Κατέβασμα παρουσίασης
1
Fiziologija životinja

2
FIZIOLOGIJA physis (priroda) i logos (nauka) Biološka disciplina koja proučava funkcije živih organizama kao i fizičke i hemijske procese koji leže u osnovi tih funkcija. ZADATAK: utvrđivanje zakonitosti po kojima se životni procesi odvijaju, da ih opiše i objasni, kao i da utvrđivaje regulacionih mehanizama životnih procesa PREDMET izučavanja: svi nivoi organizacije živih sistema (ćelije, organi, organski sistemi) kao i organizam u celini
 kao i organizam u celini.")
3
Discipline fiziologije
fiziologija životinja i fiziologija biljaka FIZIOLOGIJA ŽIVOTINJA opšta i sistemska fiziologija (osnovni principi po kojima različiti životinjski organizmi funkcionišu) uporedna fiziologija (sličnosti i razlike između vrsta) ekološka fiziologija (interakcije organizama sa sredinom) evoluciona fiziologija (promene pojedinih fizioloških osobina kroz vreme i nastanah novih pod uticajem prirodne selekcije) fiziologija čoveka patološka fiziologija (funkcionalni poremećaji u ljudskom organizmu)
 uporedna fiziologija (sličnosti i razlike između vrsta) ekološka fiziologija (interakcije organizama sa sredinom) evoluciona fiziologija (promene pojedinih fizioloških osobina kroz vreme i nastanah novih pod uticajem prirodne selekcije) fiziologija čoveka. patološka fiziologija (funkcionalni poremećaji u ljudskom organizmu)")
4
Veza fiziologije sa drugim naukama
Anatomija da bi se proučili životni procesi, neophodno je poznavanje građe ćelija, organa, organizma. Fizika i hemija životni procesi koji se odigravaju u ćeliji i organizmu odigravaju se po zakonima fizike i hemije npr. disanje, razmena gasova, delovanje enzima, procesi u nervnim i mišićnim ćelijama U sistemu bioloških disciplina fiziologija je posebno povezana sa biohemijom i biofizikom.

5
Značaj fiziologije fundamentalni (teorijski) značaj praktični značaj
uočavanje sličnosti i razlika između vrsta praktični značaj primena eksperimentalnih nalaza sa životinja na čoveka

6
Nivoi organizacije živih sistema

7
ORGANIZAM SISTEM ORGANA ORGANSKI APARAT ORGAN TKIVO ĆELIJA
biosfera biom ekosistem biocenoza populacija ORGANIZAM SISTEM ORGANA ORGANSKI APARAT ORGAN TKIVO ĆELIJA organele biomolekuli

8
Ćelija osnovna jedinica građe i funkcije svakog organizma

9
Tkivo Viši nivo organizacije. Ćelije se integrišu u tkivo
Tkivo je grupa ćelija istog oblika, građe, funkcije i embrionalnog porekla.

10
Organ Viši nivo organizacije žive materije od tkiva.
U sastav jednog organa ulazi više vrsta tkiva. Svaki organ vrši određenu funkciju npr. pluća -razmenu gasova; bubrezi-izlučivanje; srce-cirkulaciju krvi.

11
Organski aparat Čine organi koji su funkcionalno i anatomski povezani.
Svaki organ u okviru aparata vrši određenu funkciju koja je podređena organskom aparatu. Organski aparati su čulo vida (oko) i čulo sluha i ravnoteže (uho).
 i čulo sluha i ravnoteže (uho).")
12
Organski sistem Organi su funkcionalno i anatomski povezani (npr. sistem organa za varenje, disanje, krvotok). Organski sistem mogu graditi organi među kojima postoji samo funkcionalna veza. Npr. endokrini sistem. Endokrine žlezde su u organizmu prostorno jako udaljene, povezuju se putem krvi koja prenosi hormone.

13
Organizam Najviši nivo žive materije
Ostvaren je visok stepen integracije njegovih delova organa i organskih sistema. Organizam je strukturna i funkcionalna celina. U organizmu postoji koordinirano, skladno delovanje njegovih organa i organskih sistema npr. poremećaj u radu jednog organa može se odraziti i na rad drugih organa a to se može odraziti na nivou organizma kao celine.

14
Opšte i specijalizovane ćelijske funkcije
Procesi pomoću kojih ćelija obezbeđuje minimalne potrebe za održavanje integriteta i života nazivaju se opšti ili fundamentalni ćelijski procesi. - transprot metabolita i jona kroz membranu - metabolički procesi u ćeliji - sinteza proteina - umnožavanje... Posebni procesi koji se odvijaju samo u određenom tipu ćelija nazivaju specijalizovani ćelijski procesi i oni po pravilu predstavljaju specijalizaciju jednog ili više fundamentalnih ćelijskih procesa. - izlučivanje neurotransmitera (nervne ćelije) ili hormona (žlezdane ćelije) - geneza akcionih potencijala - sinteza specifičnih proteina Koordinirane telesne funkcije
 ili hormona (žlezdane ćelije) - geneza akcionih potencijala. - sinteza specifičnih proteina. Koordinirane. telesne funkcije.")
15
Integracija Visok stepen integracije delova na nivou organa, kao i njihovo koordinirano delovanje obezbeđuju dva sistema u organizmu: endokrini i nervni sistem.

