι είναι ουσιαστικής σημασίας για οποιαδήποτε άλλη βιολογικά δράση της ινσουλίνης. Το IRS-1 είναι μία πρωτεΐνη 131 kDA, η οποία διαθέτει 21 αμινοξέα τυροσίνης, πιθανούς στόχους φωσφορυλίωσης. Από αυτά, τα 8 τουλάχιστον φωσφορυλιώνονται από τον ενεργοποιημένο υποδοχέα της ινσουλίνης. Η πολλαπλή φωσφορυλίωση του IRS-1 οδηγεί στη σύνθεση των φωσφοτυροσίνων του μορίου αυτού με τις SH2/SH3 περιοχές διαφόρων πρωτεϊνών.

18
Στο σημείο αυτό αρχίζει μέσα στο κύτταρο η παράλληλη ενεργοποίηση διαφορετικών σηματοδοτικών οδών. Για παραδειγμα, η σύνδεση του IRS-1 με την p85 υπομονάδα της κινάσης της φωσφατιδυλοϊνοσιτόλης-3 (PI-3) προκαλεί τη μετανάστευση πρωτεϊνών-μεταφορέων (transporters) της γλυκόζης από το εσωτερικό του κυττάρου προς την κυτταρική μεμβράνη. Έχουν περιγραφεί διάφορες πρωτεΐνες- μεταφορείς γλυκόζης, οι GLUT 1-4 και η SGLT.
 προκαλεί τη μετανάστευση πρωτεϊνών-μεταφορέων (transporters) της γλυκόζης από το εσωτερικό του κυττάρου προς την κυτταρική μεμβράνη. Έχουν περιγραφεί διάφορες πρωτεΐνες- μεταφορείς γλυκόζης, οι GLUT 1-4 και η SGLT..")
19
Στους κύριους ιστούς-στόχους της ινσουλίνης, δηλασή το λιπώδη ιστό και τον καρδιακό μυ, κυριαρχεί ο GLUT 4. Πρόκειται για μία πρωτεΐνη που αποτελείται από 520 αμινοξέα, το μόριο της οποίας διατρέχει 12 φορές το πάχος της κυτταρικής μεμβράνης και ευθύνεται για τη μεταφορά της γλυκόζης στο εσωτερικό του κυττάρου. Η σύνδεση της ινσουλίνης με τους υποδοχείς ινσουλίνης ενός κυττάρου οδηγεί στη σύντηξη κυτταροπλασματικών κυστιδίων που φέρουν GLUT4 με την κυτταρική μεμβράνη.

20
Όταν η ινσουλίνη απομακρυνθεί από τους υποδοχείς, τα μόρια GLUT 4 αποσπώνται από την κυτταρική μεμβράνη με τη μορφή κυτταροπλασματικών κυστιδίων. Παράλληλα με τα ανωτέρω παρατηρείται αυξημένη δραστηριότητα της συνθετάσης γλυκογόνου, με αποτέλεσμα την αυξημένη παραγωγή γλυκογόνου, καθώς και αυξημένη έκφραση του γονιδίου που κωδικοποιεί της εξοκινάση-2.

21
Η βιοψία μυός ασθενών με μη ινσουλινοεξαρτώμενο διαβήτη έχει καταδείξει μειωμένη δραστηριότητα συνθετάσης γλυκογόνου και εξοκινάσης-2, ενώ έχουν επίσης αναφερθεί μείωση των επιπέδων του IRS-1 και της p85 υπομονάδας της κινάσης της PI-3. Η μιτογόνος δράση της ινσουλίνης οφείλεται σε μία δεύτερη οδό που αρχίζει επίσης με τη φωσφορυλίωση του IRS-1. Σε αυτήν την περίπτωση οι φωσφοτυροσίνες του IRS-2 δεν συνδέονται με τις SH2 περιοχές της κινάσης της PI- 3 αλλά με τις SH2/SH3 περιοχές της συνδεδεμένης με τον υποδοχέα του αυξητικού παράγοντα πρωτεΐνης-2 (growth factor receptor-bound protein 2 – πρωτεΐνη Grb2).
..")
22
Η πρωτεΐνη Grb2 μεσολαβεί μεταξύ υποδοχέων με φωσφορυλιωμένα τυροσίνης ή κυτταροπλασματικών κινάσων τυροσίνης και μορίων που ρυθμίζουν τη δραστηριότητα μικρών πρωτεϊνών G. Στη συγκεκριμένη περίπτωση η Grb2 μεσολαβεί μεταξύ του IRS-2 και της Sos πρωτεΐνης (son of sevenless protein), η οποία προκαλεί ανταλλαγή ενός GDP μορίου με ένα GTP. Το σύμπλεγμα Sos-Grb 2 στη συνέχεια πλησιάζει στην πρωτεΐνη Ras, η οποία βρίσκεται στην κυτταρική μεμβράνη.
, η οποία προκαλεί ανταλλαγή ενός GDP μορίου με ένα GTP. Το σύμπλεγμα Sos-Grb 2 στη συνέχεια πλησιάζει στην πρωτεΐνη Ras, η οποία βρίσκεται στην κυτταρική μεμβράνη..")
23
Αυτό οδηγεί στην ενεργοποίηση της Ras μέσω της ανταλλαγής GTP/GDP. Η ενεργοποίηση της Ras προκαλεί μία αλληλουχία αντιδράσεων, στις οποίες συμμετέχουν κινάσες σερίνης/θρεονίνης. Μία από αυτές, η Raf, κινητοποιεί μία άλλη κίναση σερίνης/θρεονίνης, τη ΜΕΚ, η οποία με τη σειρά της κινητοποιεί μία πολύπλευρη κινάση σερίνης/θρεονίνης που παίζει κεντρικό ρόλο στη μιτογόνο δράση της ινσουλίνης.

24
Η κινάση αυτή αρχικά ονομάστηκε πρωτεϊνική κινάση μικροσωληνίσκων (microtubule-as-sociated protein kinase). Είναι γνωστή ως πρωτεϊνική κινάση που ενεργοποιείται από μιτογόνες ουσίες (mitogen- activated protein kinase - MAPK). Η MAPK δρα σε διάφορες πρωτεΐνες του κυτταροπλάσματος και του πυρήνα του κυττάρου. Οι τελευταίες διεγείρουν τη γονιδιακή έκφραση, την πρωτεϊνοσύνθεση και την αύξηση του κυττάρου.
. Η MAPK δρα σε διάφορες πρωτεΐνες του κυτταροπλάσματος και του πυρήνα του κυττάρου. Οι τελευταίες διεγείρουν τη γονιδιακή έκφραση, την πρωτεϊνοσύνθεση και την αύξηση του κυττάρου..")
25
Έχουν αναφερθεί περισσότερες από 50 διαφορετικές μεταλλάξεις του υποδοχέα της ινσουλίνης. Μεταξύ άλλων οδηγουν και σε τρία σύνδρομα σημαντικής αντίστασης των ιστών στην ινσουλίνη, τα οποία με σειρά αυξανόμενης βαρύτητας είναι, 1.Η αντίσταση των ιστών στην ινσουλίνη τύπου Α, 2.Το σύνδρομο Rabson-Mendenhall, 3 και το σύνδρομο Leprachaunism/Donahues.

26
Στους ασθενείς αυτούς παρατηρούνται διαταραχές του μεταβολισμού της γλυκόζης και αυξημένα επίπεδα ινσουλίνης στην κυκλοφορία. Οι ασθενείς με αντίσταση των ιστών στην ινσουλίνη τύπου Α συνήθως διαγιγνώσκονται κατά την εφηβεία. Αντιθέτως οι ασθενείς με σύνδρομο Donahues εμφανίζουν βαριές διαταραχές τη ενδομήτριας ανάπτυξης και σπανίως επιζούν πέραν της βρεφικής ηλικίας.

27
Η σοβαρή αυτή μορφή αντίστασης των ιστών στην ινσουλίνη φαίνεται να οφείλεται στην απουσία λειτουργικών υποδοχέων ινσουλίνης. Αντιθέτως στο σύνδρομο Rabson-Mendenhall οι υποδοχείς ινσουλίνης είναι ελαττωματικοί αλλά όχι τελείως αδρανείς. Σε ορισμένους ασθενείς με αντίσταση των ιστών στην ινσουλίνη τύπου Α έχουν αναφερθεί μεταλλάξεις του τμήματος του υποδοχέα που έχει ιδιότητες κινάσης τυροσίνης.

28
Στην πλειονότητα των ασθενών με το σύνδρομο αυτό, οι υποδοχείς ινσουλίνης είναι καθ’όλα φυσιολογικοί. Στις περιπτώσεις αυτές είναι πιθανό να υπάρχουν μεταλλάξεις σε κάποιο άλλο μόριο της σηματοδοτικής οδού.

