Κατέβασμα παρουσίασης
1
ΠΑΡΑΓΩΓΗ ΑΝΤΙΣΩΜΑΤΩΝ ΑΝΑΓΝΩΡΙΣΗΣ
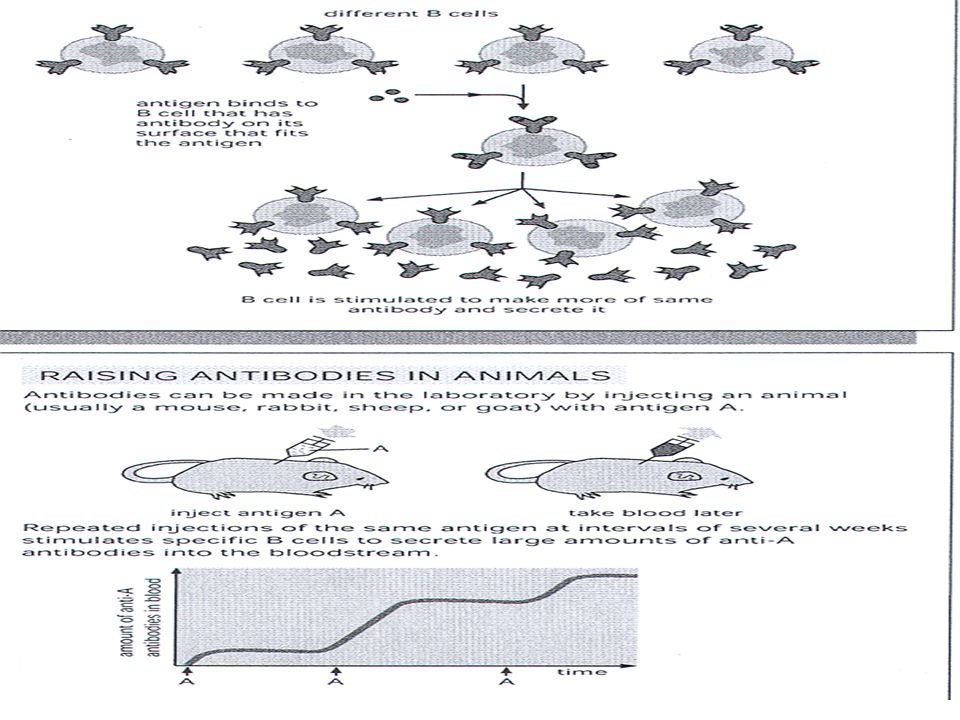
ΜΕΘΟΔΟΙ ΑΝΟΣΟΒΙΟΛΟΓΙΑΣ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΚΑΙ ΑΝΑΓΝΩΡΙΣΗ ΜΙΚΡΟΟΡΓΑΝΙΣΜΩΝ Με την χρησιμοποίηση μιας σειράς αντισωμάτων με γνωστή εξειδίκευση γίνεται δυνατή η ταξινόμηση μικροοργανισμών σε οροομάδες (serogroups). Αυτές, με την χρησιμοποίηση αντισωμάτων που λαμβάνονται ύστερα από ανοσοποιήσεις με αντιγόνα ή ομάδες αντιγόνων που είναι εκ των προτέρων γνωστό ότι διαφέρουν στα διάφορα στελέχη του μικροοργανισμού, υποδιαιρούνται παραπέρα σε ορότυπους (serotypes ή serovars). ΠΑΡΑΓΩΓΗ ΑΝΤΙΣΩΜΑΤΩΝ ΑΝΑΓΝΩΡΙΣΗΣ H οροτυπική αναγνώριση μικροοργανισμών εξαρτάται από τη διαθεσιμότητα αντισωμάτων τα οποία αναγνωρίζουν διαφορές στα αντιγόνα των διαφόρων στελεχών ενός μικροοργανισμού. Τα αντισώματα που χρησιμοποιούνται στην αναγνώριση μπορεί να είναι πολυκλωνικής ή μονοκλωνικής προέλευσης. Πολυκλωνικά αντισώματα Ο πολυκλωνικός αντι-ορός περιέχει μείγμα αντισωμάτων που εκκρίνονται από διαφορετικούς ενεργοποιημένους κλώνους Β λεμφοκυττάρων οι οποίοι, ενώ έχουν ενεργοποιηθεί από το ίδιο αντιγόνο, παράγουν διαφορετικά αντισώματα, τα οποία διαφέρουν ως προς την τάξη αντισωμάτων (IgM, IgG κλπ), την υπό-τάξη (IgG1, IgG2 κλπ.) και τον ιδιότυπο.
. Αυτές, με την χρησιμοποίηση αντισωμάτων που λαμβάνονται ύστερα από ανοσοποιήσεις με αντιγόνα ή ομάδες αντιγόνων που είναι εκ των προτέρων γνωστό ότι διαφέρουν στα διάφορα στελέχη του μικροοργανισμού, υποδιαιρούνται παραπέρα σε ορότυπους (serotypes ή serovars). ΠΑΡΑΓΩΓΗ ΑΝΤΙΣΩΜΑΤΩΝ ΑΝΑΓΝΩΡΙΣΗΣ. H οροτυπική αναγνώριση μικροοργανισμών εξαρτάται από τη διαθεσιμότητα αντισωμάτων τα οποία αναγνωρίζουν διαφορές στα αντιγόνα των διαφόρων στελεχών ενός μικροοργανισμού. Τα αντισώματα που χρησιμοποιούνται στην αναγνώριση μπορεί να είναι πολυκλωνικής ή μονοκλωνικής προέλευσης. Πολυκλωνικά αντισώματα. Ο πολυκλωνικός αντι-ορός περιέχει μείγμα αντισωμάτων που εκκρίνονται από διαφορετικούς ενεργοποιημένους κλώνους Β λεμφοκυττάρων οι οποίοι, ενώ έχουν ενεργοποιηθεί από το ίδιο αντιγόνο, παράγουν διαφορετικά αντισώματα, τα οποία διαφέρουν ως προς την τάξη αντισωμάτων (IgM, IgG κλπ), την υπό-τάξη (IgG1, IgG2 κλπ.) και τον ιδιότυπο.")
2
Ανοσοποιήσεις με το αντιγόνο σε καθαρή μορφή.
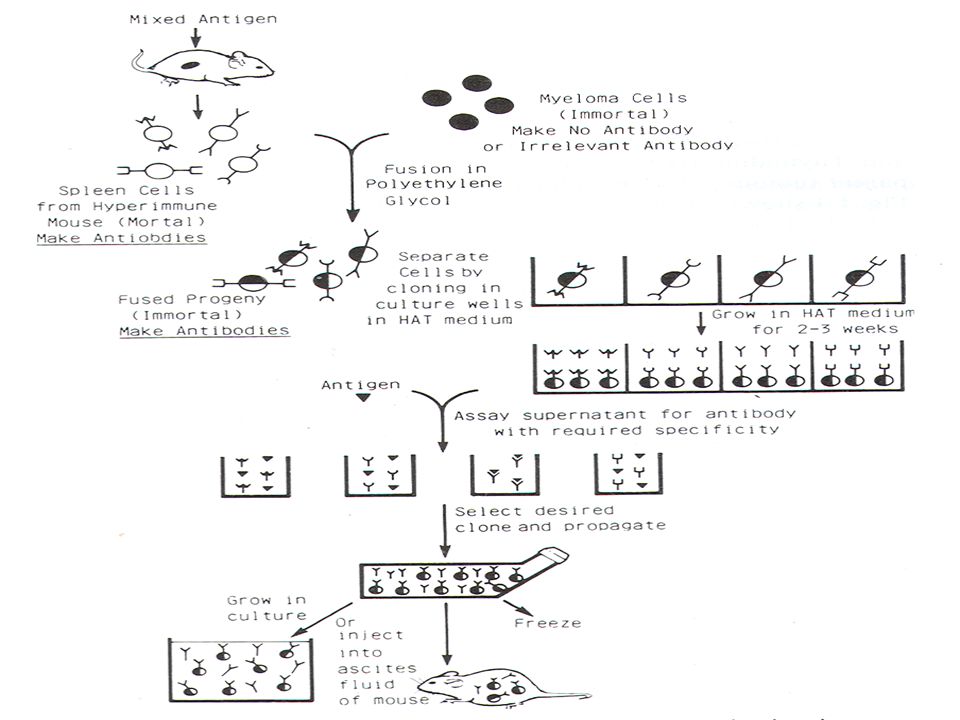
Ο ορός που προκύπτει, αν και περιέχει πολλά είδη αντισωμάτων που όλα αντιδρούν με το ίδιο αντιγόνο ( αλλά με διαφορετικούς επίτοπους) χαρακτηρίζεται ως μονοειδικός ορός (monospecific serum). Στις περισσότερες περιπτώσεις, τα προς ανοσοποίηση αντιγόνα δεν υπάρχουν σε καθαρή μορφή, γι’ αυτό και οι ανοσοποιήσεις αναγκαστικά γίνονται με υποκυτταρικά κλάσματα που περιέχουν πολλά είδη μακρομορίων. Στις περιπτώσεις αυτές ο ορός που προκύπτει περιέχει πληθώρα αντισωμάτων τα οποία αναγνωρίζουν πολλούς επίτοπους διαφορετικών αντιγόνων· ο ορός αυτός χαρακτηρίζεται ως πολυδύναμος ή πολυειδικός ορός (polyvalent serum).Από τον ορό αυτό, πριν χρησιμοποιηθεί στην αναγνώριση, θα πρέπει να αφαιρεθούν όλα τα αντισώματα που δημιουργούν προβλήματα. Αυτό γίνεται με διαδοχικές προσροφήσεις του ορού με παρασκευάσματα που είναι πλούσια σε όλα τα αντιγόνα ανοσοποίησης και από τα οποία απουσιάζει το ειδικό αντιγόνο. Μονοκλωνικά αντισώματα Η μεθοδολογία για την παραγωγή μονοκλωνικών αντισωμάτων, που περιγράφτηκε από τους Kohler και Milstein το 1975, δίνει τη δυνατότητα για την in vitro παραγωγή αντισωμάτων τα οποία είναι ειδικά για ένα συγκεκριμένο επίτοπο ενός αντιγόνου. Τα μονοκλωνικά αντισώματα παράγονται από κύτταρα υβρίδια (υβριδώματα) τα οποία προκύπτουν από την συγχώνευση δύο τύπων κυττάρων, τα B λεμφοκύτταρα από κατάλληλα ανοσοποιημένο ποντικό ή αρουραίο και τα μυελωματικά κύτταρα καλλιεργειών τα οποία προέρχονται από το ίδιο είδος ζώου. Θα πρέπει τα κύτταρα της υβριδωματικής σειράς να καλλιεργηθούν σε μεγάλη κλίμακα ή εναλλακτικά αυτά ενύονται στην ενδοπεριτοναϊκή κοιλότητα ποντικών όπου εκκρίνουν μεγάλη ποσότητα μονοκλωνικού αντισώματος στο ασκητικό υγρό (ascites)
 χαρακτηρίζεται ως μονοειδικός ορός (monospecific serum). Στις περισσότερες περιπτώσεις, τα προς ανοσοποίηση αντιγόνα δεν υπάρχουν σε καθαρή μορφή, γι’ αυτό και οι ανοσοποιήσεις αναγκαστικά γίνονται με υποκυτταρικά κλάσματα που περιέχουν πολλά είδη μακρομορίων. Στις περιπτώσεις αυτές ο ορός που προκύπτει περιέχει πληθώρα αντισωμάτων τα οποία αναγνωρίζουν πολλούς επίτοπους διαφορετικών αντιγόνων· ο ορός αυτός χαρακτηρίζεται ως πολυδύναμος ή πολυειδικός ορός (polyvalent serum).Από τον ορό αυτό, πριν χρησιμοποιηθεί στην αναγνώριση, θα πρέπει να αφαιρεθούν όλα τα αντισώματα που δημιουργούν προβλήματα. Αυτό γίνεται με διαδοχικές προσροφήσεις του ορού με παρασκευάσματα που είναι πλούσια σε όλα τα αντιγόνα ανοσοποίησης και από τα οποία απουσιάζει το ειδικό αντιγόνο. Μονοκλωνικά αντισώματα. Η μεθοδολογία για την παραγωγή μονοκλωνικών αντισωμάτων, που περιγράφτηκε από τους Kohler και Milstein το 1975, δίνει τη δυνατότητα για την in vitro παραγωγή αντισωμάτων τα οποία είναι ειδικά για ένα συγκεκριμένο επίτοπο ενός αντιγόνου. Τα μονοκλωνικά αντισώματα παράγονται από κύτταρα υβρίδια (υβριδώματα) τα οποία προκύπτουν από την συγχώνευση δύο τύπων κυττάρων, τα B λεμφοκύτταρα από κατάλληλα ανοσοποιημένο ποντικό ή αρουραίο και τα μυελωματικά κύτταρα καλλιεργειών τα οποία προέρχονται από το ίδιο είδος ζώου. Θα πρέπει τα κύτταρα της υβριδωματικής σειράς να καλλιεργηθούν σε μεγάλη κλίμακα ή εναλλακτικά αυτά ενύονται στην ενδοπεριτοναϊκή κοιλότητα ποντικών όπου εκκρίνουν μεγάλη ποσότητα μονοκλωνικού αντισώματος στο ασκητικό υγρό (ascites)")
3
που δημιουργείται στο περιτόναιο
που δημιουργείται στο περιτόναιο.Μονοκλωνικό αντίσωμα από ασκητικό υγρό μπορεί να χρησιμοποιηθεί ως έχει, χωρίς καμμιά διαδικασία καθαρισμού, αφού η συγκέντρωσή του κυμαίνεται από 0,5 μέχρι 5 mg/ml, ενώ η συγκέντρωση του στο υπερκείμενο υγρό των υβριδωματικών κυτταρικών σειρών δεν υπερβαίνει τα 25 μg/ml. Για τα μονοκλωνικά αντισώματα που χρησιμοποιούνται στην αναγνώριση στελεχών , θα πρέπει να διασφαλίζεται ότι αυτά αναγνωρίζουν επίτοπους που συναντιώνται σ’όλα τα απομονώματα του προς αναγνώριση μικροοργανισμού.