16
Interna sredina i homeostaza
Ukupan efekat aktivnosti sistema organa je takav da se unutar organizma kreira interna sredina neophodna svim ćelijama za normalno funkcionisanje. Regulacija interne sredine u relativno uskim granicama ili težnja za održavanjem stabilnosti stanja interne sredine, naziva se HOMEOSTAZA. Homeostaza održava relativnu nezavisnost organizma od spoljašnje sredine i uslov je njegovog opstanka - Ćelijska homeostaza - Telesna homeostaza - Imunološka homeostaza - Hemostaza Klod Bernar ( )
")
17
Biološki kontrolni sistemi
Koordinirano odigravanje opštih i specijalizovanih ćelijskih funkcija, funkcija tkiva, organa i sistema organa, odnosno, odigravanje koordiniranih telesnih procesa odvija se zahvaljujući postojanju preciznih bioloških kontrolnih sistema. Biološki kontrolni sistemi postoje na svim nivoima organizacije, od ćelijskog nivoa do nivoa organizma. Biološki kontrolni sistemi funkcionišu na principu povratne sprege.

18
Negativna povratna sprega Pozitivna povratna sprega
Hladnoća Prednji režanj hipofize hipotalamus Štitna žlezda Negativna povratna sprega Tireotropin Tireotropni oslobađajući faktor Tiroksin Primer: Regulacija lučenja hormona (hipotalamus-hipofiza-žlezda) Primer: Kaskadni proces koagulacije krvi
 Primer: Kaskadni proces koagulacije krvi.")
19
Integracija ćelijskih funkcija uspostavlja se zahvaljujući međusobnoj ćelijskoj komunikaciji i interakciji Osnovni tipovi međućelijskih interakcija Neposredna međućelijska komunikacija koja se ostvaruje između susednih ćelija. Lokalna signalizacija - lokalno otpuštanje signalnih molekula koji difunduju do susednih ćelija Nervne ćelije omogućavaju direktnu komunikaciju kako sa bliskim, tako i sa udaljenim ćelijama u organizmu. Endokrina signalizacija- ostvaruje se posredstvom hormona .

20
Molekulski mehanizmi međućelijskih interakcija
Svi tipovi međućelijskih interakcija zahtevaju prisustvo: signalnog molekula (primarnog glasnika) receptorskog molekula
 receptorskog molekula.")
21
površinski (membranski)
RECEPTORI: unutarćelijski površinski (membranski) Signalni molekul može da prođe kroz ćelijsku membranu (steroidni i tiroidni hormoni) Signalni molekul ne može da prođe kroz ćelijsku membranu (neurotransmiteri, faktori rasta, proteinski hormoni)
 Signalni molekul može da prođe kroz ćelijsku membranu (steroidni i tiroidni hormoni) Signalni molekul ne može da prođe kroz ćelijsku membranu (neurotransmiteri, faktori rasta, proteinski hormoni)")
22
Hemijski sastav ćelije

23
НЕОРГАНСКE МАТЕРИЈE ВОДА МАКРОЕЛЕМЕНТИ МИКРОЕЛЕМЕНТИ
кисеоник, водоник, угљеник, азот, фосфор, натријум, калијум, калцијум, магнезијум, гвожђе МИКРОЕЛЕМЕНТИ јод, бакар, манган, кобалт, цинк АНЈОНИ КАТЈОНИ Na+, K+, Mg+, Ca++ Cl-, SO4--, PO4--

24
ЧЕТИРИ ГЛАВНА ТИПА МАКРОМОЛЕКУЛА КОЈИ СЕ НАЛАЗЕ У ЋЕЛИЈИ
ЕЛЕМЕНТИ МОНОМЕРИ (Градивни блокови) ПОЛИМЕР ПРИМЕР УЛОГА Угљени хидрати C, H, O Прости шећери (10) поли сахарид скроб, гликоген, целулоза, хитин Депои енергије, структурна (ћелијски зидови, егзоскелет) Липиди Масне киселине (10) и глицерол липид масти, уља, стероиди, фосфо липиди Депои енергије, структурна (ћелијска мембрана) Протеини C, H, O, N, S Амино киселине (20) пептиди Кератин, пепсин, инсулин Структурна Ензими Хормони Антитела Нуклеинске киселине C, H, O, P Нуклеотиди (5) нуклеинске киселине DNA, RNA Информациона
 ПОЛИМЕР. ПРИМЕР. УЛОГА. Угљени хидрати. C, H, O. Прости шећери (10) поли. сахарид. скроб, гликоген, целулоза, хитин. Депои енергије, структурна (ћелијски зидови, егзоскелет) Липиди. Масне киселине (10) и глицерол. липид. масти, уља, стероиди, фосфо. липиди. Депои енергије, структурна (ћелијска мембрана) Протеини. C, H, O, N, S. Амино киселине (20) пептиди. Кератин, пепсин, инсулин. Структурна. Ензими. Хормони. Антитела. Нуклеинске киселине. C, H, O, P. Нуклеотиди (5) нуклеинске киселине. DNA, RNA. Информациона.")
25
УГЉЕНИ ХИДРАТИ ПОЛИСАХАРИДИ ДИСАХАРИДИ РЕЗЕРВНИ СТРУКТУРНИ
МОНОСАХАРИДИ ТРИОЗЕ ПЕНТОЗЕ рибоза дезоксирибоза ХЕКСОЗЕ ГЛУКОЗА ФРУКТОЗА галактоза ДИСАХАРИДИ МАЛТОЗА САХАРОЗА лактоза ПОЛИСАХАРИДИ СТРУКТУРНИ ЦЕЛУЛОЗА - биљке ХИТИН - животиње РЕЗЕРВНИ СКРОБ - биљке ГЛИКОГЕН - животиње