29
Οι σημαντικότερες ορμόνες της δεύτερης αυτής υποομάδας υποδοχέων είναι η αυξητική ορμόνη (GH), η προλακτίνη (PRL) και η λεπτίνη. Οι ορμόνες αυτές, όπως και διάφορες κυττοκίνες (π.χ οι ιντερλευκίνες και η ερυθροποιητίνη), έχουν ένα κοινό δομικό χαρακτηριστικό: αποτελόυνται από τέσσερις αντιπαράλληλες μεταξύ τους επιμηκείς άλφα έλικες. Για το λόγο αυτό, οι υποδοχείς της υποομάδας αυτής είναι γνωστοί και ως υποδοχείς των κυττοκινών και των αιμοποιητικών παραγόντων.
, έχουν ένα κοινό δομικό χαρακτηριστικό: αποτελόυνται από τέσσερις αντιπαράλληλες μεταξύ τους επιμηκείς άλφα έλικες. Για το λόγο αυτό, οι υποδοχείς της υποομάδας αυτής είναι γνωστοί και ως υποδοχείς των κυττοκινών και των αιμοποιητικών παραγόντων..")
30
Έχουν περιγραφεί τουλάχιστον 20 μέλη, η βασική δομή των οποίων είναι παρόμοια. Η εξωκυττάρια περιοχή συνδέεται με την αντίστοιχη ορμόνη, έχει μήκος 200 περίπου αμινοξέων και είναι η περιοχή που παρουσιάζει τη μεγαλύτερη ομολογία μεταξύ των διαφορών υποδοχέων. Η περιοχή αυτή του υποδοχέα είναι σταθερή (η αλληλουχία των αμινοξεών δεν ποικίλλει σε διαφορετικούς υποδοχείς της ίδιας κατηγορίας).
..")
31
Στερεοσκοπικά παρατηρούνται δύο σχηματισμοί που διαμορφώνουν ένα θύλακο στον οποίο συνδέεται η αντίστοιχη ορμόνη. Το μήκος της ενδοκυττάριας περιοχής του υποδοχέα ποικίλλει και υπάρχουν ελάχιστες ομοιότητες μεταξύ διαφορετικών υποδοχέων. Ομως μία αλληλουχία αμινοξέων πλησίον της κυτταρικής μεμβράνης, γνωστή ως Box 1, είναι εξαιρετικά σταθερή. Η αλληλουχία αυτή φαίνεται να είναι ιδιαίτερα σημαντική για τη μιτογόνο δράση των ορμονών στα κύτταρα-στόχους.

32
Η κρυσταλλική δομή του συμπλέγματος της ανθρώπειου αυξητικής ορμόνης (GH) με τον υποδοχέα της διευκρινίσθηκε το 1992. Πρόκειται για ένα τριμερές σύμπλεγμα που σχηματίζεται από ένα μόριο ορμόνης και δύο μόρια υποδοχέα. Παράλληλες έρευνες που βασίζονταν στη μελέτη μεταλλαγμένων μορίων GH έδειξαν ότι η ορμόνη διαθέτει δύο περιοχές που μπορούν να συνδεθούν με τον υποδοχέα.

33
Επιπλέον το σύμπλεγμα σχηματίζεται σε δύο στάδια. Αρχικά η GH συνδέεται με ένα μόριο υποδοχέα και στη συνέχεια το διμερές σύμπλεγμα ενώνεται με ένα δεύτερο μόριο υποδοχέα. Ο διμερισμός της ενδοκυττάριας περιοχής των δύο μορίων υποδοχέα μετά το σχηματισμό του τριμερούς συμπλέγματος έχει ιδιαίτερη σημασία για τη μετάδοση του σήματος. Ανάλογος διμερισμός μεταξύ όμοιων μορίων έχει περιγραφεί και στις περιπτώσεις της προλακτίνης και της ερυθροποιητίνης.

34
Περαιτέρω μελέτες με κυττοκίνες απέδειξαν ότι το μοντέλο αυτό (του διμερισμού ομοίων μορίων υποδοχέα) ισχύει για μία μειονότητα των κυττοκινών, ενώ στις υπόλοιπες κυττοκίνες σχηματίζονται διμερή ή ολιγομερή μεταξύ διαφόρων ενδοκυττάριων πρωτεϊνών. 1. Επιστράτευση κινασών τυροσίνης. Αν και οι ίδιοι οι υποδοχείς δεν διαθέτουν περιοχές με ενζυμική δράση κινάσης τυροσίνης, μετά τη σύνδεση τους με κυττοκίνες, όπως η ιντερλευκίνη 2 και 3, ή με την GH ή την PRL, παρατηρείται φωσφορυλίωση τυροσίνων διαφόρων πρωτεϊνών, συμπεριλαμβανομένης της ενδοκυττάριας περιοχής του ίδιου του υποδοχέα.

35
Η φωσφορυλίωση των τυροσινών ανιχνεύεται μέσα σε λίγα λεπτά από την έκθεση των κυττάρων-στόχων στα αντίστοιχα μόρια- διεγέρτες. Με τεχνικές cross-linking και ανοσοκαθίζησης διαπιστώθηκε ότι η κινάση τυροσίνης δεν κινητοποιείται απλά μετά από τη σύνδεση της αυξητικής ορμόνης με τον υποδοχέα της αλλά σχετίζεται με τον ίδιο τον υποδοχέα. Η εν λόγω κινάση, η οποία κινητοποιείται από διμερή μόρια του υποδοχέα ονομάστηκε κινάση του Ιανού (JAK-2).
..")
36
2. Κινάσες του Ιανού (Janus Associated Kinases - JAK). Όταν πρωτοανακαλύφθηκαν, οι κινάσες της οικογένειας JAK ήταν «ορφανές» δηλαδή δεν γνωρίζαμε την οδό κινητοποίησης τους, ούτε τον τρόπο δράσης τους. Γι’αυτό και ονομάστηκαν «Just Another Kinase» (JAK) («απλά άλλη μία κινάση»). Ωστόσο σε τέσσερεις από αυτές (JAK1-3 και Tyk- 2) βρέθηκε μία χαρακτηριστική δομή, συγκεκριμένα δύο συνεχόμενες περιοχές με ιδιότητες κινάσης στο καρβοξυτελικό άκρο του μορίου.
 («απλά άλλη μία κινάση»). Ωστόσο σε τέσσερεις από αυτές (JAK1-3 και Tyk- 2) βρέθηκε μία χαρακτηριστική δομή, συγκεκριμένα δύο συνεχόμενες περιοχές με ιδιότητες κινάσης στο καρβοξυτελικό άκρο του μορίου..")
37
Για αυτο μετονομάστηκαν σε «Janus Associated Kinases» (JAK) («κινάσες του Ιανού»), από το όνομα του Ιανού, της Ρωμαϊκής θεότητας που είχε δύο πρόσωπα. Οι κινάσες αυτές είναι κυτταροπλασματικές πρωτεΐνες μήκους 120-135 kDa και η ομολογία των μορίων τους ανέρχεται στο 40%. Η ενζυμική δράση του μορίου οφείλεται στην JH1 περιοχή, η οποία βρίσκεται στο καρβοξυτελικό άκρο, ενώ παρατηρείται και μία JH2 περιοχή, η οποία αν και μοιάζει με κινάση δεν διαθέτει κάποιο τμήμα που να μπορεί να συνδέεται με μόρια ATP.

38
Η JH2 πιθανώς ρυθμίζει τη λειτουργία της JH1. Τα μόρια αυτά δεν διαθέτουν περιοχές SH2/SH3 και ο μηχανισμός μέσω του οποίου κινητοποιούνται δεν έχει ακόμα αποσαφηνιστεί. Οι υποδοχείς της GH, της PRL και της ερυθροποιητίνης χρησιμοποιούν την JAK-2, αν και υπάρχουν ενδείξεις ότι ο υποδοχέας της PRL είναι μόνιμα συνδεδεμένος με τη JAK-2. Στους υποδοχείς της GH και της PRL έχει εντοπιστεί ένα πλούσιο σε προλίνη τμήμα της ενδοκυττάριας περιοχής, γνωστό ως Box 1, το οποίο συνδέεται με την JAK-2, με αποτέλεσμα την κινητοποίηση της τελευταίας.

39
Είναι πιθανό ο διμερισμός του υποδοχέα να βοηθά στην προσέγγιση δύο μορίων JAK-2, γεγονός που επιτρέπει την αλληλοφωσφορυλίωση των καταλυτικών κέντρων των δύο μορίων.

40
3. Πρωτεΐνες STAT (Signal Transduction and Activation of Transcription). Ένα βασικό υπόστρωμα των JAK κινασών, πλην των ίδιων και των υποδοχέων των κυττοκινών, είναι οι λεγόμενες πρωτεΐνες STAT. Πρόκειται για πρωτεΐνες που περιέχουν 700-800 αμινοξέα και παρουσιάζουν ομολογία 30-40% μεταξύ των μορίων τους. Οι πρωτεΐνες STAT ουσιαστικά κινητοποιούνται μετά από τη φωσφορυλίωση ενός υπολείμματος τυροσίνης που βρίσκεται στο καρβοξυτελικό άκρο και συγκεκριμένα στη θέση 694 κάθε πρωτεΐνης της οικογένειας αυτής.

41
Η λειτουργία της κινητοποιημένης πρωτεΐνης είναι διπλή: (i) φροντίζει για την ενδοκυττάρια μετάδοση του σήματος (signal tranduction) και (ii) για τη μεταγραφή κατάλληλων γονιδίων στον πυρήνα (activation of transcription). Οι πρωτεΐνες STAT έχουν ονομαστεί με τη σειρά που ταυτοποιήθηκαν. Η GH, η PRL και η ερυθροποιητίνη επάγουν την φωσφορυλίωση τυροσινών των πρωτεϊνών STAT1, 3, 5 α και 5β, αλλά η STAT 5 φαίνεται να είναι η βασικότερη εξ αυτών.