4
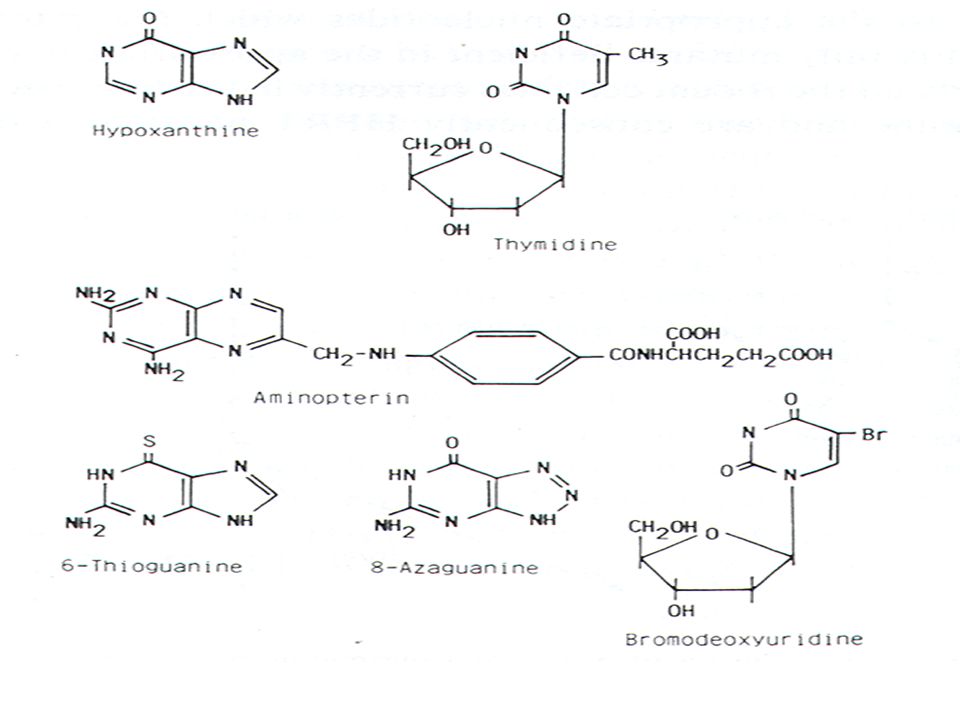
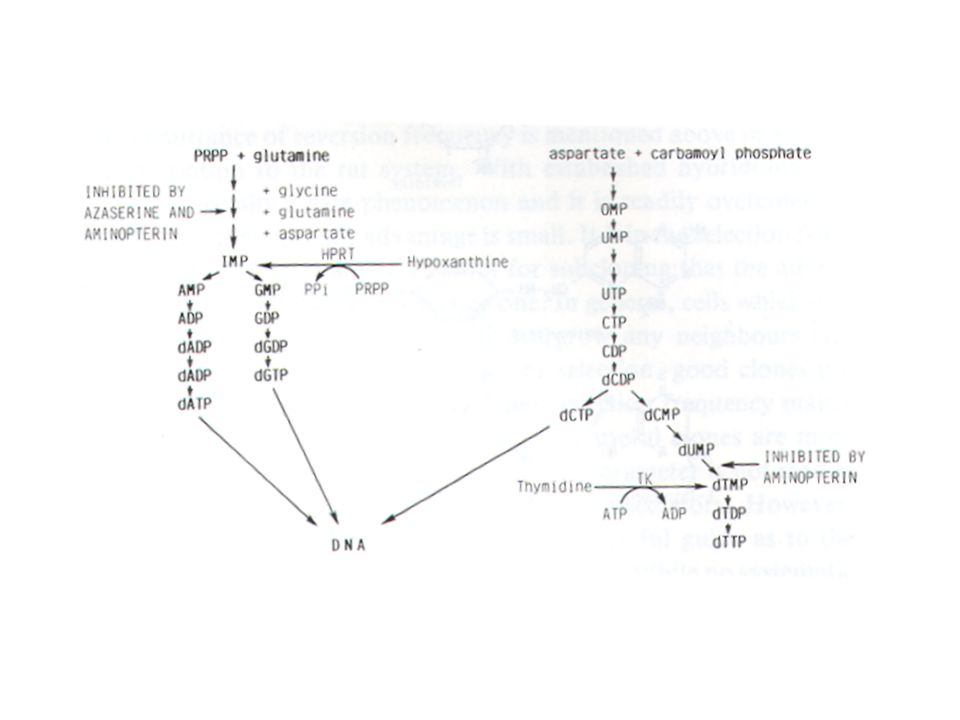
Η σύντηξη των κυττάρων επιτυγχάνεται με την προσθήκη πολυαιθυλενικής γλυκόλης και στους ετεροκαρυώτες οι οποίοι προκύπτουν ο πυρήνας συντήκεται για να δημιουργήσει σταθερά υβριδώματα τα οποία εκφράζουν και τα δύο πατρικά γενώματα ( τα λεμφοκύτταρα ποντικού παράγουν τα αντισώματα και τα μυελωματικά κύτταρα τον θεωρητικά επ’αόριστο πολλαπλασιασμό των κυττάρων). Η κυτταρική σειρά του μυελώματος που θα επιλεγεί δεν θα πρέπει να εκκρίνει αντισώματα ( γιατί τότε το υβρίδωμα θα μας έδινε ένα μίγμα αντισωμάτων) και θα πρέπει να έχει επιλεγεί παρουσία 8-αζαγουανίνης.Τα κύτταρα που θα επιζήσουν θα πρέπει να μην έχουν το ένζυμο της τρανσφεράσης της P-ριβοζυλο-υποξανθίνης. Τα κύτταρα αυτά όταν αναπτύσσονται σε υλικό καλλιέργειας HAT που περιέχει υποξανθίνη , αμινοπτερίνη και θυμιδίνη δεν θα επιβιώσουν διότι δεν μπορούν να συνθέσουν DNA. H αμινοπτερίνη σταματά το κύριο μονοπάτι της βιοσύνθεσης των πουρινών και πυριμιδινών.Τα φυσιολογικά κύτταρα μπορούν να χρησιμοποιήσουν την θυμιδίνη για την σύνθεση πυριμιδινών και την τρανσφεράση της P-ριβοζυλο-υποξανθίνης για να μετατρέψουν την υποξανθίνη σε πουρίνες.Κατά συνέπεια στο υλικό HAT ούτε τα λεμφοκύτταρα ούτε η μυελωματική σειρά μπορεί να αναπτυχθεί. Τα κύτταρα τα οποία θα αναπτυχθούν στο υλικό HAT θα είναι μόνο τα υβριδώματα τα οποία θα έχουν την ικανότητα να πολλαπλασιάζονται επ’αόριστον ( από το μυέλωμα ) και την τρανσφεράση της P-ριβοζυλο-υποξανθίνης και την έκκριση αντισωμάτων από τα λεμφοκύτταρα του ποντικού.
 και θα πρέπει να έχει επιλεγεί παρουσία 8-αζαγουανίνης.Τα κύτταρα που θα επιζήσουν θα πρέπει να μην έχουν το ένζυμο της τρανσφεράσης της P-ριβοζυλο-υποξανθίνης. Τα κύτταρα αυτά όταν αναπτύσσονται σε υλικό καλλιέργειας HAT που περιέχει υποξανθίνη , αμινοπτερίνη και θυμιδίνη δεν θα επιβιώσουν διότι δεν μπορούν να συνθέσουν DNA. H αμινοπτερίνη σταματά το κύριο μονοπάτι της βιοσύνθεσης των πουρινών και πυριμιδινών.Τα φυσιολογικά κύτταρα μπορούν να χρησιμοποιήσουν την θυμιδίνη για την σύνθεση πυριμιδινών και την τρανσφεράση της P-ριβοζυλο-υποξανθίνης για να μετατρέψουν την υποξανθίνη σε πουρίνες.Κατά συνέπεια στο υλικό HAT ούτε τα λεμφοκύτταρα ούτε η μυελωματική σειρά μπορεί να αναπτυχθεί. Τα κύτταρα τα οποία θα αναπτυχθούν στο υλικό HAT θα είναι μόνο τα υβριδώματα τα οποία θα έχουν την ικανότητα να πολλαπλασιάζονται επ’αόριστον ( από το μυέλωμα ) και την τρανσφεράση της P-ριβοζυλο-υποξανθίνης και την έκκριση αντισωμάτων από τα λεμφοκύτταρα του ποντικού.")
6
H αμινοπτερίνη σταματά το κύριο μονοπάτι της βιοσύνθεσης των πουρινών και πυριμιδινών μέσω της αναστολής του ενζύμου διυδροφυλλική αναγωγάση . Τα φυσιολογικά κύτταρα μπορούν να χρησιμοποιήσουν την θυμιδίνη για την σύνθεση πυριμιδινών μέσω του ενζύμου της θυμιδίνης κινάσης και για να μετατρέψουν την υποξανθίνη σε πουρίνες μέσω του ενζύμου τρανσφεράση της P-ριβοζυλο-υποξανθίνης ( Η(G)PRT). Tα κύτταρα μπορούν να καταστούν «ελλειματικά» ως προς αυτά τα ένζυμα μετά από καλλιέρργεια παρουσία της βρωμοδεσοξυουριδίνης ( η οποία επιλέγει κύτταρα ΤΚ- ) ή παρουσία της αζαγουανίνης ή θειογουανίνης ( οι οποίες επιλέγουν κύτταρα HGPRT - ). Σε φυσιολογικά κύτταρα οι αναστολείς αυτοί είναι κυτταροτοξικοί και κατά συνέπεια μόνο κύτταρα τα οποία παρουσιάζουν έλλειμα σχετικά με τα ανωτέρω δυο ένζυμα μπορούν να επιβιώσουν παρουσία αυτών των αναστολέων .
 ή παρουσία της αζαγουανίνης ή θειογουανίνης ( οι οποίες επιλέγουν κύτταρα HGPRT - ). Σε φυσιολογικά κύτταρα οι αναστολείς αυτοί είναι κυτταροτοξικοί και κατά συνέπεια μόνο κύτταρα τα οποία παρουσιάζουν έλλειμα σχετικά με τα ανωτέρω δυο ένζυμα μπορούν να επιβιώσουν παρουσία αυτών των αναστολέων .")
9
Τα κύτταρα αυτά όταν αναπτύσσονται σε υλικό καλλιέργειας HAT που περιέχει υποξανθίνη , αμινοπτερίνη και θυμιδίνη δεν θα επιβιώσουν διότι δεν μπορούν να συνθέσουν DNA. Κατά συνέπεια στο υλικό HAT ούτε τα λεμφοκύτταρα ούτε η μυελωματική σειρά μπορεί να αναπτυχθεί. Τα κύτταρα τα οποία θα αναπτυχθούν στο υλικό HAT θα είναι μόνο τα υβριδώματα τα οποία θα έχουν την ικανότητα να πολλαπλασιάζονται επ’αόριστον ( από το μυέλωμα ) και την τρανσφεράση της P-ριβοζυλο-υποξανθίνης και την έκκριση αντισωμάτων από τα λεμφοκύτταρα του ποντικού.
 και την τρανσφεράση της P-ριβοζυλο-υποξανθίνης και την έκκριση αντισωμάτων από τα λεμφοκύτταρα του ποντικού.")
10
ΠΑΡΑΓΩΓΗ ΜΟΝΟΚΛΩΝΙΚΩΝ ΑΝΤΙΣΩΜΑΤΩΝ ΕΝΑΝΤΙ ΤΟΥ ΑΝΤΙΓΟΝΟΥ Χ

11
Εάν όλα τα υβριδώματα που έχουν αναπτυχθεί μετά την σύντηξη μεγαλώσουν μαζί τότε θα παράγεται ένα μίγμα αντισωμάτων. Συνεπώς πρέπει να απομονωθούν υβριδώματα που θα παράγουν ένα συγκεκριμένο αντίσωμα έναντι ενός συγκεκριμμένου επίτοπου και να αναπτυχθούν ανεξάρτητα. Αυτό επιτυγχάνεται με διαδοχικές αραιώσεις της καλλιέργειας υβριδωμάτων ετσι ώστε να δημιουργηθεί καλλιέργεια η οποία θα περιέχει μόνο ένα κύτταρο.Τα κύτταρα αναπτύσονται τότε σε νέο υλικό ανάπτυξης και κάθε κλώνος εξετάζεται για την ανίχνευση του επιθυμητού μονοκλωνικού αντισώματος.