26
МАСТИ (ЛИПИДИ) ФОСФОЛИПИДИ ТРИГЛИЦЕРИДИ СТЕРОИДИ
 ФОСФОЛИПИДИ ТРИГЛИЦЕРИДИ СТЕРОИДИ")
27
ПРОТЕИНИ СТРУКТУРНИ ЕНЗИМИ ХОРМОНИ АНТИТЕЛА КЕРАТИН АМИЛАЗА ИНСУЛИН
ИМУНОГЛОБУЛИНИ

28
Карбоксилна група (-COOH)
Aмино група (-NH2) -R остатак има различиту структуру и особине (растворљивост, наелектрисање, елементарни састав) Општа структура амино киселина
 -R остатак има различиту структуру и особине (растворљивост, наелектрисање, елементарни састав) Општа структура амино киселина.")
29
N- терминус С- терминус Пептидна веза

31
KVATERNERNA STRUKTURA
PRIMARNA STRUKTURA linearna sekvenca amino kiselina SEKUNDARNA STRUKTURA - α heliks - nepravilno klupče - β paralelne ploče TERCIJARNA STRUKTURA Uvijeni lanac KVATERNERNA STRUKTURA Dva ili više polipeptidna lanca

32
КВЕРТЕНЕРНА СТРУКТУРА
ПРИМАРНА СТРУКТУРА секвенца АК (редослед и састав!!!) СЕКУНДАРНА СТРУКТУРА α хеликс, β паралелне плоче ТЕРЦИЈАРНА СТРУКТУРА линеарни или глобуларни КВЕРТЕНЕРНА СТРУКТУРА више субјединица
 СЕКУНДАРНА СТРУКТУРА. α хеликс, β паралелне плоче. ТЕРЦИЈАРНА СТРУКТУРА. линеарни или глобуларни. КВЕРТЕНЕРНА СТРУКТУРА. више субјединица.")
33
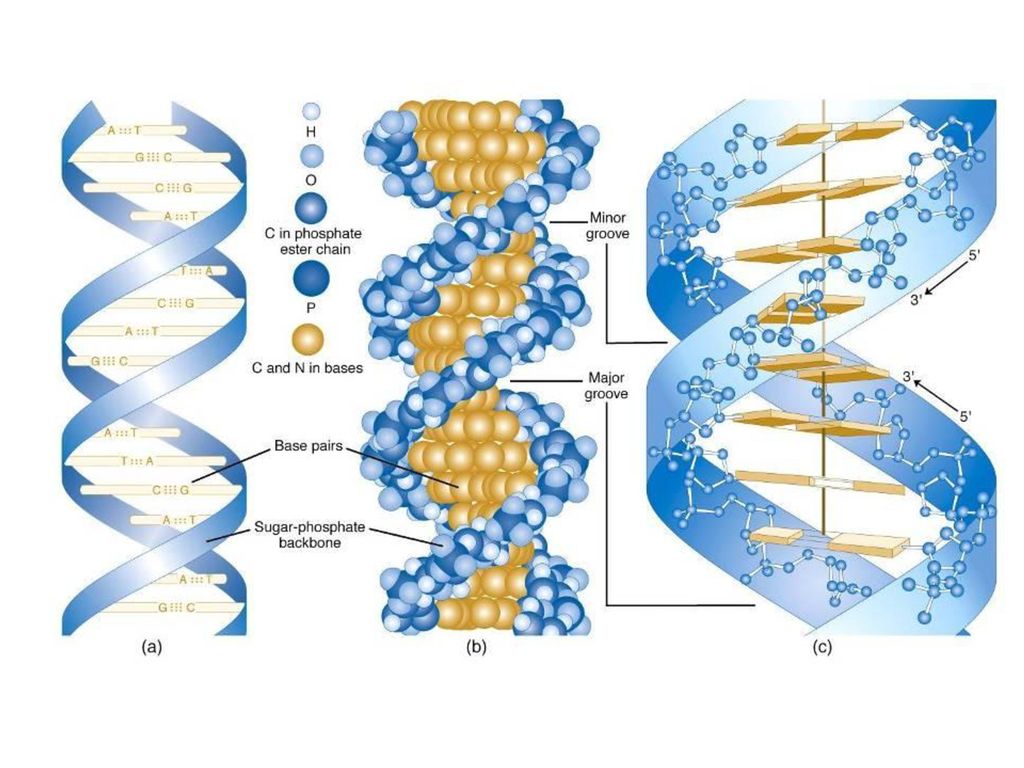
НУКЛЕИНСКЕ КИСЕЛИНЕ