42
Η φωσφορυλίωση των STAT πρωτεϊνών επίσης ανιχνεύεται σχετικά νωρίς μετά την έκθεση των κυττάρων-στόχων στις αντίστοιχες ορμόνες. Οι SH2 περιοχές των STAT πρωτεϊνών τους επιτρέπει να συνδέονται με στρατηγικά τοποθετημένες φωσφοτυροσίνες των υποδοχέων των κυττοκινών. Μετά τη σύνδεση τους, φωσφορυλιώνοται από τις JAK κινάσες οι οποίες έχουν επίσης επιστρατευτεί από το σύμπλεγμα υποδοχέα/JAK και σχηματίζουν μεταξύ τους ομο- ή ετεροδιμερή στο κυτταρόπλασμα.

43
Ο σχηματισμός διμερών φαίνεται να έχει σημασία για τη μετανάστευση των μορίων στον πυρήνα. Στον πυρήνα διεγείρουν τη μεταγραφή άμεσων (immediate early) γονιδίων που ρυθμίζουν τον πολλαπλασιασμό του κυττάρου, ή ειδικότερων γονιδίων που είναι υπεύθυνα για τη διαφοροποίηση του κυττάρου-στόχου. 4. Άλλες σηματοδοτικές οδοί. Η GH και η PRL δεν κινητοποιούν τα κύτταρα-στόχους μόνο μέσω της JAK/STAT οδού.
 γονιδίων που ρυθμίζουν τον πολλαπλασιασμό του κυττάρου, ή ειδικότερων γονιδίων που είναι υπεύθυνα για τη διαφοροποίηση του κυττάρου-στόχου. 4. Άλλες σηματοδοτικές οδοί. Η GH και η PRL δεν κινητοποιούν τα κύτταρα-στόχους μόνο μέσω της JAK/STAT οδού..")
44
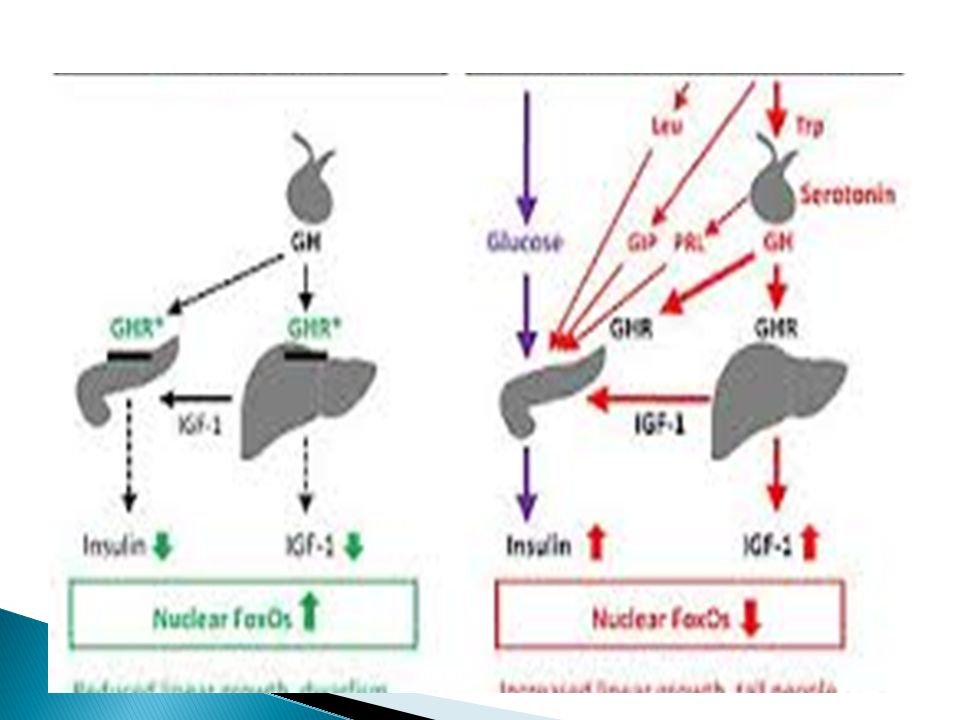
Η σύνδεση της ορμόνης με τον υποδοχέα κινητοποιεί και τις οδούς MAPK και PI-3. Η επικάλυψη των σηματοδοτικών οδών της GH με αυτές της ινσουλίνης ίσως ερμηνεύει την οξεία ινσουλινόμορφη δράση της GH.

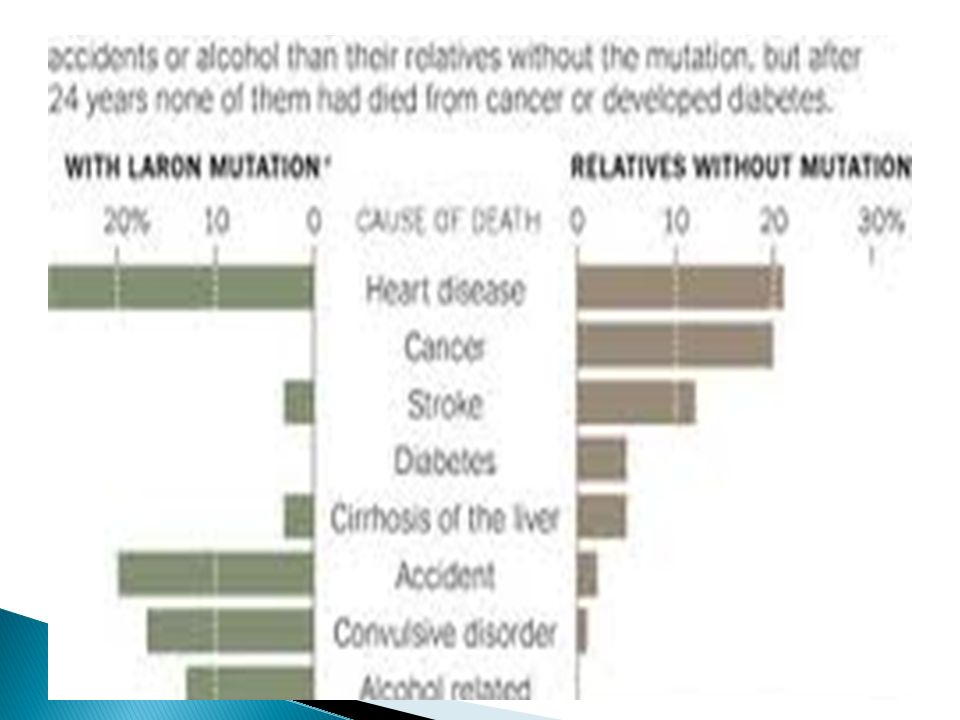
45
Η αντίσταση των περιφερικών ιστών στην GH πρωτοπεριγράφηκε από τον Laron το 1966. Εκτοτε κάθε περίπτωση μεγάλου βαθμού αντίστασης των περιφερικών ιστών στην GH με εκσεσημασμένη διαταραχή της αύξησης του οργανισμού, παρά τα φυσιολογικά ή υψηλά επίπεδα της GH στον ορό, χαρακτηρίζεται ως σύνδρομο Laron. Οι έρευνες της μοριακής γενετικής έχουν δείξει ότι η διαταραχή αυτή σχετίζεται κυρίως με μεταλλάξεις του γονιδίου του υποδοχέα της GH.

46
Οι μεταλλάξεις αυτές είτε επηρεάζουν τη σύνδεση της ορμόνης με τον υποδοχέα, είτε το διμερισμό του τελευταίου μετά τη σύνδεσή του με την ορμόνη. Η διαταραχή κληρονομείται με τον αυτοσωμικό επικρατούντα χαρακτήρα και σήμερα πιστεύουμε ότι παρουσιάζεται με ποικιλία φαινοτύπων. Στο σύνδρομο Laron παρατηρούνται εξαιρετικά χαμηλά επίπεδα του IGF-1 και της πρωτεΐνης 3 που συνδέεται με τους IGF στην κυκλοφορία.

47
Αυτό σημαίνει ότι οι ίδιοι οι ασθενείς προσφέρουν μία in vitro μέθοδο προσδιορισμού της δραστικής GH, και ότι στο σύνδρομο Laron είτε η GH, η οποία βρίσκεται σε μεγάλες συγκεντρώσεις στην κυκλοφορία, είναι βιολογικά αδρανής, είτε παρουσιάζεται ανεπάρκεια στο επίπεδο του υποδοχέα και της σηματοδοτικής οδού. Εφόσον με τη χορήγηση εξωγενούς βιοδραστικής GH σε ασθενείς με σύνδρομο Laron τα επίπεδα του IGF-1 δεν αυξάνονται, φαίνεται ότι ισχύει μάλλον η δεύτερη περίπτωση.

48
Αυτό επιβεβαιώθηκε με την κλωνοποίηση του γονιδίου του υποδοχέα της GH και την εντόπιση διαφόρων μεταλλάξεων του γονιδίου σε ασθενείς με σύνδρομο Laron. Το γονίδιο του υποδοχέα της GH περιέχει 10 εξόνια. Σχετικά νωρίς, δηλαδή το 1989, εντοπίστηκαν απαλείψει μεγάλων περιοχών του γονιδίου στα εξόνια 3, 5 και 6 σε δύο ασθενείς με σύνδρομο Laron. Εφόσον τα εξόνια 2-7 κωδικοποιούν το σηματοδοτικό πεπτίδιο και την εξωκυττάρια περιοχή του υποδοχέα της GH, δηλαδή τμήματα που επηρεάζουν τη σύνδεση της ορμόνης, είναι λογικό οι προαναφερθείσες μεταλλάξεις να οδηγούν σε αντίσταση των περιφερικών ιστών στην GH.