13
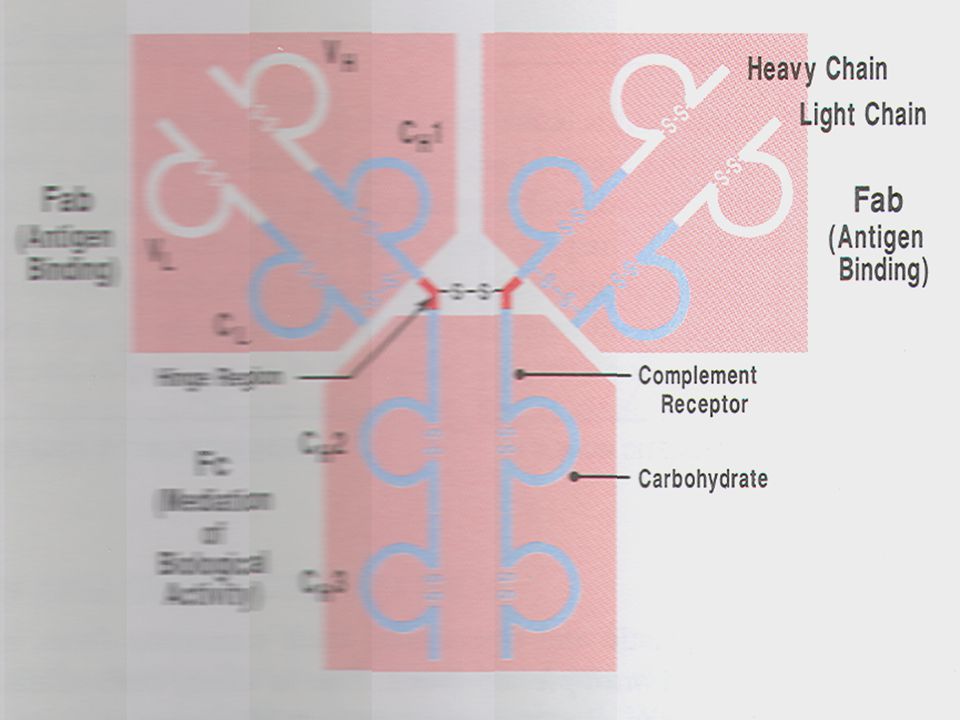
ΤΑ ΘΕΤΙΚΑ ΕΝΑΝΤΙ ΤΟΥ ΑΝΤΙΓΟΝΟΥ ΥΒΡΙΔΩΜΑΤΑ ΑΝΑΠΤΥΣΟΝΤΑΙ ΤΕΛΟΣ ΜΑΖΙΚΑ ΣΕ ΚΥΤΤΑΡΟΚΑΛΛΙΕΡΓΕΙΕΣ Η ΣΕ ΑΣΚΗΤΙΚΟ ΥΓΡΟ Υπάρχουν διάφορες κατηγορίες αντισωμάτων με κυριότερη αυτή των IgG. Tα αντισώματα αυτά αποτελούνται από δυο ίδιες ελαφριές L ( light ) και δυο βαριες αλυσίδες H (heavy). Οι αλυσίδες έχουν εσωτερικούς δισουλφιδικούς δεσμούς και ενώνονται μεταξύ τους επίσης με δισουλφιδικούς δεσμούς δίνοντας στο μόριο το σχήμα Υ. Μελέτη της αμινοξικής αλληλουχίας έδειξε οτι τόσο οι ελαφριές όσο και οι βαριές αλυσίδες αποτελούνται από μια σταθερή περιοχή ( C,constant) και μια μεταβλητή ( V, variable). Μέσα στην περιοχή V κάθε αλυσίδας η οποία ευρίσκεται στα αμινοτελικά άκρα των αλυσίδων υπάρχουν τμήματα που εμφανίζουν μεγαλύτερη ποικιλομορφία και ονομάζονται υπερμεταβλητές περιοχές και εμπλέκονται στην δέσμευση του αντιγόνου.
 και δυο βαριες αλυσίδες H (heavy). Οι αλυσίδες έχουν εσωτερικούς δισουλφιδικούς δεσμούς και ενώνονται μεταξύ τους επίσης με δισουλφιδικούς δεσμούς δίνοντας στο μόριο το σχήμα Υ. Μελέτη της αμινοξικής αλληλουχίας έδειξε οτι τόσο οι ελαφριές όσο και οι βαριές αλυσίδες αποτελούνται από μια σταθερή περιοχή ( C,constant) και μια μεταβλητή ( V, variable). Μέσα στην περιοχή V κάθε αλυσίδας η οποία ευρίσκεται στα αμινοτελικά άκρα των αλυσίδων υπάρχουν τμήματα που εμφανίζουν μεγαλύτερη ποικιλομορφία και ονομάζονται υπερμεταβλητές περιοχές και εμπλέκονται στην δέσμευση του αντιγόνου.")
15
Στο γενετικό υλικό των λεμφοκυττάρων υπάρχουν πολλά διαφορετικά γονίδια για τις μεταβλητές περιοχές των ελαφρών ( VL) και των βαριών αλυσίδων ( VH).Ετσι είναι δυνατόν να προκύψουν πολλοί συνδυασμοί βαριών και ελαφριών αλυσίδων που οδηγούν σε ένα μεγάλο αριθμό αντισωμάτων με διαφορετική εξειδίκευση. Ετσι ενώ ακόμη και αν ενύουμε ένα ποντίκι με το ιδιο αντιγόνο Α σε διαφορετικά χρονικά διαστήματα θα έχομε διεγέρση μεγάλων πλυθυσμών Β λεμφοκυττάρων ( ελαφρώς διαφορετικών μεταξύ τους ) τα οποία θα εκκρίνουν αντισώματα έναντι του αντιγόνου Α , όπου τώρα ο κάθε πλυθυσμός αντισωμάτων θα αναγνωρίζει το αντιγόνο Α αλλά με ένα ελαφρώς διαφορετικό τρόπο ( αναγνωρίζοντας διαφορετικούς επίτοπους πάνω στο ίδιο αντίσωμα).
 τα οποία θα εκκρίνουν αντισώματα έναντι του αντιγόνου Α , όπου τώρα ο κάθε πλυθυσμός αντισωμάτων θα αναγνωρίζει το αντιγόνο Α αλλά με ένα ελαφρώς διαφορετικό τρόπο ( αναγνωρίζοντας διαφορετικούς επίτοπους πάνω στο ίδιο αντίσωμα)..")
17
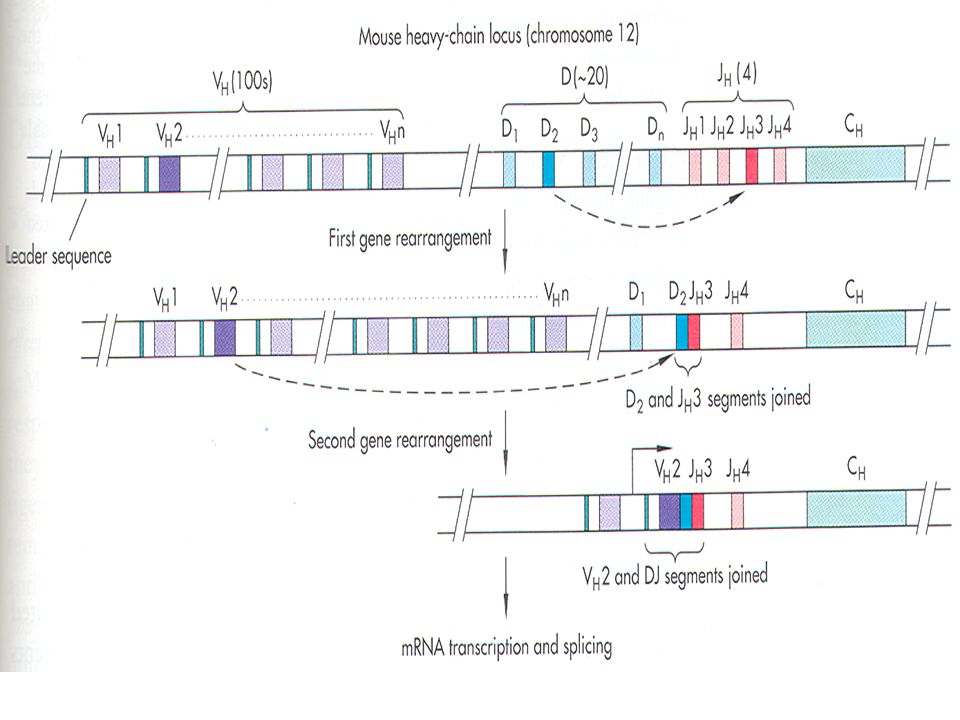
Απεδείχθη τελικά ότι τα VL και VH γονίδια που κωδικοποιούν τις αντίστοιχες περιοχές των αλυσίδων προκύπτουν από τον συνδυασμό περισσότερων γονιδίων . Ετσι τα VL προέρχονται από τα γονίδια V και J ( joining) ώστε το τελικό γονίδιο να είναι VJC. Τα VH σχηματίζονται με την συμμετοχή ακόμα μιας κατηγορίας γονιδίων , τα γονίδια D ( diversity) και ο ανασυνδυασμός γίνεται ώστε να σχηματισθεί αρχικά ένα DJ , κατόπιν ένα VDJ και τέλος VDJC γονίδιο.
 και ο ανασυνδυασμός γίνεται ώστε να σχηματισθεί αρχικά ένα DJ , κατόπιν ένα VDJ και τέλος VDJC γονίδιο.")
18
Ανασυνδυασμός των ελαφριών αλυσίδων κ
Η ελαφριά αλυσίδα κ του ποντικού περιέχει περίπου : 300 μεταβλητές αλληλουχίες (segments V), 4 J και μια C . Οταν ανασυνδυάζονται π.χ. η αλληλουχία Vκ3 ενώνεται με την αλληλουχία Jκ4 και αποτελούν μια μεταγραφική μονάδα η οποία μετά το μάτισμα δίνει ένα mRNA ( Vκ3Jκ4Cκ ) το οποίο κωδικοποιεί για την ώριμη πλέον κ αλυσίδα. Η αλληλουχία L κωδικοπoιεί για ένα πεπτίδιο οδηγό το οποίο ευρίσκεται στο αμινο- τελικό άκρο της κάθε αλυσίδας .Το πεπτίδιο L κόβεται κατά την διάρκεια της έκκρισης
, 4 J και μια C . Οταν ανασυνδυάζονται π.χ. η αλληλουχία Vκ3 ενώνεται με την αλληλουχία Jκ4 και αποτελούν μια μεταγραφική μονάδα η οποία μετά το μάτισμα δίνει ένα mRNA ( Vκ3Jκ4Cκ ) το οποίο κωδικοποιεί για την ώριμη πλέον κ αλυσίδα. Η αλληλουχία L κωδικοπoιεί για ένα πεπτίδιο οδηγό το οποίο ευρίσκεται στο αμινο- τελικό άκρο της κάθε αλυσίδας .Το πεπτίδιο L κόβεται κατά την διάρκεια της έκκρισης.")
19
Ανασυνδυασμός των βαριών αλυσίδων του ποντικού.
Εκτός των 100 περίπου μεταβλητών αλληλουχιών ( segments – 100 V ) , των 4J και της μια C οι βαριές αλυσίδες περιέχουν επίσης 20 μεταβλητές αλληλουχίες ( D segments ) οι οποίες προσθέτουν επι πλέον ποικιλομορφία . Ο ανασυνδυασμός των βαριών αλυσίδων γίνεται σε δυο στάδια: ανασυνδυασμός μεταξύ μιας μεταβλητής αλληλουχίας D και μιας J , π.χ. D2 και JH3 και ανασυνδυασμός μεταξύ της ήδη ανασυνδυασμένης αλληλουχίας π.χ. D2 και JH3 και μιας μεταβλητής V αλληλουχίας π.χ. VH2 και D2 -JH3. Εν συνεχεία το ανασυνδυασμένο γονίδιο μεταγράφεται και μετά το μάτισμα δίνει το mRNA το οποίο κωδικοποιεί πλέον για την ώριμη βαριά αλυσίδα VH2 - D2 -JH3 - CH.
 , των 4J και της μια C οι βαριές αλυσίδες περιέχουν επίσης 20 μεταβλητές αλληλουχίες ( D segments ) οι οποίες προσθέτουν επι πλέον ποικιλομορφία . Ο ανασυνδυασμός των βαριών αλυσίδων γίνεται σε δυο στάδια: ανασυνδυασμός μεταξύ μιας μεταβλητής αλληλουχίας D και μιας J , π.χ. D2 και JH3 και. ανασυνδυασμός μεταξύ της ήδη ανασυνδυασμένης αλληλουχίας π.χ. D2 και JH3 και μιας μεταβλητής V αλληλουχίας π.χ. VH2 και. D2 -JH3. Εν συνεχεία το ανασυνδυασμένο γονίδιο μεταγράφεται και μετά το μάτισμα δίνει το mRNA το οποίο κωδικοποιεί πλέον για την ώριμη βαριά αλυσίδα VH2 - D2 -JH3 - CH.")
21
Μεταγραφική Ρύθμιση Καθεμία των 300 μεταβλητών αλληλουχιών (segments V) στο αλλήλιο κ εχει τον δικό της επαγωγέα( P ).Oταν μια μεταβλητή αλληλουχία V ανασυνδυάζεται με μια αλληλουχία J τότε ο επαγωγέας τίθεται υπό τον έλεγχο ενός μεταγραφικού ενισχυτή ( Ε ) ο οποίος ευρίσκεται στην περιοχή των εσονίων μεταξύ J και C.
 στο αλλήλιο κ εχει τον δικό της επαγωγέα( P ).Oταν μια μεταβλητή αλληλουχία V ανασυνδυάζεται με μια αλληλουχία J τότε ο επαγωγέας τίθεται υπό τον έλεγχο ενός μεταγραφικού ενισχυτή ( Ε ) ο οποίος ευρίσκεται στην περιοχή των εσονίων μεταξύ J και C.")
22
Αλλαγή της τάξης των βαριών αλυσίδων
Αρχικά η ανασυνδυασμένη βαριά αλυσίδα V-D-J είναι της τάξης των IgM ( V-D-J-Cμ ). Κατά την αλλαγή της τάξης των ανοσοσφαιρινών η ανασυνδυασμένη μονάδα V-D-J ενώνεται με ένα νέο γονίδιο βαριών αλυσίδων π.χ. το Cγ2b, κωδικοποιώντας με αυτό τον τρόπο για μια νέα ανοσοσφαιρίνη την IgG2b.Ετσι ο ίδιος ανασυνδυασμός V-D-J μπορεί να χρησιμοποιηθεί για την παραγωγή διαφορετικών τάξεων ανοσοσφαιρινών.
. Κατά την αλλαγή της τάξης των ανοσοσφαιρινών η ανασυνδυασμένη μονάδα V-D-J ενώνεται με ένα νέο γονίδιο βαριών αλυσίδων π.χ. το Cγ2b, κωδικοποιώντας με αυτό τον τρόπο για μια νέα ανοσοσφαιρίνη την IgG2b.Ετσι ο ίδιος ανασυνδυασμός V-D-J μπορεί να χρησιμοποιηθεί για την παραγωγή διαφορετικών τάξεων ανοσοσφαιρινών.")
23
ΤΑΞΕΙΣ ΚΑΙ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΩΝ ΑΝΟΣΟΣΦΑΙΡΙΝΩΝ
ΤΑΞΕΙΣ ΚΑΙ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΤΩΝ ΑΝΟΣΟΣΦΑΙΡΙΝΩΝ