34
Грађа ДНК Молекул ДНК се састоји из два комплементарна ланца нуклеотида, односно од два полинуклеотидна ланца који се спирално увијају и повезани су водоничним везама. Сваки нуклеотид састоји се из три молекула пентоза десоксирибоза азотна база (пуринске: аденин и гуанин и пиримидинске: цитозин и тимин) фосфорна киселина или фосфатна група
 фосфорна киселина или фосфатна група.")
35
NUKLEOTID Dezoksiribonukleotid fosfat timin dezoksiriboza

36
Komplementarno sparivanje baznih parova
timin A adenin C citozin G guanin Komplementarno sparivanje baznih parova

37
Vodonične veze Fosfodiestarske veze

39
Грађа RNK РНК гради 1 полинуклеотидни ланац
Нуклеотид РНК се састоји од: пентозе рибозе азотне базе (пуринске: аденин и гуанин; пиримидинске: цитозин и урацил) фосфатне групе.
 фосфатне групе.")
40
Постоје 3 типа РНК: информациона РНК (iRNK, mRNK - messenger)
рибозомална РНК (rRNK) транспортна РНК (tRNK)
 транспортна РНК (tRNK)")
41
Разлике молекула РНК и ДНК
молекул је једноланчан, садржи шећер рибозу у нуклеотидима уместо тимина, садржи урацил. ДНК молекул је дволанчан, садржи шећер дезоксирибозу у нуклеотидима

42
Uloga ćelijskih organela

43
Animalna ćelija Citoskelet Plazma membrana
Glatki endoplazmatični retikulum Granularni endoplazmatični retikulum Jedro Bič Citoskelet Lizozom Centriola Ribozomi Peroksizom Goldžijev aparat Mikrotubule Plazma membrana Intermedijarni filamenti Citoskelet Mikrofilamenti Mitohondrija

44
ЋЕЛИЈСКЕ ОРГАНЕЛЕ-УНУТРАШЊЕ МЕМБРАНЕ
ОРГАНЕЛЕ КОЈЕ УЧЕСТВУЈУ У ПРОЦЕСИМА СИНТЕЗЕ РИБОЗОМИ, ЕНДОПЛАЗМАТИЧНИ РЕТИКУЛУМ, ГОЛЏИЈЕВ КОМПЛЕКС

45
ЋЕЛИЈСКЕ ОРГАНЕЛЕ-УНУТРАШЊЕ МЕМБРАНЕ
ОРГАНЕЛЕ У КОЈИМА СЕ СИНТЕТИШЕ ATP: МИТОХОНДРИЈЕ

46
ЋЕЛИЈСКЕ ОРГАНЕЛЕ-УНУТРАШЊЕ МЕМБРАНЕ
ОРГАНЕЛЕ У КОЈИМА СЕ СКЛАДИШТЕ ХИДРОЛИТИЧКИ ЕНЗИМИ ЛИЗОЗОМИ И ПЕРОКСИЗОМИ

47
ЋЕЛИЈСКИ СКЕЛЕТ (ЦИТОСКЕЛЕТ)
ЦЕНТРОЗОМ – ЦЕНТРИОЛЕ ЈЕДРО (NUCLEUS) ЋЕЛИЈСКА МЕМБРАНА (ПЛАЗМА МЕМБРАНА)
 ЋЕЛИЈСКА МЕМБРАНА (ПЛАЗМА МЕМБРАНА)")
48
Metabolizam

49
BIOLOŠKI ZNAČAJ METABOLIZMA
METABOLIZAM = KATABOLIZAM + ANABOLIZAM AMFIBOLIZAM 1 da oslobodi hemijsku energiju iz redukovanih jedinjenja KATABOLIZAM 2 da konverguje nutrijente iz spoljašnje sredine u gradivne blokove ili druge prekursore za biosintezu 3 da povezuje gradivne blokove u makromolekule ANABOLIZAM 4 da omogući formiranje i degradaciju biomolekula potrebnih za specijalne funkcije pojedinih }elija

50
CIKLUS ENERGIJE PREKO ATP-a
Redukovana jedinjenja H2O CO2 O2 KATABOLIZAM Pi ADP ATP MEHANIČKI RAD TRANSPORT BIOSINTETSKI RAD