50
Μετά την αρχική αυτή μελέτη, οι επόμενες μεταλλάξεις που εντοπίστηκαν στο γονίδιο ήταν σημειακές μεταλλάξεις, δηλαδή αφορούσαν αντικατάσταση ενός μόνο νουκλεοτιδίου. Σήμερα είναι γνωστές διάφορες μεταλλάξεις των εξονίων 4 και 7 (μεταλλάξεις μετατόπισης αναγνωστικού πλαισίου, μεταλλάξεις χωρίς νόημα ή λάθος νόημα, μεταλλάξεις που δημιουργούν πρόβλημα στην ορθή συναρμολόγηση – splicing –του γονιδίου), οι οποίες επίσης διαταράσσουν τη λειτουργία του εξωκυττάριου τμήματος του υποδοχέα.
, οι οποίες επίσης διαταράσσουν τη λειτουργία του εξωκυττάριου τμήματος του υποδοχέα..")
51
Το εξωκυττάριο τμήμα του υποδοχέα μπορεί να ανιχνευθεί σην κυκλοφορία, σαν μία πρωτεΐνη που δεσμεύει την αυξητική ορμόνη (GH binding protein - GHBP). Η ύπαρξη φυσιολογικών επιπέδων της GHBP σε ασθενείς με πιθανό σύνδρομο Laron, δείχνει ότι το πρόβλημα εντοπίζεται σε κάποιο σημείο μετά τη σύνδεση της ορμόνης με τον υποδοχέα. Η διαπίστωση λοιπόν ότι οι ασθενείς με φυσιολογικά επίπεδα GHBP έχουν κάπως ηπιότερο φαινότυπο αποκτά ιδιαίτερη σημασία.

52
Μάλιστα το 20-25% των ασθενών με σύνδρομο Laron ανήκουν σε αυτή την κατηγορία. Π.χ έχει αποδειχθεί ότι μία μετάλλαξη χωρίς νόημα στο εξωκυττάριο τμήμα του υποδοχέα, αλλά όχι στη θέση σύνδεσης της GH, δημιουργει προβλήματα στο σχηματισμό διμερών του υποδοχέα, χωρίς να επηρεάζει τη σύνδεση του με τη GH.

54
Επιπλέον έχουν βρεθεί μεταλλάξεις των εξονίων 8-10, οι οποίες επηρεάζουν το διαμεμβρανικό και το ενδοκυττάριο τμήμα του υποδοχέα διαταράσσοντας την αλληλεπίδραση του υποδοχέα με την JAK-2 και κατά συνέπεια την ενδοκυττάρια μετάδοση του σήματος. Από την άλλη πλευρά υπάρχουν ασθενείς με σύνδρομο Laron, στους οποίους δεν εντοπίζονται μεταλλάξεις του γονιδίου του υποδοχέα.

55
Στις περιπτώσεις αυτές είναι πιθανό το πρόβλημα να εντοπίζεται σε γονίδια πρωτεϊνών που συμμετέχουν στην περαιτέρω μετάδοση του σήματος. Η μερική απαλείψεις (deletion) του γονιδίου που κωδικοποιεί τον IGF-1 (γεγονός που καθιστά τον άξονα GH-IGF-1 αδρανή) έχει συσχετιστεί με μία κατάσταση που χαρακτηρίζεται από βαριά ενδομήτριο καθυστέρηση της αύξησης, διαταραχές της αύξησης μετά τον τοκετό, νευροαισθητήριο βαρηκοΐα, αντίσταση των περιφερικών ιστών στην GH και μέτρια διανοητική καθυστέρηση.
 του γονιδίου που κωδικοποιεί τον IGF-1 (γεγονός που καθιστά τον άξονα GH-IGF-1 αδρανή) έχει συσχετιστεί με μία κατάσταση που χαρακτηρίζεται από βαριά ενδομήτριο καθυστέρηση της αύξησης, διαταραχές της αύξησης μετά τον τοκετό, νευροαισθητήριο βαρηκοΐα, αντίσταση των περιφερικών ιστών στην GH και μέτρια διανοητική καθυστέρηση..")
56
Δομή των υποδοχέων: Η δεύτερη μεγάλη ομάδα υποδοχέων περιλαμβάνει τους υποδοχείς που συνδέονται με τις πρωτεΐνες G, οι οποίες βρίσκονται στη εσωτερική επιφάνεια της κυτταρικής μεμβράνης. Η σύνδεση αυτή οδηγεί στο σχηματισμό δεύτερων αγγελιοφόρων, όπως η κυκλική μονοφωσφορική αενοσίνη (cAMP) και η 1, 4, 5-τριφωσφορική ινοσιτόλη (IP3). Πρόκειται για τη μεγαλύτερη ομάδα υποδοχέων της κυτταρικής μεμβράνης αφού αριθμεί περίπου 140 διαφορετικά είδη.
 και η 1, 4, 5-τριφωσφορική ινοσιτόλη (IP3). Πρόκειται για τη μεγαλύτερη ομάδα υποδοχέων της κυτταρικής μεμβράνης αφού αριθμεί περίπου 140 διαφορετικά είδη..")
57
Έχουν περιγραφεί και άλλοι υποδοχείς πλην των ορμονικών που συνδέονται με πρωτεΐνες G, π.χ οι υποδοχείς του γλουταμικού, της θρομβίνης, οι οσφρητικοί και οι οπτικοί υποδοχείς. Υπάρχει μία χαρακτηριστική δομική διαφορά από τους υποδοχείς κινάσης τυροσίνης, η οποία εντοπίζεται στο διαμεμβρανικό τμήμα. Το διαμεμβρανικό τμήμα των GPCRs έχει μία οφιοειδή μορφολογία και διασχίζει επτά φορές τη διπλοστιβάδα λιπιδίων της κυτταρικής μεμβράνης.

58
Οι υδρόφοβες διαμεμβρανικές έλικες του μορίου δεν ποικιλλουν (έχουν σταθερή αλληλουχία αμινοξέων) μεταξύ των διαφόρων υποδοχέων και μπορούν να διαταχθούν έτσι ώστε να σχηματίζουν ένα υδρόφοβο δίαυλο στην κυτταρική μεμβράνη. Το μέγεθος των υποδοχέων κυμαίνεται: ο υποδοχέας της ορμόνης που προκαλεί έκκριση γοναδοτροφίνων αριθμεί μόνο 337 αμινοξέα, ενώ ο υποδοχέας ανίχνευσης εξωκυττάριων ιόντων ασβεστίου αριθμεί 1085 αμινοξέα.

59
Η εξωκυττάρια περιοχή του τελευταίου υποδοχέα (613 αμινοξέα) έχει μικρή συγγένεια (affinity) για τα ιόντα ασβεστίου σε σύγκριση με την υψηλή συγγένεια των ορμονικών υποδοχέων για τις αντίστοιχες ορμόνες. Αυτό φαίνεται να οφείλεται στη συγκέντρωση του ασβεστίου, η οποία είναι πολύ μεγαλύτερη από αυτή των ορμονών.

60
Οι πρωτεΐνες G παίζουν κεντρικό ρόλο στην κινητοποίηση του κυττάρου μετά τη διέγερση ενός GPCR υποδοχέα. Οι πρωτεΐνες G ανήκουν σε μία υπεροικογένεια ρυθμιστικών πρωτεϊνών που λειτουργούν ως μοριακοί διακόπτες, συνδεόμενες με μόρια GTP τα οποία υδρολύονται σε GDP. Οι πρωτεΐνες G είναι είναι από τα μεγαλύτερα μέλη της οικογένειας αυτής, ενώ η πρωτεΐνη Ras είναι από τις μικρότερες G πρωτεΐνες.

61
Η σύνδεση της ορμόνης με έναν GPCR υποδοχέα οδηγεί σε αλλαγή της στερεοδιομής του μορίου, η οποία αφορά και το διαμεμβρανικό και το ενδοκυττάριο τμήμα του υποδοχέα, με αποτέλεσμα την αλληλεπίδραση αυτού με τις G πρωτεΐνες. Οι ενδοκυττάριες αγκύλες και το καρβοξυτελικό άκρο του υποδοχέα έρχονται σε επαφή με ειδικές περιοχές του συμπλέγματος των G πρωτεϊνών. Αυτό οδηγεί στην απελευθέρωση των μορίων του GDP από τη G πρωτεΐνη και στη δέσμευση μορίων GTP.

62
Οι διεγερθείσες G πρωτεΐνες υφίστανται σημαντικές δομικές τροποποιήσεις του μορίου τους, οι οποίες τελικά επηρεάζουν τη δραστηριότητα της αδενυλικής κυκλάσης ή της φωσφολιπάσης C (PLC) της κυτταρικής μεμβράνης.
 της κυτταρικής μεμβράνης.")
63
Σε κατάσταση ηρεμίας, οι G πρωτεΐνες είναι ετεροτριμερή μόρια που αποτελούνται από α, β και γ υπομονάδες. Το μοριακό τους βάρος είναι περίπου 90000. Οι υπομονάδες α,β και γ έχουν μοριακά βάρη 45000, 35000 και 5000 αντιστοίχως. Στην πράξη οι υπομονάδες β και γ είναι τόσο ισχυρά συνδεδεμένες ώστε λειτουργικά το μόριο διακρίνεται σε δύο τμήματα το Gα και το Gβ/γ.