24
ΜΕΘΟΔΟΙ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΤΗΣ ΑΝΤΙΔΡΑΣΗΣ ΑΝΤΙΓΟΝΟΥ-ΑΝΤΙΣΩΜΑΤΟΣ
ΜΕΘΟΔΟΙ ΓΙΑ ΤΗΝ ΑΝΙΧΝΕΥΣΗ ΤΗΣ ΑΝΤΙΔΡΑΣΗΣ ΑΝΤΙΓΟΝΟΥ-ΑΝΤΙΣΩΜΑΤΟΣ Η Συγκόλληση (Αgglutination) στηρίζεται στη δημιουργία ορατών συσσωματωμάτων εξαιτίας της σύνδεσης των ειδικών αντισωμάτων με αντιγόνα της επιφάνειας κυττάρων ή με αντιγόνα που είναι ακινητοποιημένα σε τεχνητά υποστρώματα. Εχει εφαρμοσθεί για την αναγνώριση τόσο των κατά Gram αρνητικών βακτηρίων (Salmonella, Escherichia coli), όσο και των κατά Gram θετικών (Streptococcus pneumoniae). Μια τροποποίηση αυτής της τεχνικής είναι η λεγόμενη συνσυγκόλληση (Co-agglutination) στην οποία το αντίσωμα, πριν την χρησιμοποίησή του για αναγνώριση, αναμειγνύεται με κύτταρα του Staphylococcus aureus όπου η περιοχή Fc των IgG αντισωμάτων συνδέεται μη ειδικά με την πρωτεϊνη Α του S. aureus, ενώ οι περιοχές Fab του αντισώματος παραμένουν ελεύθερες για την σύνδεσή τους με τα συγκεκριμένα αντιγόνα.
 στηρίζεται στη δημιουργία ορατών συσσωματωμάτων εξαιτίας της σύνδεσης των ειδικών αντισωμάτων με αντιγόνα της επιφάνειας κυττάρων ή με αντιγόνα που είναι ακινητοποιημένα σε τεχνητά υποστρώματα. Εχει εφαρμοσθεί για την αναγνώριση τόσο των κατά Gram αρνητικών βακτηρίων (Salmonella, Escherichia coli), όσο και των κατά Gram θετικών (Streptococcus pneumoniae). Μια τροποποίηση αυτής της τεχνικής είναι η λεγόμενη συνσυγκόλληση (Co-agglutination) στην οποία το αντίσωμα, πριν την χρησιμοποίησή του για αναγνώριση, αναμειγνύεται με κύτταρα του Staphylococcus aureus όπου η περιοχή Fc των IgG αντισωμάτων συνδέεται μη ειδικά με την πρωτεϊνη Α του S. aureus, ενώ οι περιοχές Fab του αντισώματος παραμένουν ελεύθερες για την σύνδεσή τους με τα συγκεκριμένα αντιγόνα.")
25
Οι σταφυλόκοκκοι,πριν την ανάμειξη τους με το αντίσωμα, μονιμοποιούνται με φορμαλδεϊδη (0,5%) στους 80ºC για λίγα λεπτά. Η διαδικασία αυτή έχει ως απότέλεσμα τη διατήρηση της ικανότητας σύνδεσης τους με το αντίσωμα για αρκετούς μήνες εφ’ όσον αυτοί διατηρούνται στο ψυγείο. Για την εκτέλεση του ελέγχου της συν-συγκόλλησης αναμειγνύονται σε μια αντικειμενοφόρο πλάκα μια σταγόνα του αιωρήματος του σταφυλόκοκκου (S. aureus σε φυσιολογικό ορό) με μια σταγόνα του αντι-ορού και εν συνεχεία προσθέτονται τα βακτήρια τα οποία αναγνωρίζονται από το αντίσωμα με αποτέλεσμα την εμφάνιση συγκόλλησης ( agglutination). Σε όλες τις αντιδράσεις αντιγόνου-αντισώματος μεγάλη σημασία έχει η τιτλοποίηση του αντισώματος σε σχέση με την συγκέντρωση αντιγόνου. Για αυτό το λόγο συνίσταται η κάθε αντίδραση συγκόλλησης να πραγματοποιείται σε 2-3 διαφορετικές αραιώσεις του αντισώματος.

26
Στο ανωτέρω παράδειγμα οι αραιώσεις αντισώματος (1/4,1/8 και 1/2048 ) δεν δίνουν συγκόλληση , ενώ συγκόλληση παρατηρείται από τις αραιώσεις αντισώματος 1/16 μέχρι και 1/1024. Η τελευταία αυτή αραίωση που δίνει συγκόλληση αποτελεί και τον τίτλο του συγκεκριμένου αντισώματος έναντι της συγκεκριμένης συγκέντρωσης αντιγόνου. Κατά συνέπεια η σχέση αντιγόνου –αντισώματος είναι μια δυναμική σχέση και στη συγκόλληση απαιτείται ισορροπία μεταξύ των δυο αντιδρώντων .Για μια σταθερή συγκέντρωση αντιγόνου η πολύ υψηλή συγκέντρωση αντισώματος ( φαινόμενο προζώνης ) δεν επιτρέπει την συγκόλληση γιατί κάθε επίτοπος έχει την «πολυτέλεια» να δεσμεύει από μόνος του ένα αντίσωμα αποτρέποντας ετσι τον σχηματισμό «γέφυρας» μεταξύ διαφορετικών επιτόπων και αντισώματος .Τέλος όταν η αραίωση του αντισώματος είναι πολύ μεγάλη τότε οι πιθανότητες σχηματισμού συγκόλλησης ελαχιστοποιούνται ( αραίωση 1/2048 και μεγαλύτερες).
 δεν επιτρέπει την συγκόλληση γιατί κάθε επίτοπος έχει την «πολυτέλεια» να δεσμεύει από μόνος του ένα αντίσωμα αποτρέποντας ετσι τον σχηματισμό «γέφυρας» μεταξύ διαφορετικών επιτόπων και αντισώματος .Τέλος όταν η αραίωση του αντισώματος είναι πολύ μεγάλη τότε οι πιθανότητες σχηματισμού συγκόλλησης ελαχιστοποιούνται ( αραίωση 1/2048 και μεγαλύτερες).")
27
Σε περίσεια αντισώματος δεν παρατηρείται συγκόλληση
Σε συνθήκες ισορροπίας αντιγόνου–αντισώματος παρατηρείται συγκόλληση