51
DIREKTNI PRENOS ENERGIJE PREKO CIKLUSA
NADP /NADPH + Oksidovana jedinjenja Redukovana jedinjenja KATABOLIZAM NADPH + H + NADP + REDUKTIVNE BIOSINTETSKE REAKCIJE Redukovana jedinjenja Oksidovana jedinjenja

52
ENZIMOLOGIJA

53
Enzimi su katalizatori biološkog porekla
po hemijskoj prirodi su proteini u ćeliji obezbeđuju tok i odigravanje hemijskih reakcija

54
malo istorije... Enzimski procesi u organizmu su poznati od davnina (npr. sirenje sira, kišeljenje testa, prevođenje šire u vino). Naziv ‘enzim’ dao je Kun 1878.godine od reči en - u i zyme - kvasac, jer je pravilno uočio da u kvascu postoji supstanca koja katalizuje reakciju vrenja. Naučno proučavanje enzima je počelo u XIX veku kada su stvoreni temelji enzimologije tj nauke o enzimima. Prvi enzim je izolovao Samner g. i to je ureaza u kristalnom obliku - po hemijskoj prirodi je protein. Posle su izolovani i drugi enzimi proteinske prirode.

55
Hemijske reakcije u organizmu su inače moguće tj
Hemijske reakcije u organizmu su inače moguće tj. mogu se odigravati bez enzima bez enzima bi one bile spore proizvodi tih reakcija bi se javljali u smanjenoj koncentraciji Enzimi su jako efikasni katalizatori, u jedinici vremena izmene velike količine supstrata

56
Nomenklatura - ili kako dati naziv enzimu-
trivijalni nazivi: ptijalin, pepsin, tripsin, himotripsin. sistemski nazivi: enzimu se daje naziv po imenu supstrata na koji deluje, doda se samo sufiks AZA. ureaza koja deluje na ureu, amilaza deluje na amilum (skrob), arginaza deluje na arginin (aminokiselina). najnovije preporuke: enzimu se daje naziv tako da se vidi na koji supstrat deluje i koji tip reakcije katalizuje. Npr. protein hidraza (vrši hidrolizu proteina)
, arginaza deluje na arginin (aminokiselina). najnovije preporuke: enzimu se daje naziv tako da se vidi na koji supstrat deluje i koji tip reakcije katalizuje. Npr. protein hidraza (vrši hidrolizu proteina)")
57
Enzimi su svrstani u 6 grupa na osnovu hemijskih reakcija
Enzimi oksidoreduktaze (katalizuju oksidoredukciju između dva supstrata) Hidrolaze (katalizuju hidrolizu peptidnih, estarskih, etarskih i glukoznih veza; tu spadaju proteaze i lipaze). Transferaze (katalizuju prenos ili transfer funkcionalnih grupa; to su npr. aminotransferaze i fosfotransferaze) Liaze (uklanjaju određenu grupu iz supstrata pri čemu supstratu ostaje dvoguba veza) Izomeraze (katalizuju unutarmolekulsko preuređenje supstrata, npr. međusobno prevođenje optičkih i geometrijskih izomera - cis i trans oblik) Ligaze ili sintetaze (katalizuju vezivanje dva supstrata uz utrošak ATP-a)
 Hidrolaze (katalizuju hidrolizu peptidnih, estarskih, etarskih i glukoznih veza; tu spadaju proteaze i lipaze). Transferaze (katalizuju prenos ili transfer funkcionalnih grupa; to su npr. aminotransferaze i fosfotransferaze) Liaze (uklanjaju određenu grupu iz supstrata pri čemu supstratu ostaje dvoguba veza) Izomeraze (katalizuju unutarmolekulsko preuređenje supstrata, npr. međusobno prevođenje optičkih i geometrijskih izomera - cis i trans oblik) Ligaze ili sintetaze (katalizuju vezivanje dva supstrata uz utrošak ATP-a)")
58
Hemijska priroda enzima
Svi enzimi su proteini ali se na osnovu složenosti građe dele na: Protein enzime koji se sastoje samo od amino kiselina Protein enzime sa metalnim kofaktorom, u čiji sastav ulazi metalni jon i ako se on ukloni enzim gubi aktivnost. Složene enzime - jedan složeni enzim se sastoji iz apoenzima (protein) i koenzima (neproteinski deo) i naziva se još i holoenzim.
 i koenzima (neproteinski deo) i naziva se još i holoenzim.")
59
još o složenim enzimima
APOENZIM i KOENZIM su reverzibilno vezani i čine HOLOENZIM ako se apoenzim i koenzim odvoje, holoenzim gubi aktivnost. Apoenzim određuje na koji će supstrat holoenzim delovati i tip hemijske reakcije koji će katalizovati. Koenzimi se javljaju kao prenosioci atoma i atomskih grupa koje se uklanjaju sa supstrata ili se dobijaju (npr. holoenzimi su dehidrogenaze a njihovi koenzimi su NAD i FAD). Kofaktor Apoenzim Holoenzim
. Kofaktor. Apoenzim. Holoenzim.")
60
Specifičnost enzima Enzimi su specifični što znači da deluju na tačno određene supstrate u tačno određenim reakcijama specifičnost može biti: Apsolutna (samo na jedno jedinjenje): npr ureaza na ureu, arginaza na arginin Relativna ili grupna (na grupu jedinjenja slične strukture): npr. lipaze hidrolizuju estarsku vezu u lipidima između svih alkohola i masnih kiselina, a najefikasniji su ako se radi o estrima glicerola i masne kiseline.
: npr ureaza na ureu, arginaza na arginin. Relativna ili grupna (na grupu jedinjenja slične strukture): npr. lipaze hidrolizuju estarsku vezu u lipidima između svih alkohola i masnih kiselina, a najefikasniji su ako se radi o estrima glicerola i masne kiseline.")
61
model “ključ-brava” enzim ima premodifikovano mesto vezivanja za supstrat koje tačno odgovara strukturi supstrata