64
Όταν δεν παρατηρείται ενεργοποίηση κάποιου GPCR (απουσία της αντίστοιχης ορμόνης), οι πρωτεΐνες G απλά «περιμένουν» στο εσωτερικό της κυτταρικής μεμβράνης (αποτελώντας, όπως λέγεται, τη «δεξαμενή των G πρωτεϊνών). Όταν κάποιος GPCR διεγερθεί από την αντίστοιχη ορμόνη, συνδέεται με μία G πρωτεΐνη, γεγονός που οδηγεί σε αλλαγή της στερεοδομής της α υπομονάδας, αποδέσμευση του GDP και αντικατάσταση του από GTP.

65
Η ανταλλαγή αυτή των νουκλεοτιδίων γουανίνης οδηγεί με τη σειρά της στην αποδέσμευση της α- υπομονάδας από το ετεροτριμερές σύμπλεγμα της G πρωτεΐνης. Η αποσπασθείσα α-υπομονάδα στη συνέχεια συνδέεται με ένα άλλο μόριο με δραστηριότητα αδενυλικής κυκλάσης ή PLC. G υπομονάδες. Αν και έχουν περιγραφεί περισσότερες από 20 ισομερείς μορφές της Gα- υπομονάδας, μπορούμε να τις ταξινομήσουμε σε 4 κυρίως ομάδες ανάλογα με τη λειτουργία τους.

66
Οι ομάδες αυτές είναι: οι G s α και G i α, οι οποίες διεγείρουν και αναστέλλουν αντιστοίχως της αδενυλική κυκλάση, η G q α, η οποία διεγείρει διαύλους ιόντων. Κάθε GPCR μπορεί να αλληλεπιδρά με μία ή περισσότερες από τις παραπάνω ομάδες G πρωτεϊνών. Περισσότεροι από τους μισούς GPCRs αλληλεπιδρούν με διάφορες ομάδες Gα μορίων, γεγονός που φαίνεται σε πρώτη άποψη αντιφατικό, αφού έτσι θα εκινητοποιούντο αντιτιθέμενα συστήματα ενδοκυττάριων δεύτερων αγγελιαφόρων.

67
Tο φαινόμενο αυτό οφείλεται στην ύπαρξη διαφορετικών υποτύπων GPCRs. Στην περίπτωση της καλσιτονίνης π.χ το κύτταρο εκφράζει διαφορετικούς GPCR ανάλογα με τη φάση του κυτταρικού κύκλου στην οποία βρίσκεται. Εκτός αυτού, οι GPCRs μπορεί να συνδέεται με διαφορετική Gα-υπομονάδα ανάλογα με η συγκέντρωση της ορμόνης.

68
Π.χ. οι υποδοχείς της TSH, της καλσιτονίνης και των LH/hCG διεγείρουν την αδενυλική κυκλάση σε χαμηλές συγκεντρώσεις της αντίστοιχης ορμόνης, ενώ σε μεγαλύτερες συγκεντρώσεις επιστρατεύουν την G q α, η οποία διεγείρει την PLC. Μελέτες τη δομής του μορίου, στις οποίες χρησιμοποιήθηκαν χιμαιρικές Gα-υπομονάδες και μιτογόνες ουσίες, ή υπομονάδες με απάλειψη αμινοξέων του καρβοξυτελικού άκρου, έδειξαν ότι η περιοχή αυτή του μορίου καθορίζει ποιες Gα-υπομονάδες θα επιστρατευθούν από τον διεγερθέντα GPCR.

69
Ουσιαστικά τα δύο άκρα (αμινοτελικό και καρβοξυτελικό της Gα-υπομονάδας δεν απέχουν μεταξύ τους και είναι στραμμένα προς την κυτταρική μεμβράνη, όπου βρίσκεται το ετεροδιμερές Gβ/γ, ο GPCR και τα καταλυτικά μόρια της αδενυλικής κυκλάσης και της PLC. Η σύνδεση της Gα με την κυτταρική μεμβράνη επιτυγχάνεται χάρη σε χημικά τροποποιημένα λιπίδια του καρβοξυ- και αμινοτελικού άκρου καθώς και άλλους παράγοντες.

70
Ποιές Gα-υπομονάδες θα συνδεθούν με τον GPCR καθορίζεται και από τα δομικά χαρακτηριστικά του ενδοκυττάριου τμήματος του υποδοχέα. Έχει διαπιστωθεί π.χ ότι η τρίτη ενδοκυττάρια αγκύλη και το καρβοξυτελικό άκρο του μορίου παίζουν καθοριστικό ρόλο στη σύζευξη των β- αδρενεργικών υποδοχέων με τις G s α- υπομονάδες και των υποδοχέων αγγειοτενσίνης II με τις G q α υπομονάδες.

71
Επιπλέον έχει βρεθεί ότι η προοδευτική απομάκρυνση αμινοξέων από το καρβοξυτελικό άκρο του υποδοχέα της PTH μειώνει την εκλεκτικότητα του υποδοχέα για συγκεκριμένες Gα-υπομονάδες. Με τη βοήθεια πολυπεπτιδίων ομόλογων με την ενδοκυττάρια περιοχή του υποδοχέα έχουμε πετύχει διέγερση των Gα-υπομονάδων. Από την άλλη πλευρά, έχουν συντεθεί πολυπεπτίδια, τα οποία εμποδίζουν τη σύνδεση των υποδοχέων με τις Gα-υπομονάδες, δηλαδή αναστέλλουν εκλεκτικά την ενδοκυττάρια σηματοδοτική οδό.

72
Οι Gα-υπομονάδες έχουν εγγενή δράση GTPάσης, η οποία αποκόπτει μία φωσφορική ομάδα από το GTP, οπότε προκύπτει ένα σύμπλεγμα Gα-GDP. Με αυτόν τον τρόπο οι G-πρωτεΐνες αυτοπεριορίζουν τη δράση τους και παύουν να διεγείρουν τις καταλυτικές υπομονάδες (αδενυλικής κυκλάσης και PLC). Η υδρόλυση του GTP απελευθερώνει την Gα- υπομονάδα από το καταλυτικό μόριο και την συνδέει εκ νέου με το σύμπλεγμα Gβ/γ.
. Η υδρόλυση του GTP απελευθερώνει την Gα- υπομονάδα από το καταλυτικό μόριο και την συνδέει εκ νέου με το σύμπλεγμα Gβ/γ..")
73
Το ανασχηματισθέν ετεροτριμερές επιστρέφει στη «δεξαμενή των G- πρωτεϊνών» της κυτταρικής μεμβράνης. Έτσι το σύμπλεγμα της G-πρωτεΐνης ανακυκλώνεται και είναι έτοιμο να διεγερθεί εκ νέου από κάποιον GPCR.

74
Υπάρχουν αρκετά παραδείγματα ενδοκρινοπαθειών, οι οποίες οφείλονται σε μεταλλάξεις που προκαλούν υπερ- ή υπο- λειτουργία των πρωτεϊνών G και των GPC υποδοχέων. Ο ψευδοϋποπαραθυρεοειδισμός και η κληρονομική οστεοδυστροφία του Albright σχετίζονται με υπολειτουργία των G s α- υπομονάδων ενώ το σύνδρομο Mc-Cune- Albright και ορισμένες περιπτώσεις μεγαλακρίας σχετίζονται με υπερλειτουργία των G s α-υπομονάδων.

75
Ο περίεργος συνδυασμός ευρημάτων που χαρακτηρίζει τις παραπάνω παθήσεις οφείλεται στο γεγονός ότι επηρεάζεται ο τρόπος δράσης των τροφικών ορμονών που ρυθμίζουν τη λειτουργία και τον κυτταρικό πολλαπλασιασμό διάφορων ενδοκρινών αδένων προκαλώντας αύξηση του ενδοκυττάριου cAMP. Από την άλλη πλευρά, ο νεφρογενής άποιος διαβήτης φαίνεται να οφείλεται σε μετάλλαξη του ίδιου του GPCR.

76
Μεταλλάξεις σε επίπεδο γεννητικών κυττάρων της Xq2.8 περιοχής, όπου κωδικοποιείται ο V2 υποδοχέας, οδηγούν σε υπολειτουργία του υποδοχέα, με αποτέλεσμα να μην επιτυγχάνεται συμπύκνωση των ούρων παρά τα υψηλά επίπεδα της αντιδιουρητικής ορμόνης στην κυκλοφορια. Αντιθέτως η οικογενής εμφάνιση πρόωρης ήβης σε άρρενες (testotoxicosis), μία μορφή αυτόνομης υπερλειτουργίας των ενδοκρινών αδένων, οφείλεται σε μεταλλάξεις «χωρίς νόημα» του γονιδίου του LH υποδοχέα στα γεννητικά κύτταρα των γονέων, οι οποίες οδηγούν σε υπερλειτουργία του υποδοχέα.
, μία μορφή αυτόνομης υπερλειτουργίας των ενδοκρινών αδένων, οφείλεται σε μεταλλάξεις «χωρίς νόημα» του γονιδίου του LH υποδοχέα στα γεννητικά κύτταρα των γονέων, οι οποίες οδηγούν σε υπερλειτουργία του υποδοχέα..")
77
Έτσι ενώ οι GPCRs φυσιολογικά βρίσκονται σε κατάσταση ηρεμίας, μία μετάλλαξη μπορεί να επηρεάσει τις σχέσεις των ελίκων του μορίου και να διεγείρει τον υποδοχέα, κάτι που κανονικά συμβαίνει κατόπιν σύνδεσης του μορίου με την αντίστοιχη ορμόνη. Σε τοξικά αδενώματα του θυρεοειδούς έχουν αναφερθεί μεταλλάξεις του ενδομεμβρανικού τμήματος του υποδοχέα της TSH και μία μετάλλαξη του γονιδίου που κωδικοποιεί την G s α-υπομονάδα έχει συσχετιστεί με αυτόνομους όζους του θυρεοειδούς. Από την άλλη πλευρά η μη ανταπόκριση του υποδοχέα στην TSH έχει αποδοθεί σε μεταλλάξεις του εξωκυττάριου τμήματος του μορίου.