28
Αιμοσυγκόλληση Για ιούς, όπως οι ιοί της γρίππης, οι οποίοι , λόγω της αιμοσυγκολλητίνης που περιέχεται στο φάκελλό τους προκαλούν συγκόλληση ερυθροκυττάρων ( κοτόπουλου ή χήνας), η τιτλοποίηση γίνεται με επώαση του ιού με ερυθροκύτταρα και με μακροσκοπική παρατήρηση τους για τον προσδιορισμό της μεγαλύτερης αραίωσης του ιού που προκαλεί συγκόλληση των ερυθροκυττάρων ( αιμοσυγκόλληση). Η συσχέτιση μονάδων αιμοσυγκόλλησης προς τον αριθμό ιοσωματίων γίνεται από τον αριθμό ερυθροκυττάρων στην κάθε οπή και από την μεγαλύτερη αραίωση του ιού που προκαλεί ολική συγκόλληση των ερυθροκυττάρων . Στο κατωτέρω παράδειγμα ο ιός τιτλοποιείται εις διπλούν με υποδιπλάσιες αραιώσεις ( αρχίζοντας από την θέση 1Α και 1Β αραίωση 1/4 και εν συνεχεία 1/8 , 1/16 , 1/32 , 1/64 , 1/128 , 1/256 κ.ο.κ.) . Ως τίτλος του ιού δίνεται η τελευταία αραίωση η οποία δίνει μια πλήρη αιμοσυγκόλληση δηλ. η αραίωση 1/32 στη θέση 4Α και 4Β.Οταν δεν παρατηρείται αιμοσυγκόλληση τα ερυθρά καθιζάνουν ( θέσεις 5,6,7 Α και Β).
, η τιτλοποίηση γίνεται με επώαση του ιού με ερυθροκύτταρα και με μακροσκοπική παρατήρηση τους για τον προσδιορισμό της μεγαλύτερης αραίωσης του ιού που προκαλεί συγκόλληση των ερυθροκυττάρων ( αιμοσυγκόλληση). Η συσχέτιση μονάδων αιμοσυγκόλλησης προς τον αριθμό ιοσωματίων γίνεται από τον αριθμό ερυθροκυττάρων στην κάθε οπή και από την μεγαλύτερη αραίωση του ιού που προκαλεί ολική συγκόλληση των ερυθροκυττάρων . Στο κατωτέρω παράδειγμα ο ιός τιτλοποιείται εις διπλούν με υποδιπλάσιες αραιώσεις ( αρχίζοντας από την θέση 1Α και 1Β αραίωση 1/4 και εν συνεχεία 1/8 , 1/16 , 1/32 , 1/64 , 1/128 , 1/256 κ.ο.κ.) . Ως τίτλος του ιού δίνεται η τελευταία αραίωση η οποία δίνει μια πλήρη αιμοσυγκόλληση δηλ. η αραίωση 1/32 στη θέση 4Α και 4Β.Οταν δεν παρατηρείται αιμοσυγκόλληση τα ερυθρά καθιζάνουν ( θέσεις 5,6,7 Α και Β).")
29
Μια παραλλαγή της ανωτέρω τεχνικής είναι η αναστολή της αιμοσυγκόλλησης.Στην τεχνική αυτή επωάζονται πρώτα π.χ. το αντίσωμα έναντι του ιού της Γρίππης Α με τον ιό και εν συνεχεία προσθέτουμε τα ερυθρά ( χήνας ή κοτόπουλου) τα οποία προκαλούν αιμοσυγκόλληση.Στην περίπτωση αυτή η ποσότητα του αντισώματος που επωάσαμε δεσμεύει την αιμοσυγκολλητίνη του ιού και ετσι έχουμε αναστολή της αιμοσυγκόλλησης.Πρακτικά η μέθοδος αυτή χρησιμοποιείται για την ανίχνευση στον ορό ασθενών αντισωμάτων έναντι του ιού της Γρίππης Α. Συνήθως χρησιμοποιούμε 4 αιμοσυγκολλητικές μονάδες του ιού τις οποίες έχουμε προσδιορίσει προηγουμένως με τον τρόπο που αναφέρεται ανωτέρω. Ως αιμοσυγκολλητική μονάδα ορίζεται η υψηλότερη αραίωση του ιού η οποία δίνει αιμοσυγκόλληση ( στο ανωτέρω παράδειγμα 1/32 ) οπότε οι 4 αιμοσυγκολλητικές μονάδες αντιστοιχούν στην αραίωση 1/8 του ιού. Πραγματοποιούμε διαδοχικές αραιώσεις του ορού του ασθενούς συνήθως από 1/8 μέχρι 1/1024 σε ρυθμιστικό διάλυμα και απ’ευθείας σε μικροπλάκες τιτλοποιήσεως των 96 δεξαμενών σχήματος V ή U . Προσθέτουμε εν συνεχεία 4 αιμοσυγκολλητικές μονάδες του ιού σε όλες τις δεξαμενές εκτός από την δεξαμενή στη θέση 1 η οποία θα αποτελέσει τον μάρτυρα ορού ( serum control ) και στην θέση 12 η οποία θα αποτελέσει τον μάρτυρα ερυθροκυττάρων. Οι οροί είναι πιθανόν να περιέχουν αναστολείς της αιμοσυγκόλλησης και γι’αυτό το λόγο τους επωάζουμε συνήθως στους 56 0C επί 30 λεπτά. Όπως επίσης επωάζουμε τον ορό με τα ερυθροκύτταρα πριν το τεστ έτσι ώστε ο ορός να απαλλαγεί από τις τυχόν μη ειδικές αιμοσυγκολλητίνες τις οποίες ενδεχομένως να περιέχει.
 και στην θέση 12 η οποία θα αποτελέσει τον μάρτυρα ερυθροκυττάρων. Οι οροί είναι πιθανόν να περιέχουν αναστολείς της αιμοσυγκόλλησης και γι’αυτό το λόγο τους επωάζουμε συνήθως στους 56 0C επί 30 λεπτά. Όπως επίσης επωάζουμε τον ορό με τα ερυθροκύτταρα πριν το τεστ έτσι ώστε ο ορός να απαλλαγεί από τις τυχόν μη ειδικές αιμοσυγκολλητίνες τις οποίες ενδεχομένως να περιέχει.")
30
Η μικροπλάκα αφήνεται στη συνέχεια επί 60 λεπτά σε θερμοκρασία δωματίου και εν συνεχεία προσθέτουμε 0,5% ερυθρά και επωάζουμε την μικροπλάκα επι 60 λεπτά στους +4 0C. Η μικροπλάκα είναι πλέον έτοιμη για ανάγνωση. Στις θέσεις 1 και 12 της μικροπλάκας τα ερυθροκύτταρα θα καθιζάνουν στο βάθος της δεξαμενής.Στις θέσεις όπου τα αντισώματα θα έχουν εξουδετερώσει τον ιό θα έχουμε αναστολή της αιμοσυγκόλλησης .Ο τίτλος του αντισώματος κάθε ασθενούς εκφράζεται με την υψηλότερη αραίωση του ορού η οποία δίνει αναστολή της αιμοσυγκόλλησης . Στις θέσεις όπου θα παρατηρείται αιμοσυγκόλληση τα αντισώματα δεν θα είναι σε συγκέντρωση ικανή να δεσμεύσουν τον ιό.

31
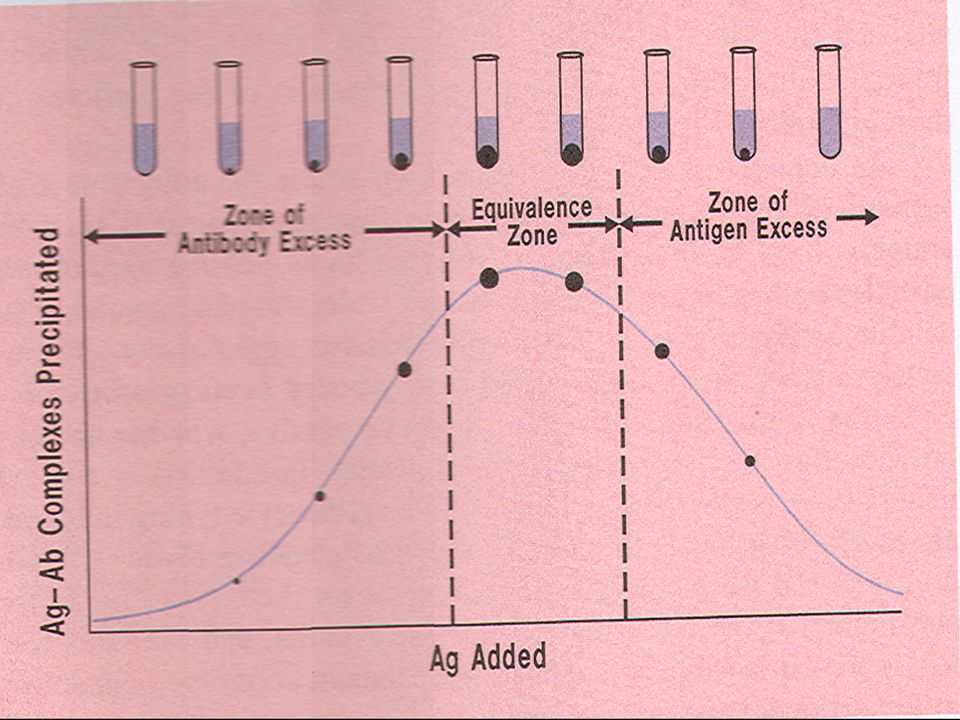
Ανοσοδιάχυση Η αρχή της μεθόδου στηρίζεται στο γεγονός ότι, όταν ένα αντίσωμα συνδεθεί με το αντίστοιχό του αντιγόνο, σχηματίζεται ίζημα η ποσότητα του οποίου εξαρτάται από το μέγεθος των συμπλόκων που δημιουργούνται και από τις σχετικές συγκεντρώσεις αντισώματος/αντιγόνου. Υπάρχει πάντα η ιδανική αναλογία στις συγκεντρώσεις αντισώματος/αντιγόνου όπου σχηματίζονται μεγάλα συσσωματώματα που εύκολα δίνουν ίζημα. Για μια δεδομένη συγκέντρωση αντισώματος σε αυξανόμενες συγκεντρώσεις αντιγόνου όταν η αντίδραση αυτή πραγματοποιείται σε διάλυμα έχουμε την καμπύλη όπως παρουσιάζεται κατωτέρω. Ετσι στη ζώνη ισοδυναμίας και στη γειτνίαση αυτής θα έχουμε τα πιο έντονα ιζήματα. Πρίν τη ζώνη ισοδυναμίας , την ζώνη όπου το αντίσωμα είναι σε πλεόνασμα θα έχουμε την παρουσία μικρού έως ελάχιστου ιζήματος όπως και στη ζώνη όπου πλεονάζει το αντιγόνο. Και στις δύο αυτές περιπτώσεις υπάρχουν στο υπερκείμενο διαλυτά σύμπλοκα αντιγόνου-αντισώματος τα οποία δεν δίνουν ίζημα εκτός της ζώνης ισοδυναμίας.