62
model “indukovanog uklapanja”
enzim nema premodifikovano mesto vezivanja za supstrat tek u prisustvu supstrata dolazi do uklapanja

63
MEHANIZAM ENZIMSKE REAKCIJE:
Enzimi ne VRŠE reakcije, oni ih UBRZAVAJU E + S ES P + E tumačenje formule: ENZIM se vezuje za SUPSTRAT (jedinjenje/a koja resguju) enzim i supstrat stupaju u prelazni kompleks ili tranziciono stanje (odigrava se reakcija) rezultat reakcije je PRODUKT, a ENZIM izlazi NEIZMENJEN!!!!!
 enzim i supstrat stupaju u prelazni kompleks ili tranziciono stanje (odigrava se reakcija) rezultat reakcije je PRODUKT, a ENZIM izlazi NEIZMENJEN!!!!!")
64
S P ES G D G D G Shematski dijagram efekta enzima na hemijsku reakciju
TRANZICIONO STANJE G # D G # D G S P Tok reakcije G slobodna energija S polazni supstrat P produkt reakcije G slobodna energija aktivacije ( ) nekatalizovana reakcija; ( ••••••• ) enzimom katalizovana reakcija.
 nekatalizovana reakcija; ( ••••••• ) enzimom katalizovana reakcija.")
65
Brzina enzimske reakcije
Hemijske reakcije u ćeliji moraju se odigravati velikom brzinom kako bi se proizvodi tih reakcija javljali u dovoljnoj koncentraciji da bi se zadovoljile potrebe organizma. Brzina enzimskih reakcija = promena koncentracije supstrata u jedinici vremena ili promena koncentracije proizvoda reakcije u jedinici vremena.