78
cAMP. Η G s α-υπομονάδα διεγείρει την αδενυλική κυκλάση της κυτταρικής μεμβράνης, η οποία καταλύει τη μετατροπή του ATP σε cAMP, ένα δεύτερο αγγελιαφόρο με ισχυρή ενδοκυττάρια δράση. Το κυκλικό αυτό νουκλεοτίδιο διεγείρει με τη σειρά του μία πρωτεϊνική κινάση η οποία εξαρτάται από το cAMP (PKA) και ρυθμίζει διάφορες κυτταρικές λειτουργίες.
 και ρυθμίζει διάφορες κυτταρικές λειτουργίες..")
79
Έχουν περιγραφεί τουλάχιστον 10 ισομερή της αδενυλικής κυκλάσης. Το μόριο της αδενυλικής κυκλάσης αποτελείται από 14 τομείς (domains). Παρατηρούνται δύο διαμεμβρανικές ομάδες που αποτελούνται από έξι τομείς έκαστη και δύο ενδοκυττάριοι τομείς. Τα ισομερή διαφέρουν ως προς την αλληλεπίδραση τους με τους αναστολείς του ενζύμου, όπως είναι η G i α-υπομονάδα και το σύμπλεγμα Gβ/γ.
. Παρατηρούνται δύο διαμεμβρανικές ομάδες που αποτελούνται από έξι τομείς έκαστη και δύο ενδοκυττάριοι τομείς. Τα ισομερή διαφέρουν ως προς την αλληλεπίδραση τους με τους αναστολείς του ενζύμου, όπως είναι η G i α-υπομονάδα και το σύμπλεγμα Gβ/γ..")
80
Το cAMP συνδέεται με μία κατασταλτική υπομονάδα της PKA, η οποία στη συνέχεια αποσπάται απότο ολοένζυμο, αποκαλύπτοντας το ενεργό κέντρο του ενζύμου, το οποίο φωσφορυλιώνει σερίνης και θρεονίνης. Με τον τρόπο αυτό οι G s α-υπομονάδες ρυθμίζουν ζωτικές μεταβολικές οδού, μέσω του cAMP και των PKAs, όπως αυτές της λιπόλυσης, της γλυκογονόλυσης και της στεροειδογένεσης.

81
Η PKA φωσφορυλιώνει τον GREB (cAMP response element binding protein), έναν παράγοντα που επηρεάζει τη μεταγραφική δραστηριότητα του κυττάρου. Ο GREB στη συνέχεια εισέρχεται στον πυρήνα, όπου συνδέεται με μία μικρή παλίνδρομη αλληλουχία νουκλεοτιδίων, γνωστή ως «στοιχείο που διεγείρεται από το cAMP» (cAMP response element, GRE).
..")
82
Τα GRE εντοπίζονται στις περιοχές των εκκινητών των γονιδίων, η μεταγραφή των οποίων ρυθμίζεται από το cAMP. Με τον τρόπο αυτό το cAMP αυξάνει τη μεταγραφή συγκεκριμένων γονιδίων. Η σωματοστατίνη επάγει τη μεταγραφή γονιδίων με τον παραπάνω μηχανισμό. Ο βασικότερος μηχανισμός τερματισμού της δράσης του cAMP είναι αυτός των φωσφοδιεστερασών (PDEs).
..")
83
Οι PDEs διεγείρονται με διάφορους τρόπους, συμπεριλαμβανομένης της άμεσης φωσφορυλίωσης από την PKA. Οι PDEs υδρολύουν ταχέως το cAMP σε αδρανές 5’AMP. Επιπλέον το cAMP κινητοποιεί μόρια όπως η PKA, η PKC και οι πρόσφατα ανακαλυφθείσες κινάσες του υποδοχέα (GRKs), τα οποία πιθανώς φωσφορυλιώνουν αμινοξέα σερίνης και θρεονίνης στις ενδοκυττάριες αγκύλες και το καρβοξυτελικό άκρου του GPCR, γεγονός που οδηγεί στην απευαισθητοποίηση του υποδοχέα.
, τα οποία πιθανώς φωσφορυλιώνουν αμινοξέα σερίνης και θρεονίνης στις ενδοκυττάριες αγκύλες και το καρβοξυτελικό άκρου του GPCR, γεγονός που οδηγεί στην απευαισθητοποίηση του υποδοχέα..")
84
Σε ασθενείς με χρόνια καρδιακή ανεπάρκεια έχει περιγραφεί αυξημένη συγκέντρωση μία συγκεκριμένης GBK, η οποία φωσφορυλιώνει τους β- αδρενεργικούς υποδοχείς.

85
Περισσότερες από 20 εξωκυττάριες ρυθμιστικές ουσίες, στις οποίες περιλαμβάνονται η TRH, η GnRH και η ωκυτοκίνη, διεγείρουν τα κύτταρα-στόχους μέσω GPC υποδοχέων που επιστρατεύουν πρωτεΐνες G με G q α-υπομονάδες. Η G q α-υπομονάδα διεγείρει τη φωσφολιπάση C (PLC) της κυτταρικής μεμβράνης. Το ένζυμο αυτό (έχουν παρατηρηθεί τρεις ισομερείς μορφές: οι α,β και γ) καταλύει την αντίδραση.
 της κυτταρικής μεμβράνης. Το ένζυμο αυτό (έχουν παρατηρηθεί τρεις ισομερείς μορφές: οι α,β και γ) καταλύει την αντίδραση..")
86
PIP 2 → DAG + IP 3 Όπου PIP 2 είναι 4,5 διφωσφορική ινοσιτόλη και DAG είναι η διακυλογλυκερόλη. Η PIP 2 είναι ένα δευτερεύον φωσφολιπίδιο της κυτταρικής μεμβράνης. Το ποσοστό της είναι μικρότερο από το 1% των συνολικών φωσφολιπιδίων της κυτταρικής μεμβράνης.

87
Η DAG, με τη βοήθεια της φωσφατιδυλοσερίνης, διεγείρει την πρωτεϊνική κινάση C (PKC) της κυτταρικής μεμβράνης, ενώ η IP 3,απελευθερώνεται στο κυτταρόπλασμα, όπου συνδέεται με διαύλους- υποδοχείς του ενδοπλασματικού δικτύου, οι οποίοι κινητοποιούν το ενδοκυττάριο ασβέστιο. Αυτό οδηγεί ταχύτατα σε δεκαπλασιασμό των επιπέδων Ca ++ στο κυτταρόπλασμα (η φυσιολογική συγκέντρωση είναι περίπου 0,1 μΜ).
..")
88
Τα ιόντα Ca ++ κινητοποιούν αρκετά ένζυμα, συμπεριλαμβανομένων της πρωτεϊνικής κινάσης που διεγείρεται από την καλμοδουλίνη και ορισμένων ισομερών της PKC. Μάλιστα το όνομα PKC προέρχεται από την εξάρτηση του μορίου αυτού από το Ca ++. Το Ca ++ επίσης κινητοποιεί τη φωσφολίπαση Α 2, η οποία προκαλεί απελευθέρωση αραχιδονικού οξέος από τα φωσφολιπίδια και γενικότερα οδηγεί στην παραγωγή εικοσανοειδών με ισχυρή δράση στους τοπικούς ιστούς.

89
Στα εικοσανοειδή περιλαμβάνονται ιοι θρομβοξάνες, τα λευκοτριένια, οι λιποξίνες και οι προσταγλανδίνες. Οι προσταγλανδίνες δρουν σε αυτοκρινείς και παρακρινείς μηχανισμούς, αυξάνοντας ή παρατείνοντας την απάντηση των κυττάρων στο αρχικό ορμονικό ερέθισμα. Τα ιόντα Ca ++ κινητοποιούν και την κυτταροπλασματική γουανυλική κυκλάση, ένα ένζυμο που καταλύει την αντίδραση σχηματισμού της κυκλικής μονοφωσφορικής γουανοσίνης (cGMP).
..")
90
Το κολπικό νατριουρητικό πεπτίδιο δρα μέσω υποδοχέων που διεγείρουν τη γουανυλική κυκλάση. Η αύξηση των ελεύθερων ιόντων Ca ++ στο κυτταρόπλασμα είναι παροδική. Η κινητοποίηση του ασβεστίου διακόπτεται με διάφορους μηχανισμούς. Π.χ η PLC-β μπορεί να επιταχύνει την υδρόλυση του GTP, απενεργοποιώντας το σύμπλεγμα G q α/GTP, δρώντας σαν «πρωτεΐνη που διεγείρει τις GTP-άσες» (GTPase-activating protein - GAP).
..")
91
Τα μόρια των στεροειδών και των θυρεοειδικών ορμονών δεν παρουσιάζουν χημική συγγένεια μεταξύ τους, ωστόσο συνδέονται με πρωτεϊνικούς υποδοχείς που ανήκουν στην ίδια υπεροικογένεια ενδοκυττάριων υποδοχέων. Δηλαδή οι υποδοχείς των στεροειδών και των θυρεοριδικών ορμονών μοιάζουν δομικά μεταξύ τους. Οι παραπάνω ορμόνες, όντας υδρόφοβες, διαχέονται ελεύθερα διά της κυτταρικής μεμβράνης των κυττάρων-στόχων για να συνδεθούν με τους υποδοχείς τους που βρίσκονται στο κυτταρόπλασμα και στον πυρήνα.