33
Αυτές οι αντιδράσεις μπορούν επίσης να γίνουν και σε πήκτωμα αγαρόζης όπου το διάλυμα του αντισώματος και του αντιγόνου τοποθετούνται σε δεξαμενές που έχουν δημιουργηθεί μέσα στην αγαρόζη.Τα δυο αντιδρώντα συστατικά διαχέονται μέσα στην αγαρόζη σχηματίζοντας μια βαθμίδωση συγκέντρωσης και στο σημείο όπου η αναλογία αντιγόνου /αντισώματος είναι μέγιστη θα έχουμε σχηματισμό ιζήματος. Αν οι δεξαμενές οι οποίες περιέχουν το αντίσωμα (Ab) και το αντιγόνο ( Ag) περιέχουν τα αντισώματα 1,2,3 ειδικά των αντιγόνων 1,2 και 3 αντίστοιχα τα οποία διαχέονται με διαφορετική ταχύτητα ( το αντιγόνο 1 με μεγαλύτερη από το 2 και το 2 με μεγαλύτερη από το 3 ) , τότε θα έχουμε τον σχηματισμό 3 διαφορετικών ιζημάτων στην ζώνη ισοδυναμίας.
 και το αντιγόνο ( Ag) περιέχουν τα αντισώματα 1,2,3 ειδικά των αντιγόνων 1,2 και 3 αντίστοιχα τα οποία διαχέονται με διαφορετική ταχύτητα ( το αντιγόνο 1 με μεγαλύτερη από το 2 και το 2 με μεγαλύτερη από το 3 ) , τότε θα έχουμε τον σχηματισμό 3 διαφορετικών ιζημάτων στην ζώνη ισοδυναμίας.")
34
Με την τεχνική αυτή είναι δυνατόν να ελέγξουμε την αντιγονική σχέση 2 αντιγόνων ( Ag) με την βοήθεια 2 αντισωμάτων (Ab). Αριστερά: τα αντιγόνα 1 αναγνωρίζονται από το ίδιο αντίσωμα και κατά συνέπεια έχουν κοινούς αντιγονικούς καθοριστές. Στο κέντρο: τα αντιγόνα 1 και 2 αναγνωρίζονται από 2 διαφορετικά αντισώματα και δεν παρουσιάζουν κοινούς αντιγονικούς καθοριστές. Δεξιά: τα αντιγόνα 1 και 2 αναγνωρίζονται από το ίδιο αντίσωμα παρουσιάζοντας κοινούς αλλά και μη κοινούς αντιγονικούς καθοριστές.Το αντίσωμα 1 αναγνωρίζει περισσότερους αντιγονικούς καθοριστές στο αντιγόνο 1 από ότι στο αντιγόνο 2 ( η γραμμή εκτός της συνεχόμενης κυρτής γραμμής).
.")
35
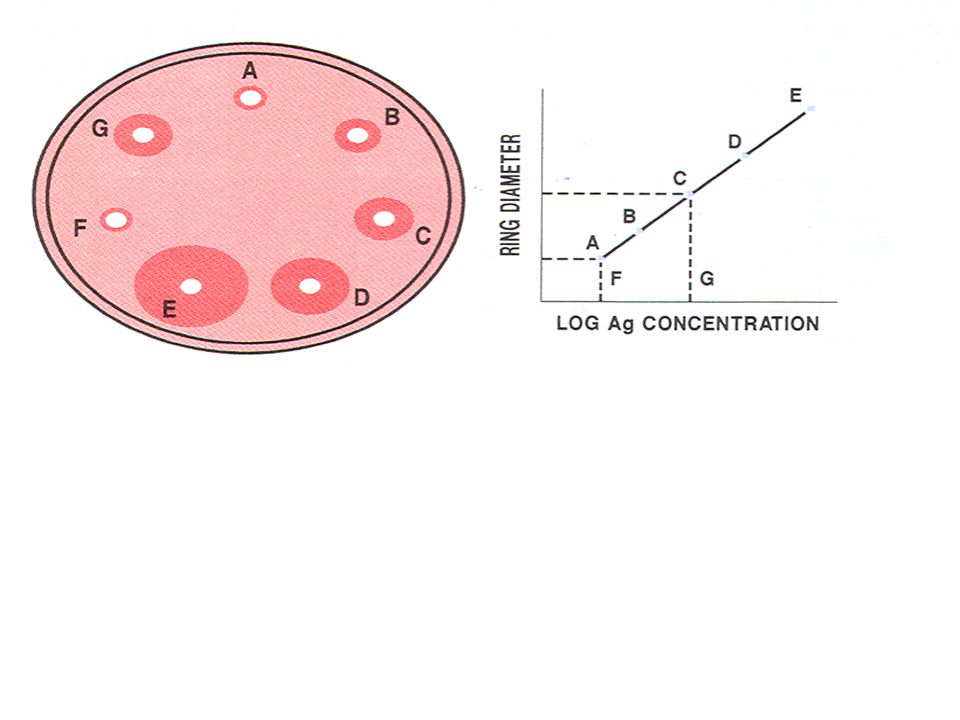
Αν στο κέντρο ενός τρυβλίου τοποθετήσουμε το αντίσωμα και περιφερειακά 6 αντιγόνα είναι δυνατόν να μελετήσουμε την αντιγονική συσχέτιση μεταξύ των αντιγόνων αυτών. Aν σε όλο το πήκτωμα αγαρόζης διαχυθεί αντίσωμα και σε 5 τουλάχιστον δεξαμενές τοποθετηθούν αυξανόμενες γνωστές συγκεντρώσεις αντιγόνου ( Α, Β, C, D, E ) είναι δυνατόν να προσδιορίσουμε την συγκέντρωση των 2 αγνώστων δειγμάτων ( F ,G ) βάσει της καμπύλης η οποία παρουσιάζεται κατωτέρω.Στην τεχνική αυτή η διάχυση των αντιγόνων γίνεται περιμετρικά των δεξαμενών και το ίζημα το οποίο σχηματίζεται εχει την μορφή δακτυλίου.Μετρώντας την διάμετρο κάθε δακτυλίου για τις γνωστές συγκεντρώσεις αντιγόνου και εκφράζοντας τις συγκεντρώσεις αυτές σε λογαριθμική μορφή παίρνουμε την κατωτέρω πρότυπη καμπύλη πάνω στην οποία αναφερόμαστε για να προσδιορίσουμε τα άγνωστα δείγματα.
 είναι δυνατόν να προσδιορίσουμε την συγκέντρωση των 2 αγνώστων δειγμάτων ( F ,G ) βάσει της καμπύλης η οποία παρουσιάζεται κατωτέρω.Στην τεχνική αυτή η διάχυση των αντιγόνων γίνεται περιμετρικά των δεξαμενών και το ίζημα το οποίο σχηματίζεται εχει την μορφή δακτυλίου.Μετρώντας την διάμετρο κάθε δακτυλίου για τις γνωστές συγκεντρώσεις αντιγόνου και εκφράζοντας τις συγκεντρώσεις αυτές σε λογαριθμική μορφή παίρνουμε την κατωτέρω πρότυπη καμπύλη πάνω στην οποία αναφερόμαστε για να προσδιορίσουμε τα άγνωστα δείγματα.")
37
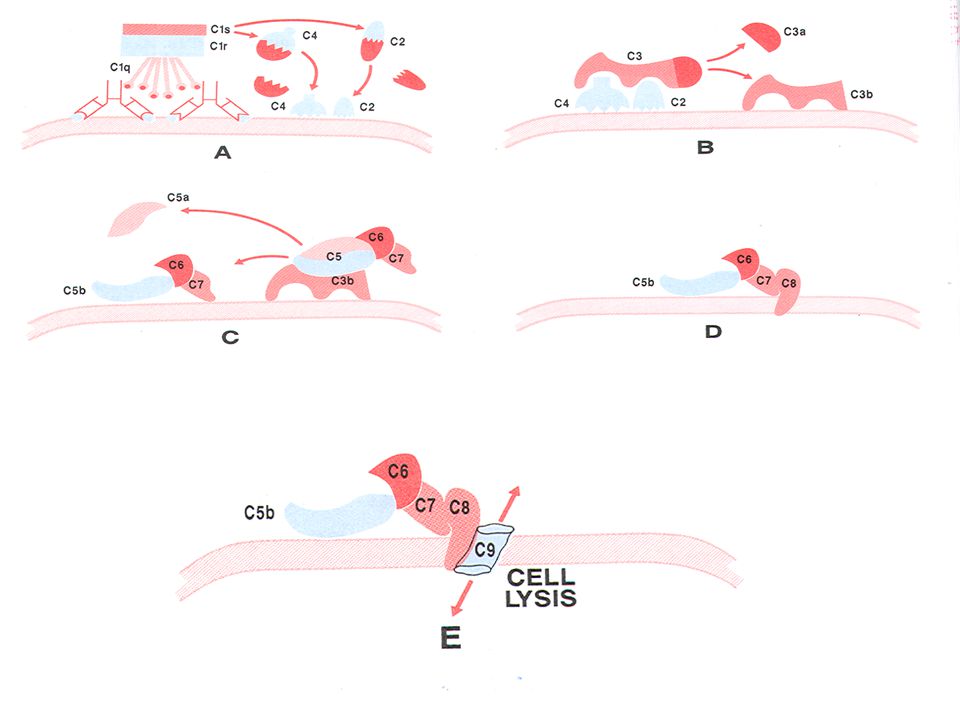
Σύνδεση συμπληρώματος
Σύνδεση συμπληρώματος Δεδομένου ότι το σύστημα του συμπληρώματος ενεργοποιείται μόνο από σύμπλοκα αντισώματος/αντιγόνου είναι δυνατόν η ιδιότητα αυτή να χρησιμοποιηθεί για την ανίχνευση ειδικών αντισωμάτων ή αντιγόνων. Δεδομένου ότι η σύνδεση συστατικών του συμπληρώματος με σύμπλοκα αντισώματος/αντιγόνου δεν δημιουργεί εμφανή αποτελέσματα χρησιμοποιείται στον έλεγχο αυτόν, ένα σύστημα-δείκτης τα ερυθροκύτταρα προβάτου. Το συμπλήρωμα από ορό ινδικών χοιριδίων αποτελεί τη συνηθέστερη πηγή συμπληρώματος. Ο έλεγχος του συμπληρώματος περιλαμβάνει δύο στάδια αφού χρειάζεται πρώτα να αντιδράσει το αντιγόνο με το αντίσωμα και στη συνέχεια με το συμπλήρωμα· το μείγμα αφήνεται ακολούθως να αντιδράσει με τα ερυθροκύτταρα προβάτου καλυμμένα με αντίσωμα έναντι των ερυθρών του προβάτου. Η τεχνική αυτή πραγματοποιείται επίσης σε μικροπλάκα και προϋποθέτει εκτός από την τιτλοποίηση αντισώματος στην οποία αναφερθήκαμε ανωτέρω και την ιδανική συγκέντρωση του αντιγόνου ( ώστε να εργαζόμαστε σε συνθήκες πολύ κοντά στην ισορροπία του συστήματος) και τους εξής 3 ελέγχους : 1. Μία δεξαμενή όπου θα υπάρχει μόνο το αντιγόνο και το συμπλήρωμα για την ανίχνευση μη ειδικών αντιδράσεων. Στην περίπτωση αυτή τα ερυθρά θα πρέπει να καθιζάνουν στο βάθος της δεξαμενής. 2.Τιτλοποίση του συμπληρώματος παρουσία σταθερής συγκέντρωσης αντισώματος αντι ερυθρών προβάτου και % ερυθρών προβάτου (π.χ. 0,5%). 3.Λύση των ερυθρών. Εχοντας προηγουμένως τιτλοποιήσει το συμπλήρωμα θα χρησιμοποιηθεί στον συγκεκριμένο αυτό τίτλο για την επιτυχή λύση ( 100%) των ερυθρών επικαλυμμένων με το αντίσωμα αντι-ερυθρά προβάτου.
 και τους εξής 3 ελέγχους : 1. Μία δεξαμενή όπου θα υπάρχει μόνο το αντιγόνο και το συμπλήρωμα για την ανίχνευση μη ειδικών αντιδράσεων. Στην περίπτωση αυτή τα ερυθρά θα πρέπει να καθιζάνουν στο βάθος της δεξαμενής. 2.Τιτλοποίση του συμπληρώματος παρουσία σταθερής συγκέντρωσης αντισώματος αντι ερυθρών προβάτου και % ερυθρών προβάτου (π.χ. 0,5%). 3.Λύση των ερυθρών. Εχοντας προηγουμένως τιτλοποιήσει το συμπλήρωμα θα χρησιμοποιηθεί στον συγκεκριμένο αυτό τίτλο για την επιτυχή λύση. ( 100%) των ερυθρών επικαλυμμένων με το αντίσωμα αντι-ερυθρά προβάτου.")
39
Η ενεργοποίηση του συμπληρώματος πραγματοποιείται υπό μορφή καταρράκτη μέσω των 9 κύριων συστατικών του ( C1- C9 ). To C1q είναι ένα πολυμερές 6 όμοιων υπομονάδων και για την ενεργοποίηση του απαιτεί την σύνδεσή του με το Fc τουλάχιστιν δυο παραπλήσιων IgG ή μιας IgM . Κατά συνέπεια το συμπλήρωμα ενεργοποιείται πρωτίστως από τις ανοσοσφαιρίνες IgM και κατά δεύτερο λόγο από τις IgG. Τέλος τα συστατικά του συμπληρώματος C8 και C9 εισέρχονται μέσα στη μεμβράνη του κυττάρου δημιουργώντας διόδους ιόντων διαταράσσοντας την οσμωτική ισορροπία με αποτέλεσμα την είσοδο νερού στο κύτταρο που οδηγεί στη λύση του κυττάρου. Συνήθως πραγματοποιούμε υποδιπλάσιες αραιώσεις του ορού ασθενούς σε μικροπλάκα τιτλοποίησης ( αραιώσεις 1/8 μέχρι 1/1023 ) .Προσθέτουμε το αντιγόνο στη συγκεκριμένη συγκέντρωση , το συμπλήρωμα στον συγκεκριμένο τίτλο και επωάζουμε επί 60 λεπτά. Προσθέτουμε τέλος τα ερυθρά προβάτου και το αντίσωμα αντι ερυθρά προβάτου και επωάζουμε επί λεπτά . Η μικροπλάκα είναι έτοιμη για ανάγνωση. Στις δεξαμενές όπου το αντίσωμα ( ορός του ασθενούς) αναγνωρίζει το αντιγόνο το συμπλήρωμα καταναλώνεται στην αντίδραση και δεν μπορεί να λύσει τα ερυθρά .Στην περίπτωση αυτή τα ερυθρά καθιζάνουν. Αν δεν υπάρχει επαρκής ή καθόλου ποσότητα αντισώματος έναντι του αντιγόνου τότε το συμπλήρωμα δεν καταναλώνεται στην 1η αντίδραση αλλά στην αντίδραση ερυθρά προβάτου - αντίσωμα αντιερυθρών προβάτου και προκαλεί λύση των ερυθρών.
 .Προσθέτουμε το αντιγόνο στη συγκεκριμένη συγκέντρωση , το συμπλήρωμα στον συγκεκριμένο τίτλο και επωάζουμε επί 60 λεπτά. Προσθέτουμε τέλος τα ερυθρά προβάτου και το αντίσωμα αντι ερυθρά προβάτου και επωάζουμε επί λεπτά . Η μικροπλάκα είναι έτοιμη για ανάγνωση. Στις δεξαμενές όπου το αντίσωμα ( ορός του ασθενούς) αναγνωρίζει το αντιγόνο το συμπλήρωμα καταναλώνεται στην αντίδραση και δεν μπορεί να λύσει τα ερυθρά .Στην περίπτωση αυτή τα ερυθρά καθιζάνουν. Αν δεν υπάρχει επαρκής ή καθόλου ποσότητα αντισώματος έναντι του αντιγόνου τότε το συμπλήρωμα δεν καταναλώνεται στην 1η αντίδραση αλλά στην αντίδραση ερυθρά προβάτου - αντίσωμα αντιερυθρών προβάτου και προκαλεί λύση των ερυθρών.")
40
Ο τίτλος αντισώματος στην 1η γραμμή είναι μεταξύ 1/8 και 1/16 , ενώ στη 2η γραμμή είναι 1/32.Πρόκειται για τον ίδιο ασθενή ο οποίος εξετάσθηκε με 15 ημέρες διαφορά ανάμεσα από το 1ο και το 2ο δείγμα. Η διαφορά αυτή του τίτλου ( περίπου 4πλάσια αύξηση) συνηγορεί υπερ της ενεργούς λοίμωξης .
 συνηγορεί υπερ της ενεργούς λοίμωξης ..")
41
Ανοσοφθορισμός Η μέθοδος αυτή, που εφαρμόζεται για την ανίχνευση αντιγόνων ή αντισωμάτων , χρησιμοποιεί αντίσωμα συνδεδεμένο με φλουοροσκεϊνη. Η φλουοροσκεϊνη είναι μια χρωστική η οποία όταν ακτινοβοληθεί με υπεριώδες φως, μήκους κύματος nm, εκπέμπει ορατό φως μεγαλύτερου μήκους κύματος (525 nm). Τα προς ανίχνευση αντιγόνα (βακτήρια, κύτταρα μολυσμένα με ιούς) είναι μονιμοποιημένα σε αντικει- μενοφόρους πλάκες και επικαλύπτονται με τα αντισώματα ορού ασθενούς ή από πολυκλωνικό ή μονοκλωνικό αντίσωμα ή από το φθορίζον αντίσωμα, ανάλογα με την μέθοδο που πρόκειται να εφαρμόσουμε : άμεσος ανοσοφθορισμός έμμεσος ανοσοφθορισμός
. Τα προς ανίχνευση αντιγόνα (βακτήρια, κύτταρα μολυσμένα με ιούς) είναι μονιμοποιημένα σε αντικει- μενοφόρους πλάκες και επικαλύπτονται με τα αντισώματα ορού ασθενούς ή από πολυκλωνικό ή μονοκλωνικό αντίσωμα ή από το φθορίζον αντίσωμα, ανάλογα με την μέθοδο που πρόκειται να εφαρμόσουμε : άμεσος ανοσοφθορισμός. έμμεσος ανοσοφθορισμός.")
42
Συνήθως πραγματοποιούμε τον έμμεσο ανοσοφθορισμό διότι :
η τεχνική αυτή είναι πιο ευαίσθητη από τον άμεσο ανοσοφθορισμό μια που δίνεται η δυνατότητα στο 2ο αντίσωμα να συνδεθεί με δυο αντισώματα πάνω στο 1ο ήδη συνδεδεμένο αντίσωμα επιτυγχάνοντας με αυτό τον τρόπο πολλαπλασιασμό του σήματος και μας απαλλάσει από την σήμανση κάθε 1ου αντισώματος με την φλουορεσκείνη μια που είναι πολύ πιο εύκολο να έχουμε σημάνει το 2ο αντίσωμα π.χ. αντι-ΙgG ανθρώπου ή ποντικού κ.ο.κ. Η τεχνική που ακολουθείται π.χ. για την ανίχνευση του ιού HSV σε μολυσμένα κύτταρα είναι η εξής: Τα μονιμοποιημένα κύτταρα σε αντικειμενοφόρο πλάκα επωάζονται επί 60 λεπτά στους 37οC με μονοκλωνικό ή πολυκλωνικό αντίσωμα έναντι του ιού HSV.Εν συνεχεία η πλάκα πλένεται 3 φορές με PBS και ακολουθεί 2η επώαση για 30 λεπτά με το 2ο αντίσωμα σημασμένο με φλουορεσκείνη.Η πλάκα πλένεται και πάλι ως ανωτέρω και παρατηρείται στο μικροσκόπιο φθορισμού.