66
Brzina enzimskih reakcija zavisi od:
koncentracije enzima koncentracije supstrata temperature pH sredine.

67
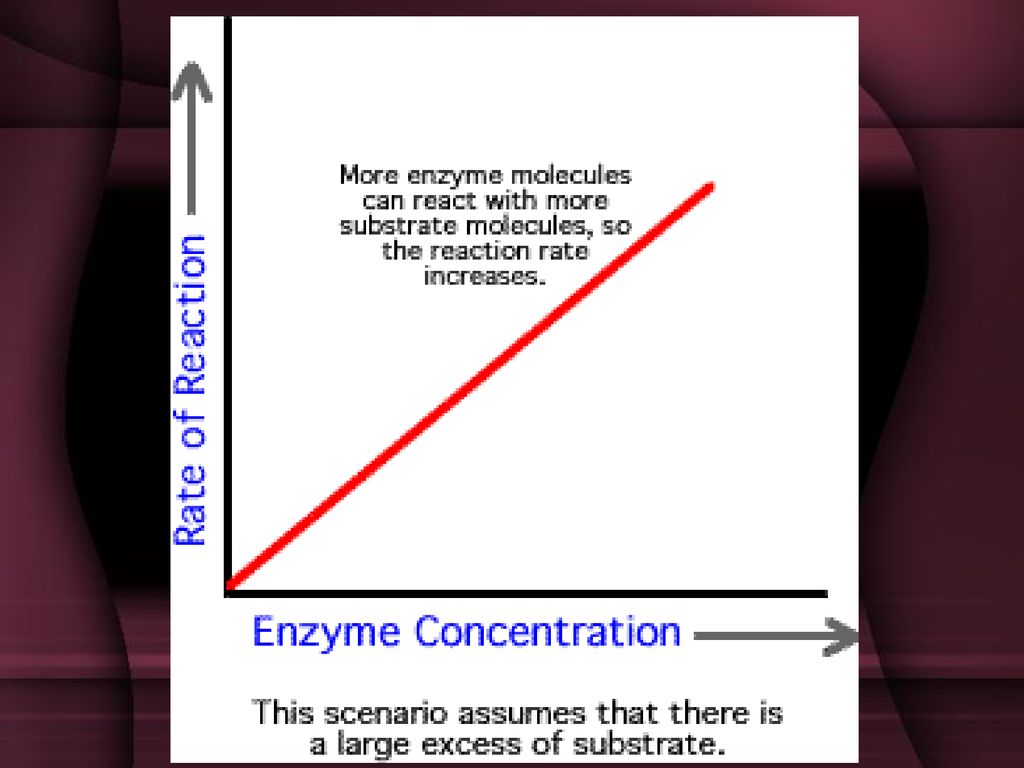
Uticaj koncentracije enzima
Enzimi su veoma efikasni katalizatori male količine enzima tranformišu velike količine supstrata u jedinici vremena (npr. 1g labfermenta može da zgruša 5 000l mleka, 1g ureaze može da razgradi 10000g uree). Što je koncentracija enzima veća – veća je i brzina enzimske reakcije
. Što je koncentracija enzima veća – veća je i brzina enzimske reakcije.")
69
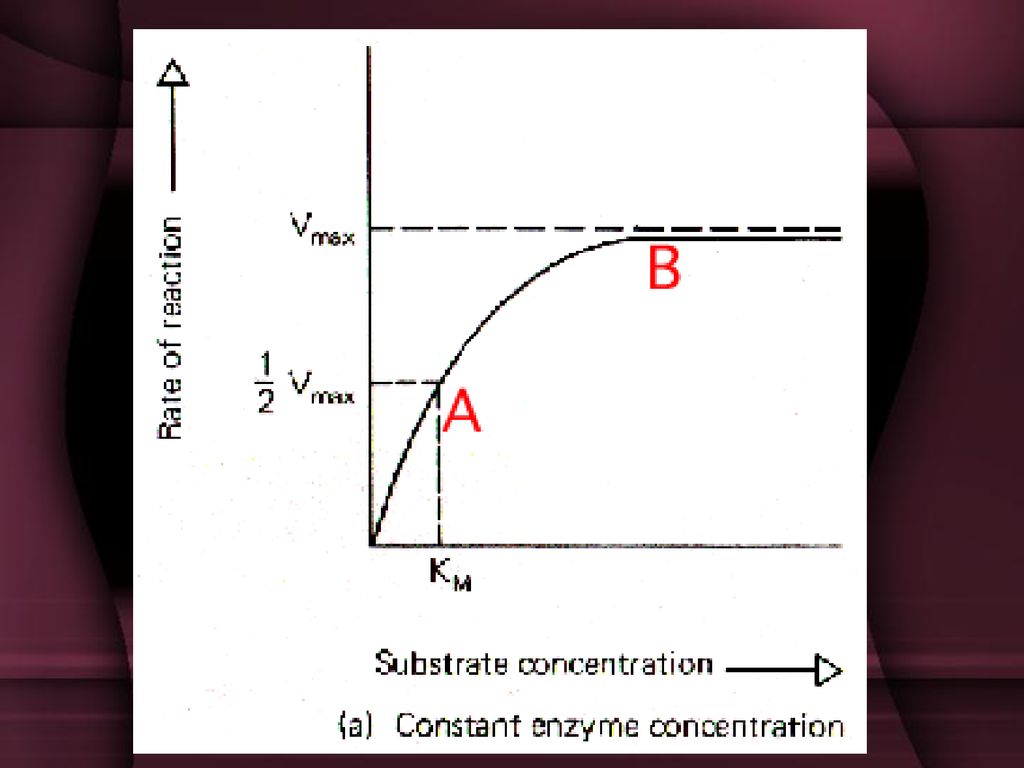
uticaj koncentracije supstrata
Brzina enzimskih reakcija zavisi od koncentracije kompleksa enzim-supstrat jedno vreme sa koncentracijom supstrata raste brzina Kada koncentracija supstrata prevaziđe koncentraciju enzima, svi molekuli enzima su u kompleksu sa supstratom i zato povećanje koncentracije supstrata ne povećava brzinu.