92
Οι υποδοχείς αυτοί επηρεάζουν τη μεταγραφή συγκεκριμένων γονιδίων (transcription factors) αλληλεπιδρώντας με περιοχές που βρίσκονται κοντά στους εκκινητές των γονιδίων (promotors). Η βιολογική δράση των στεροειδών και θυρεοειδικών ορμονών καθυστερεί να εμφανιστεί, σε αντίθεση με την άμεση απάντηση που επιτυγχάνεται με τις ορμόνες που συνδέονται με τους υποδοχείς της κυτταρικής μεμβράνης και τα συστήματα των δεύτερων αγγελιαφόρων.

93
Αυτό οφείλεται στο γεγονός ότι οι στεροειδείς και θυρεοειδικές ορμόνες δρουν μέσω της επαγωγής συγκεκριμένων γονιδίων τα οποία μεταγράφονται σε mRNA και μεταφράζονται σε πρωτεΐνες, διαδικασίες που αργούν να ολοκληρωθούν. Ετσι παρατηρείται μία λανθάνουσα περίοδος μεταξύ της έκθεσης του κυττάρου-στόχου στην ορμόνη και της in vitro βιολογικής απάντησης του κυττάρου.

94
Η υπεροικογένεια αυτή αριθμεί περισσότερα από 150 είδη υποδοχέων. Ωστόσο στην πλειονότητα τους είναι «ορφανοί», δηλαδή δεν έχει βρεθεί το αντίστοιχο μόριο διεγέρτης (ligand).
..")
95
Κάθε υποδοχέας αποτελείται από μία πολυπεπτιδική αλυσίδα, στην οποία διακρίνονται τρία χαρακτηριστικά τμήματα: 1. Μία περιοχή που συνδέεται εκλεκτικά με την αντίστοιχη ορμόνη. Το καρβοξυτελικό άκρο περιέχει μία περιοχή (AF2) που παίζει ρόλο στην ορμονοεξαρτώμενη επαγωγή των γονιδίων. Η θέση σύνδεσης της ορμόνης είναι ένας υδρόφοβος θύλακας του μορίου.
 που παίζει ρόλο στην ορμονοεξαρτώμενη επαγωγή των γονιδίων. Η θέση σύνδεσης της ορμόνης είναι ένας υδρόφοβος θύλακας του μορίου..")
96
2. Μία γενετικά σταθερή περιοχή σύνδεσης με το DNA. 3. Μία γενετικά ευμετάβλητη αμινοτελική περιοχή που ποικιλλεί τόσο σε σύσταση όσο και σε μήκος από άτομο σε άτομο. Σε μερικούς υποδοχείς η περιοχή αυτή φαίνεται να παίζει ρόλο στη μη ορμονοεξαρτώμενη επαγωγή γονιδίων, μία ενδογενή δηλαδή ικανότητα του μορίου να προάγει τη μεταγραφή ορισμένων γονιδίων.

97
Οι παραπάνω περιοχές λειτουργούν ανεξάρτητα η μία από την άλλη, όπως έχει φανεί από έρευνες με υβριδικούς ή χιμαιρικούς υποδοχείς. Το μοριακό βάρος των υποδοχέων ποικιλλεί από 46kDa στην περίπτωση του Τ 3 υποδοχέα μέχρι και 100kDa στην περίπτωση των υποδοχέων της προγεστερόνης και των αλατοκορτικοειδών.

98
Μεταξύ των υποδοχέων παρατηρείται αξιοσημείωτη ομολογία στην αλληλουχία των αμινοξέων, η οποία φτάνει μέχρι και το 60-90%, γεγονός που αποδεικνύει την εξελικτική συγγένεια των μορίων αυτών. Έχει διατυπωθεί η άποψη ότι οι υποδοχείς αυτοί έχουν σαν κοινο πρόγονό τους το ογκογονίδιο v-erb A. Σε κύτταρα που εκφράζουν τα γονίδια των εν λόγω υποδοχέων παρατηρείται έντονη ανοσοϊστοχημική χρώση των πυρήνων με αντισώματα που εμφανίζουν μεγάλη ειδικότητα για τους υποδοχείς αυτούς.

99
Οι υποδοχείς συνήθως διακρίνονται σε δύο ομάδες με βάση τον τρόπο επαγωγής των γονιδίων και τη δομή τους σε κατάσταση ηρεμίας. Οι στεροειδείς ορμόνες δρουν στους υποδοχείς της ομάδας I ενώ η καλσιτριόλη, το ρετινοϊκό οξύ και η T 3 δρουν στους υποδοχείς της ομάδας II.

100
Η περιοχή αυτή χαρακτηρίζεται από την ύπαρξη δύο «δακτύλων ψευδαργύρου (zinc fingers). Πρόκειται για δύο πολυπεπτιδικές αγκύλες μήκους 10-20 αμινοξέων έκαστη. Σε κάθε αγκύλη δύο κυστεΐνες και δύο ιστιδίνες συνδέονται με ένα ιόν ψευδαργύρου, το οποίο σταθεροποιεί τη συγκεκριμένη δομή. Οι δυό δάκτυλοι απέχουν περίπου 12 αμινοξέα μεταξύ τους.

101
Οι χαρακτηριστικές αυτές δομές είναι απαραίτητες για τη σύνδεση του υποδοχέα με το DNA. Σε κατάσταση ηρεμίας, οι δάκτυλοι ψευδαργύρου των υποδοχέων της ομάδας I είναι κρυμμένοι λόγω σύνδεσης του υποδοχέα με ένα διμερές μόριο πρωτεΐνης θερμικού shock, όπως η hsp70, η hsp90 και άλλες.

102
Αυτά τα συμπλέγματα υψηλού μοριακού βάρους κατανέμονται μεταξύ του κυτταροπλάσματος και του πυρήνα σε αναλογία που ποικιλλεί ανάλογα με την ορμόνη. Π.χ το 90% των υποδοχέων των γλυκοκορτικοειδών βρίσκονται στο κυτταρόπλασμα ενώ η πλειοψηφία των υποδοχέων των ανδρογόνων εντοπίζεται στον πυρήνα του κυττάρου.

103
Η σύνδεση των υποδοχέων με τα διμερή μόρια των hsp, καλύπτει τους δακτύλους ψευδαργύρου και εμποδίζει τη σύνδεση του υποδοχέα με το DNA. Η σύνδεση του στεροειδούς με τον υποδοχέα οδηγεί στην απομάκρυνση του διμερούς hsp και την αποκάλυψη των δακτύλων ψευδαργύρου. Στη συνέχεια τα μόρια των υποδοχέων εισέρχονται στον πυρήνα, σχηματίζουν διμερή και οι τέσσερις συνολικά δάκτυλοι ψευδάργυρου συνδέονται με μία μικρή περιοχή του DNΑ, γνωστή ως «στοιχείο απάντησης σε ορμονικά ερεθίσματα» (hormone response element - HRE) ή στην περίπτωση του T 3 υποδοχέα, «στοιχείο απάντησης στις θυρεοειδικές ορμόνες» (thyroid hormone response element - TRE).
 ή στην περίπτωση του T 3 υποδοχέα, «στοιχείο απάντησης στις θυρεοειδικές ορμόνες» (thyroid hormone response element - TRE)..")
104
Πιστεύεται ότι κάθε δάκτυλος ψευδαργύρου αναγνωρίζει μία ειδική αλληλουχία περίπου 5 νουκλεοτιδίων του HRE. Οι λεπτομέρειες της αλληλεπίδρασης των δακτύλων ψευδαργύρου των πρωτεϊνικών υποδοχέων με τη μείζονα σχισμή του DNA έχουν αποσαφηνιστεί με τη βοήθεια της κρυσταλλογραφικής ανάλυσης με ακτίνες X και της φασματογραφίας των πρωτεϊνών με την τεχνική του NMR (nuclear magnetic resonance).
..")
105
H ειδικότητα του συμπλέγματος ορμόνης/υποδοχέα για ένα συγκεκριμένο HRE αποδίδεται σε ελάχιστα αμινοξέα του υποδοχέα. Αυτά εντοπίζονται σε μία περιοχή που ονομάζεται P Box, η οποία βρίσκεται στη βάση του πρώτου δακτύλου ψευδάργυρου του υποδοχέα των γλυκοκορτικοειδών οδηγεί σε απώλεια της ειδικότητας του, με αποτέλεσμα την επαγωγή γονιδίων, η μεταγραφή των οποίων κανονικά διεγείρεται από τα οιστρογόνα.

106
Συνεπώς στη δομή των υποδοχέων αυτών μπορούμε να εντοπίσουμε δύο περιοχές σημαντικές για την ειδικότητα του μορίου: τη θέση σύνδεσης της ορμόνης και τη θέση σύνδεσης με το DNA. Οι υποδοχείς των θυρεοειδικών ορμονών εντοπίζονται αποκλειστικά στον πυρήνα. Σε κατάσταση ηρεμίας είναι ήδη συνδεδεμένοι με θέσεις του DNA και διεγείρονται όταν η θυρεοειδική ορμόνη συνδεθεί με την αντίστοιχη θέση του μορίου τους.