43
ΕLISA (Enzyme Linked Immuno-Sorbent Assay)
Η ίδια έμμεσος τεχνική μπορεί να εφαρμοσθεί και για την ανίχνευση IgG, IgM και IgA αντισωμάτων στον ορό ασθενούς αρκεί το 2ο αντίσωμα το οποίο έχει σημανθεί με φλουορεσκείνη να αναγνωρίζει το Fc των ανοσοσφαιρινών IgG ή IgM ή IgA. Η τεχνική αυτή χρησιμοποιείται ευρύτατα για την διάγνωση της λοίμωξης από τον ιό EBV . ΕLISA (Enzyme Linked Immuno-Sorbent Assay) Mε τον έλεγχο αυτό γίνεται ανίχνευση ειδικών αντισωμάτων στον ορό, εφ’ όσον υπάρχει διαθέσιμο το αντίστοιχο αντιγόνο και αντίστροφα, ανίχνευση του αντιγόνου, εφ’ όσον είναι διαθέσιμα τα αντίστοιχα αντισώματα. Ο έλεγχος ELISA ως προς την αρχή του μοιάζει με τον ανοσοφθορισμό, αφού γίνεται χρήση αντισώματος που έχει συνδεδεμένο στο μόριό του, αντί φλουοροσκεϊνης, ένζυμο που φέρει σε πέρας μια χημική αντίδραση η οποία συνοδεύεται με μεταβολή του χρώματος του υποστρώματος της αντίδρασης.Τα συνδεδεμένα στο αντίσωμα ένζυμα που συνήθως χρησιμοποιούνται στον έλεγχο ELISA είναι η αλκαλική φωσφατάση με υπόστρωμα το p-nitrophenyl phosphate και η υπεροξειδάση με υπόστρωμα το H2O2 και την o-phenylene diamine 2 HCl. Στην άμεσο μέθοδο τα αντισώματα του αντιορού είναι συνδεδεμένα με το κατάλληλο ένζυμο.H μέθοδος αυτή χρησιμοποιείται για την άμεση ανίχνευση αντιγόνου το οποίο έχει ήδη προσροφηθεί σε μικροπλάκα πολυστυρενίου.Το σημασμένο αντίσωμα με το ειδικό ένζυμο (αλκαλική φωσφατάση ή υπεροξειδάση) συνδέεται με το αντιγόνο και η αντίδραση εμφανίζεται μέσω της κατάλυσης της ενζυμικής αντίδρασης.
 Mε τον έλεγχο αυτό γίνεται ανίχνευση ειδικών αντισωμάτων στον ορό, εφ’ όσον υπάρχει διαθέσιμο το αντίστοιχο αντιγόνο και αντίστροφα, ανίχνευση του αντιγόνου, εφ’ όσον είναι διαθέσιμα τα αντίστοιχα αντισώματα. Ο έλεγχος ELISA ως προς την αρχή του μοιάζει με τον ανοσοφθορισμό, αφού γίνεται χρήση αντισώματος που έχει συνδεδεμένο στο μόριό του, αντί φλουοροσκεϊνης, ένζυμο που φέρει σε πέρας μια χημική αντίδραση η οποία συνοδεύεται με μεταβολή του χρώματος του υποστρώματος της αντίδρασης.Τα συνδεδεμένα στο αντίσωμα ένζυμα που συνήθως χρησιμοποιούνται στον έλεγχο ELISA είναι η αλκαλική φωσφατάση με υπόστρωμα το p-nitrophenyl phosphate και η υπεροξειδάση με υπόστρωμα το H2O2 και την o-phenylene diamine 2 HCl. Στην άμεσο μέθοδο τα αντισώματα του αντιορού είναι συνδεδεμένα με το κατάλληλο ένζυμο.H μέθοδος αυτή χρησιμοποιείται για την άμεση ανίχνευση αντιγόνου το οποίο έχει ήδη προσροφηθεί σε μικροπλάκα πολυστυρενίου.Το σημασμένο αντίσωμα με το ειδικό ένζυμο (αλκαλική φωσφατάση ή υπεροξειδάση) συνδέεται με το αντιγόνο και η αντίδραση εμφανίζεται μέσω της κατάλυσης της ενζυμικής αντίδρασης.")
44
Για την έμμεσο ELISA η οποία εφαρμόζεται και συνηθέστερα ( για τους ίδιους λόγους που αναφέρθηκαν ανωτέρω), το αντιγόνο έχει ήδη προσροφηθεί στην μικροπλάκα και η επώαση με τον ειδικό αντιορό ακολουθείται από αρκετά πλυσίματα με PBS-0,1% Tween 20 για την απομάκρυνση των μη συνδεδεμένων αντισωμάτων και στη συνέχεια γίνεται η επώαση με αντι-αντίσωμα το οποίο είναι συνδεδεμένο με το ειδικό ένζυμο (αλκαλική φωσφατάση ή υπεροξειδάση) και η ενζυμική αντίδραση εμφανίζεται ως ανωτέρω.
, το αντιγόνο έχει ήδη προσροφηθεί στην μικροπλάκα και η επώαση με τον ειδικό αντιορό ακολουθείται από αρκετά πλυσίματα με PBS-0,1% Tween 20 για την απομάκρυνση των μη συνδεδεμένων αντισωμάτων και στη συνέχεια γίνεται η επώαση με αντι-αντίσωμα το οποίο είναι συνδεδεμένο με το ειδικό ένζυμο (αλκαλική φωσφατάση ή υπεροξειδάση) και η ενζυμική αντίδραση εμφανίζεται ως ανωτέρω.")
45
Είναι σημαντικό σε όλες αυτές τις αντιδράσεις να αποδειχθεί ότι τόσο το 1ο αντίσωμα όσο και το δεύτερο δεν συνδέονται μη ειδικά στο πλαστικό της μικροπλάκας.Γι ‘αυτό το λόγο μετά την προσρόφηση του αντιγόνου η μικροπλάκα καλύπτεται με 1% βόειο αλβουμίνη σε PBS . Μικροπλάκα ELISA Oσο πιο έντονο το κίτρινο χρώμα τόσο πιο θετικό το δείγμα . Με λευκό τα αρνητικά δείγματα.

46
Capture ELISAs - Antigen Capture
- Antibody Capture Antigen Capture Η τεχνική αυτή εφαρμόζεται για την ανίχνευση αντιγόνου έχοντας ήδη προσροφήσει στη μικροπλάκα ένα 1ο αντίσωμα το οποίο αναγνωρίζει το προς ανίχνευση αντιγόνο. Εν συνεχεία ακολουθούν πλύσεις ως ανωτέρω , επωάζουμε με ένα 2ο αντίσωμα σημασμένο με το ειδικό ένζυμο (αλκαλική φωσφατάση ή υπεροξειδάση) και η ενζυμική αντίδραση εμφανίζεται ως ανωτέρω. Απαραίτητη προυπόθεση για να λειτουργήσει σωστά η τεχνική αυτή είναι το 2ο αντίσωμα να αναγνωρίζει διαφορετικούς επίτοπους από το 1ο αντίσωμα.
 και η ενζυμική αντίδραση εμφανίζεται ως ανωτέρω. Απαραίτητη προυπόθεση για να λειτουργήσει σωστά η τεχνική αυτή είναι το 2ο αντίσωμα να αναγνωρίζει διαφορετικούς επίτοπους από το 1ο αντίσωμα.")
47
Antibody Capture Στην τεχνική αυτή ένα 1ο αντίσωμα το οποίο έχει ήδη προσροφηθεί στη μικροπλάκα χρησιμοποιείται για να αναγνωρίσει αντισώματα της τάξης των IgG, IgA ή IgM. Προσθέτουμε εν συνεχεία τον ορό του ασθενούς και επωάζουμε.Στην περίπτωση όπου το 1ο αντίσωμα είναι αντι - IgM θα αναγνωρίσει και θα δεσμεύσει τα IgM του ορού. Πλένουμε την μικροπλάκα ,προσθέτουμε το αντιγόνο , επωάζουμε , πλένουμε και πάλι και τέλος προσθέτουμε το σημασμένο με ένζυμο αντίσωμα έναντι του αντιγόνου και εμφανίζουμε την αντίδραση ως ανωτέρω