107
Παρόμοια συμπεριφορά επιδεικνύουν και οι υποδοχείς της καλσιτριόλης και του ρετινοϊκού οξέος, γι΄αυτό και τα τρία αυτά είδη υποδοχέων έχουν ταξινομηθεί στην ομάδα IΙ της υπεροικογένειας των ενδοκυττάριων υποδοχέων. Οι υποδοχείς της T 3 οι οποίοι εξ ορισμού βρίσκονται στον πυρήνα, μπορεί να αναστέλλουν τη μεταγραφή των γονιδίων- στόχων απουσία της αντίστοιχης τους ορμόνης.

108
Αυτό μπορεί να επιτυγχάνεται με τη βοήθεια μίας συν-κατασταλτικής πρωτεΐνης, η οποία εμποδίζει ακόμα και τη στοιχειώδη μεταγραφική δραστηριότητα που επάγει ο εκκινητής, αλλά απομακρύνεται από τον υποδοχέα παρουσία της T 3.

109
Ο διμερισμός του υποδοχέα είναι απαραίτητος για τη σύνδεση του συμπλέγματος ορμόνη- υποδοχέα με το HRE. Οι υποδοχείς της ομάδας I σχηματίζουν ομοδιμερή τα οποία συνδέονται με παλινδρομικά διατεταγμένες αλληλουχίες μήκους 6 νουκλεοτιδίων. Αντιθέτως, οι υποδοχείς της ομάδας II, όπως αυτός της T 3, σχηματίζουν ετεροδιμερή με ελεύθερους υποδοχείς X ρετινοειδών, αν και έχουν περιγραφεί και ομοδιμερή μόρια.

110
Τα διμερή των υποδοχέων συνδέονται με το HRE, το οποίο βρίσκεται πριν το γονίδιο και τον εκκινητή του. Η δραστηριότητα της DNA-εξαρτώμενης- RNA-πολυμεράσης καθορίζεται από τον εκκινητή. Η δράση του τελευταίου ενισχύεται, ή σε ορισμένες περιπτώσεις καταστέλλεται, από το διμερισμένο σύμπλεγμα ορμόνης- υποδοχέα που συνδέεται με το HRE.

111
Τα συμπλέγματα μπορεί επίσης να επιστρατεύουν βοηθητικές πρωτεΐνες που διεγείρουν ή καταστέλλουν τη μεταγραφή. Οι πρωτεΐνες αυτές δρουν ευοδώνοντας ή καταστέλλοντας τη δραστηριότητα του συμπλέγματος έναρξης της μεταγραφής (το σύμπλεγμα σχηματίζεται από την RNA πολυμεράση και άλλους παράγοντες που σχετίζονται με τη μεταγραφή του γονιδίου).
..")
112
Το παραγόμενο mRNA υφίσταται περαιτέρω επεξεργασία κατά την οποία αποκόπτονται τα ιντρόνια και συνενώνονται μεταξύ τους τα εξόνια (splicing). Το τελικό αποτέλεσμα της δράσης των στεροειδών ορμονών είναι η σύνθεση νέων πρωτεϊνών στο κυτταρόπλασμα. Οι στεροειδείς ορμόνες όμως ενισχύουν άμεσα τη μεταγραφή ελάχιστων γονιδίων, οπότε ο αριθμός των παραγόμενων πρωτεϊνών είναι μικρός.

113
Σε ορισμένα κύτταρα παρατηρείται μία δευτερογενής απάντηση που ακολουθεί την πρώιμη αυτή αντιδραση του κυττάρου. Η δευτερογενής απάντηση οφείλεται σε πρωτεΐνες που παράγονται κατά την πρωτογενή αντίδραση του κυττάρου, οι οποίες με τη σειρά τους επάγουν τη μεταγραφική διαφόρων γονιδίων. Έτσι παρατηρείται ενίσχυση και συντονισμός της κυτταρικής απάντησης στο ορμονικό ερέθισμα.

114
Από τη στιγμή που επετεύχθη η απομόνωση του cDNA των πυρηνικών υποδοχέων των ορμονών μέχρι σήμερα, έχουν γίνει εκτεταμένες έρευνες για τον εντοπισμό γονιδιακών μεταλλάξεων και τη συσχέτιση τους με γνωστές ενδοκρινοπάθειες. Έχουν εντοπιστεί μεταλλάξεις, οι οποίες επηρεάζουν αρνητικά τη σύνδεση ορμόνης- υποδοχέα, το διμερισμό των υποδοχέων και τη σύνδεση των διμερών συμπλεγμάτων με τα αντίστοιχα HREs.

115
Εκτός των παραπάνω παραγόντων μεταγραφής, δηλαδή του CREB και των πυρηνικών υποδοχέων των ορμονών, έχει ανιχνευτεί ένας ακόμη παράγοντας, ο οποίος βρέθηκε στα κύτταρα της υπόφυσης και ρυθμίζει την έκφραση των γονιδίων της αυξητικής ορμόνης, της προλακτίνης και της β- υπομονάδας της TSH.

116
Ο παράγοντας αυτός είναι γνωστός ως Pit- 1 και ανήκει στην POU οικογένεια παραγόντων, οι οποίοι είναι σημαντικοί για την ανάπτυξη των θηλαστικών. Έχουν βρεθεί αλληλουχίες του DNA που συνδέονται με τον Pit-1 και εντοπίζονται στην περιοχή των εκκινητών των γονιδίων της αυξητικής ορμόνης, της προλακτίνης και της TSH-β.

117
Αυτό εξηγεί την παράδοξη έλλειψη ειδικότητας της TRH. Ωστόσο, η TRH δεν αυξάνει την έκκριση αυξητικής ορμόνης, εκτός αν το άτομο παρουσιάζει μεγαλακρία, γεγονός που ερμηνεύεται από τις υψηλές συγκεντρώσεις Pit-1 που απαιτούνται για την επαγωγή του γονιδίου της αυξητικής ορμόνης.

118
Έχουν εντοπιστεί ασθενείς με σημειακές μεταλλάξεις του γονιδίου του Pit-1 και μείωση των επιπέδων ή και έλλειψη αυξητικής ορμόνης, προλακτίνης και TSH. Οι ασθενείς αυτοί δεν ανταποκρίνονται στις δοκιμασίες διέγερσης. Η νόσος συσχετίζεται με μειωμένο ανάστημα και αν ο υποθυρεοειδισμός δεν διαγνωστεί έγκαιρα τοτε θα υπαρξουν σοβαρές μαθησιακές δυσκολίες.

119
Υπάρχουν αρκετά παραδείγματα κυττάρων- στόχων που συνθέτουν ειδικά ένζυμα, τα οποία μετατρέπουν τοπικά την ορμόνη σε μία δραστικότερη μορφή. Το νέο αυτό μόριο συνδέεται ισχυρότερα με τους ορμονικούς υποδοχείς του κυττάρου. Π.χ ειδικές 5’δεϊωσινάσες μετατρέπουν τη θυροξίνη σε T 3, η 5 α –αναγωγάση (ή ρεδουκτάση) μετατρέπει την τεστοστερόνη σε διϋδροτεστοστερόνη και η μιτοχονδριακή 1 α- υδροξυλάση των κυττάρων των νεφρικών σωληναρίων μετατρέπει την 25-ΟΗ-βιταμίνη D σε καλσιτριόλη.
 μετατρέπει την τεστοστερόνη σε διϋδροτεστοστερόνη και η μιτοχονδριακή 1 α- υδροξυλάση των κυττάρων των νεφρικών σωληναρίων μετατρέπει την 25-ΟΗ-βιταμίνη D σε καλσιτριόλη..")
120
Αντιθέτως στα νεφρικά κύτταρα, η λειτουργία των οποίων ρυθμίζεται από την αλδοστερόνη, μία 11β-υδροξυστεροειδική-αφυδρογονάση (11β-HSD) αδρανοποιεί την κορτιζόλη μετατρέποντας την στο 11-κετοστεροειδές ανάλογο της, την κορτιζόνη. Η αδρανοποίηση της κορτιζόλης έχει ιδιαίτερη σημασία, αφού η κορτιζόλη συνδέεται με τον υποδοχέα των αλατοκορτικοειδών και η συγκέντρωση της στην κυκλοφορία είναι 2-3 φορές υψηλότερη από αυτή της αλδοστερόνης.

121
Η μη αδρανοποίηση του μορίου θα επέτρεπε την υπερδιέγερση των αλατοκορτικοειδικών υποδοχέων των σωληναριακών κυττάρων. Η 11β- HSD προστατεύει τον «αδηφάγο» αλατοκορτικοειδικό υποδοχέα από τη δράση της κορτιζόλης. Η δυσλειτουργία ή η έλλειψη του παραπάνω ενζύμου οδηγούν στο σύνδρομο «φαινομενικής περίσσειας αλατοκορτικοειδών» (Apparent Miner- alocorticoid Excess - AME), το οποίο χαρακτηρίζεται από υπέρταση και υποκαλιαιμία.
, το οποίο χαρακτηρίζεται από υπέρταση και υποκαλιαιμία..")
















 Παίζει ρόλο σαν μεταφορέας ενέργειας με την μορφή ATP & ADP, σηματοδοτεί γενετικές.>")