48
ΦΥΣΙΟΛΟΓΙΚΗ ΑΝΤΙΔΡΑΣΗ ΨΕΥΔΩΣ ΘΕΤΙΚΗ ΑΝΤΙΔΡΑΣΗΣΗ
Η τεχνική αυτή βρίσκει ευρύτατη εφαρμογή για την διάγνωση των IgM αντισωμάτων ( ένδειξη πρόσφατης λοίμωξης) για δύο κυρίως λόγους. Αν εφαρμοσθεί η έμμεσος τεχνική ELISA τότε τα IgM αντισώματα ανταγωνίζονται τις θέσεις πρόσδεσης στο αντιγόνο και δεδομένου ότι ευρίσκονται σε συγκέντρωση 15mg/ml ενώ τα IgM σε συγκέντρωση 1mg/ml υπερισχύουν πάντοτε των IgM με αποτέλεσμα ψευδώς αρνητικά αποτελέσματα. Ομοίως αν εφαρμοσθεί η έμμεσος τεχνική ELISA ο ρευματοειδής παράγων (RF) ο οποίος είναι συνήθως της τάξης των IgM με ειδικότητα αντι- IgG συνδέεται στα ειδικά έναντι του αντιγόνου IgG αντισώματα τα οποία έχουν συνδεθεί στο αντιγόνο .Οταν προσθέτουμε τα σημασμένα με το ένζυμο αντι-IgM αντισώματα τότε αυτά συνδέονται στο σύμπλοκο RF-IgG και κατά συνέπεια έχουμε ψευδώς θετική IgM αντίδραση. ΦΥΣΙΟΛΟΓΙΚΗ ΑΝΤΙΔΡΑΣΗ ΨΕΥΔΩΣ ΘΕΤΙΚΗ ΑΝΤΙΔΡΑΣΗΣΗ
 για δύο κυρίως λόγους. Αν εφαρμοσθεί η έμμεσος τεχνική ELISA τότε τα IgM αντισώματα ανταγωνίζονται τις θέσεις πρόσδεσης στο αντιγόνο και δεδομένου ότι ευρίσκονται σε συγκέντρωση 15mg/ml ενώ τα IgM σε συγκέντρωση 1mg/ml υπερισχύουν πάντοτε των IgM με αποτέλεσμα ψευδώς αρνητικά αποτελέσματα. Ομοίως αν εφαρμοσθεί η έμμεσος τεχνική ELISA ο ρευματοειδής παράγων (RF) ο οποίος είναι συνήθως της τάξης των IgM με ειδικότητα αντι- IgG συνδέεται στα ειδικά έναντι του αντιγόνου IgG αντισώματα τα οποία έχουν συνδεθεί στο αντιγόνο .Οταν προσθέτουμε τα σημασμένα με το ένζυμο αντι-IgM αντισώματα τότε αυτά συνδέονται στο σύμπλοκο RF-IgG και κατά συνέπεια έχουμε ψευδώς θετική IgM αντίδραση. ΦΥΣΙΟΛΟΓΙΚΗ ΑΝΤΙΔΡΑΣΗ ΨΕΥΔΩΣ ΘΕΤΙΚΗ ΑΝΤΙΔΡΑΣΗΣΗ.")
49
ΠΡΩΤΟΓΕΝΗΣ ΚΑΙ ΔΕΥΤΕΡΟΓΕΝΗΣ ΑΝΟΣΟΛΟΓΙΚΗ ΑΠΑΝΤΗΣΗ ΕΝΑΝΤΙ ΒΑΚΤΗΡΙΩΝ Η/ΚΑΙ ΙΩΝ

50
Η μέθοδος Western blotting περιλαμβάνει 3 στάδια: ηλεκτροφόρηση
των αντιγόνων σε πηκτή πολυακρυλαμίδης, ηλεκτρομεταφορά των ζωνών που σχηματίζονται σε φύλλο νιτροκυτταρίνης (electroblotting) και ανίχνευση του αντιγόνου. Ηλεκτροφόρηση αντιγόνων Η ανάλυση των αντιγόνων με ηλεκτροφόρηση γίνεται σε πηκτή πολυμερισμένης ακρυλαμίδης-bis ακρυλαμίδης-SDS στην οποία, ανάλογα του μεγέθους των πεπτιδίων που πρόκειται να διαχωρισθούν, το ποσοστό της ακρυλαμίδης κυμαίνεται από 5 μέχρι 20%. Βάσει της ηλεκτροφόρησης σε SDS-PAGE οι πρωτεϊνες χωρίζουν μόνο βάσει του μοριακού τους βάρους μια που οι δευτεροταγείς, τριτοταγείς και τεταρτοταγείς δομές τους έχουν καταστραφεί από το SDS (Sodium Dodecyl Sulfate) , ένα ισχυρό απορρυπαντικό το οποίο αποδιατάσσει τις πρωτείνες και τους προσθέτει και αρνητικό φορτίο ( sulfate). Mε αυτό τον τρόπο οι πρωτείνες διατηρούν μόνο την πρωτοταγή δομή τους και έχουν όλες ένα σαφώς αρνητικό φορτίο που τους επιβάλλει να ταξιδεύσουν από τον αρνητικό προς τον θετικό πόλο κατά την διάρκεια της ηλεκτροφόρησης.
 και ανίχνευση του αντιγόνου. Ηλεκτροφόρηση αντιγόνων. Η ανάλυση των αντιγόνων με ηλεκτροφόρηση γίνεται σε πηκτή πολυμερισμένης ακρυλαμίδης-bis ακρυλαμίδης-SDS στην οποία, ανάλογα του μεγέθους των πεπτιδίων που πρόκειται να διαχωρισθούν, το ποσοστό. της ακρυλαμίδης κυμαίνεται από 5 μέχρι 20%. Βάσει της ηλεκτροφόρησης σε SDS-PAGE οι πρωτεϊνες χωρίζουν μόνο βάσει του μοριακού τους βάρους μια που οι δευτεροταγείς, τριτοταγείς και τεταρτοταγείς δομές τους έχουν καταστραφεί από το SDS (Sodium Dodecyl Sulfate) , ένα ισχυρό απορρυπαντικό το οποίο αποδιατάσσει τις πρωτείνες και τους προσθέτει και αρνητικό φορτίο ( sulfate). Mε αυτό τον τρόπο οι πρωτείνες διατηρούν μόνο την πρωτοταγή δομή τους και έχουν όλες ένα σαφώς αρνητικό φορτίο που τους επιβάλλει να ταξιδεύσουν από τον αρνητικό προς τον θετικό πόλο κατά την διάρκεια της ηλεκτροφόρησης.")
51
Όταν η ηλεκτροφόρηση γίνεται σε πηκτή πολυμερισμένης ακρυλαμίδης-bis ακρυλαμίδης ( PAGE) το πήκτωμα αποτελείται από ένα λαβύρινθο καναλιών τα οποία επεκτείνονται από την αρχή μέχρι το τέλος του πηκτώματος η δε διάμετρος τους προσδιορίζεται από την συγκέντρωση του ακρυλαμιδίου- bis ακρυλαμιδίου
 το πήκτωμα αποτελείται από ένα λαβύρινθο καναλιών τα οποία επεκτείνονται από την αρχή μέχρι το τέλος του πηκτώματος η δε διάμετρος τους προσδιορίζεται από την συγκέντρωση του ακρυλαμιδίου- bis ακρυλαμιδίου")
52
Όταν οι αποδιατεταγμένες πρωτείνες υποβάλλονται σε ηλεκτροφόρηση στο ανωτέρω πήκτωμα θα ταξιδεύσουν από τον αρνητικό προς τον θετικό πόλο βάσει μόνο του μοριακού τους βάρους.Κατά συνέπεια αν 2 πολυπεπτίδια έχουν το ίδιο μοριακό βάρος αλλά διαφορετική σύσταση αμινοξέων ( είναι δηλ. διαφορετικά πολυπεπτίδια) θα ταξιδεύσουν ως μια ζώνη
 θα ταξιδεύσουν ως μια ζώνη.")
53
Μετά το τέλος της ηλεκτροφόρησης βάφουμε το πήκτωμα συνήθως με Coomassie blue όπως κατωτέρω .
Η συγκέντρωση του συνόλου των πρωτεϊνών που τοποθετήσαμε σε κάθε γραμμή κυμαίνεται από ng ( αριστερό άκρο) μέχρι 8 ng ( δεξιό άκρο ) της εικόνας.
 μέχρι 8 ng ( δεξιό άκρο ) της εικόνας.")
54
Μεταφορά σε μεμβράνες Η μεταφορά των διαχωρισμένων πολυπεπτιδίων από την πηκτή στη μεμβράνη γίνεται με τη βοήθεια ηλεκτρικού πεδίου. Στα διάφορα συστήματα μεταφοράς τα οποία διακρίνονται σε υγρά ή ημι-στεγνά, οι συνθήκες της ηλεκτροφόρησης και μεταφοράς ποικίλουν. Για τα υγρά συστήματα συνήθως εφαρμόζεται ηλεκτρικό ρεύμα 300 mA για 3 ώρες ή100 mA για μια νύκτα, ενώ η μεταφορά στα ημι-στεγνά συστήματα γίνεται γρηγορότερα λεπτά στα V. Υγρό σύστημα μεταφοράς σε νιτροκυτταρίνη. Δημιουργούμε ένα «sandwich» ανάμεσα στην πηκτή και την νιτροκυτταρίνη και το υποβάλλουμε σε ηλεκτροφόρηση στα 100 mA για μια νύκτα.

55
Σε ορισμένες περιπτώσεις εφαρμογών που η ηλεκτροφόρηση σε πολυακρυλαμίδη δεν θεωρείται απαραίτητη, η σύνδεση των αντιγόνων με τη νιτροκυτταρίνη γίνεται απευθείας με εναπόθεσή τους σ’ αυτή, χωρίς μετουσίωση, υπό μορφή κηλίδων (μέθοδος γνωστή ως dot blotting) με την βοήθεια κενού.Το κενό επιταχύνει την σύνδεση των πρωτεϊνών στην νιτροκυτταρίνη και διευκολύνει τα επακόλουθα πλυσίματα της νιτροκυτταρίνης. Συσκευή Dot-Blotting 96 δεξαμενών. Στην θέση 5 τοποθετείται η νιτροκυτταρίνη.

56
Aνίχνευση Το ειδικό αντίσωμα στην άμεση παραλλαγή της μεθόδου είναι συνδεδεμένο με το ένζυμο ανίχνευσης ( π.χ. υπεροξειδάση). Στην έμμεση παραλλαγή της μεθόδου το σύστημα ανίχνευσης βρίσκεται στο 2ο αντίσωμα ( αντι-αντίσωμα ) που μπορεί να είναι η αλκαλική φωσφατάση ή υπεροξειδάση ή ενδεχομένως η πρωτεϊνη Α του Staphylococcus aureus η οποία είναι ραδιενεργά σημασμένη (με ιώδιο Ι125). Ακολουθεί η εμφάνιση της μεμβράνης η οποία, ανάλογα με το σύστημα ανίχνευσης που χρησιμοποιείται, γίνεται με διαφορετικό τρόπο ( ενζυμική αντίδραση ή αυτοραδιογραφία).
. Στην έμμεση παραλλαγή της μεθόδου το σύστημα ανίχνευσης βρίσκεται στο 2ο αντίσωμα ( αντι-αντίσωμα ) που μπορεί να είναι η αλκαλική φωσφατάση ή υπεροξειδάση ή ενδεχομένως η πρωτεϊνη Α του Staphylococcus aureus η οποία είναι ραδιενεργά σημασμένη (με ιώδιο Ι125). Ακολουθεί η εμφάνιση της μεμβράνης η οποία, ανάλογα με το σύστημα ανίχνευσης που χρησιμοποιείται, γίνεται με διαφορετικό τρόπο ( ενζυμική αντίδραση ή αυτοραδιογραφία).")
57
Η ανωτέρω τεχνική εκτός της εφαρμογής της σε ερευνητικά πρωτόκολλα , βρίσκει μεγάλη εφαρμογή στην διαγνωστική Ιολογία ως επιβεβαιωτικό τεστ για κάθε ELISA έναντι των ιών ΗΙV 1 & 2. Για να χαρακτηρισθεί ως HIV θετικός ένας φορέας θα πρέπει τα αντισώματά του να αναγνωρίζουν τουλάχιστον 4 πολυπεπτίδια του ιού που κωδικοποιούνται από τα γονίδια Gag, Pol και Env.












>")