71
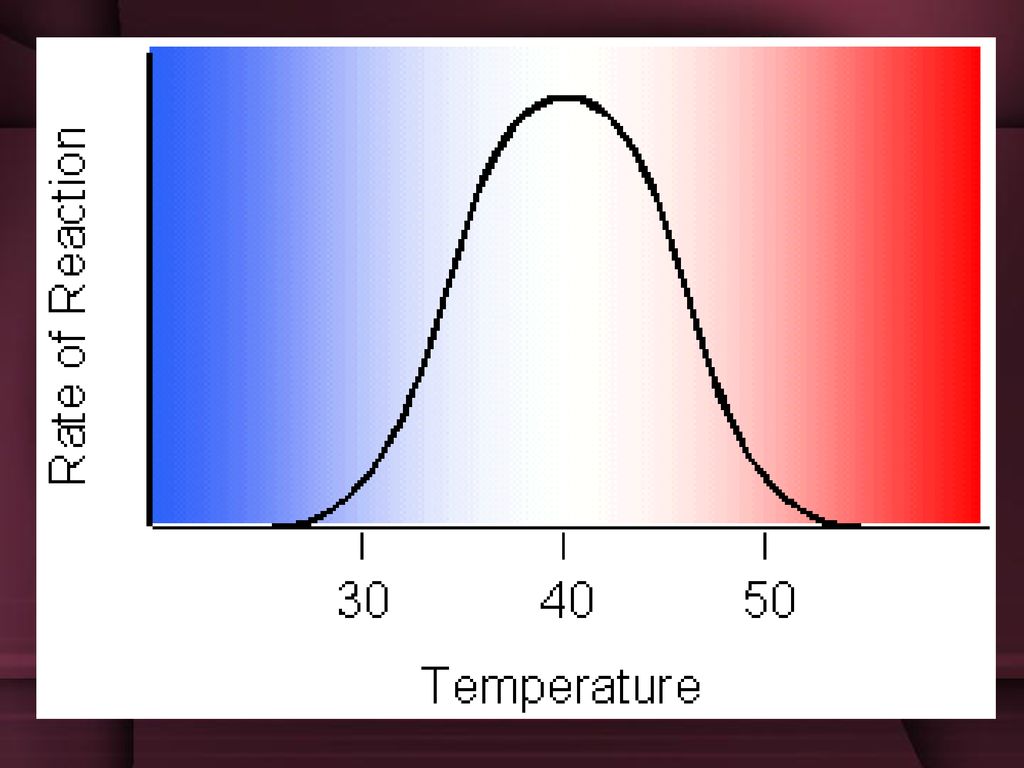
uticaj temperature Za sve enzime postoji optimalna temperatura na kojoj enzim transformiše najveću količinu supstrata i ta temperatura je 37C. Ako se temperatura smanji ispod optimalne, brzina se smanji. Ako se temperatura poveća iznad optimalne, brzina se takođe smanjuje. Na temperaturi od 42 C prestaje enzimska aktivnost jer se enzimi denaturišu toplotom.

73
uticaj pH Dok je optimalna temperatura istovetna za sve enzime, optimalni pH je različit za razne enzime. Npr. enzim amilaza koji deluje u ustima ima optimalni pH 7 (neutralna sredina) pepsin u želudcu pH 1,5 do 2,5 (kisela sredina) tripsin i himotripsin u tankom crevu pH oko 11 (jako bazna sredina)
 pepsin u želudcu pH 1,5 do 2,5 (kisela sredina) tripsin i himotripsin u tankom crevu pH oko 11 (jako bazna sredina)")
75
Inhibitori enzima To su mnoge supstance koje ometaju ili potpuno koče aktivnost enzima i na taj način sprečavaju odvijanje nekih hemijskih reakcija u organizmu. Jedni se mogu vezivati za enzim na istom mestu za koje se vezuje i supstrat, i tako sprečavaju vezivanje supstrata i enzima, te koče enzimsku reakciju. Drugi se ne moraju vezivati za isto mesto već za neko drugo mesto, ali kada se inhibitor veže za enzim , on toliko izmeni strukturu enzima da je nemoguće vezivanje enzima i supstrata. Treći su otrovi za živa bića, npr. enzimi koji u sebi sadrže Fe mogu se inhibirati cijanidima, Co - fluoridima, oksalatima, živom, i arsenom.

76
Ćelijska membrana

77
Ćelijska membrana Ćelijska membrana izgrađena je od lipida i proteina, kojima su pridruženi polisaharidi. Lipidi i proteini su međusobno u takvom odnosu da grade tzv. tečni mozaik Neki proteini su potpuno uronjeni u dvosloj lipida i izviruju i na spoljašnjoj i na unutrašnjoj strani membrane, dok se drugi nalaze ili na spoljašnjoj ili na unutrašnjoj strani membrane (vidi sliku). polisaharidi se vezuju za lipide i proteine ali samo na spoljašnjoj strani ćelijske membrane.
. polisaharidi se vezuju za lipide i proteine ali samo na spoljašnjoj strani ćelijske membrane.")
78
Lipidi membrane Lipidi u membrani eukariotskih ćelija mogu biti:
fosfolipidi, glikolipidi i holesterol. Lipidi membrane su polarizovani (razlikuju se dva kraja – pola) jedan kraj je hidrofilan i naziva se glava, a drugi je hidrofoban i nazvan je rep. Zahvaljujući tome oni formiraju dva sloja u membrani tako što se repovi grupišu u sredini, a glave se okreću prema spolja . fosfolipidi
 jedan kraj je hidrofilan i naziva se glava, a drugi je hidrofoban i nazvan je rep. Zahvaljujući tome oni formiraju dva sloja u membrani tako što se repovi grupišu u sredini, a glave se okreću prema spolja . fosfolipidi.")
79
Proteini membrane Proteini membrane se prema ulozi dele na:
strukturne koji uspostavljaju organizaciju ćelije tako što se vezuju za ćelijski skelet ili okoloćelijsku sredinu (ili oboje), kao i obrazovanje ćelijskih veza enzime receptore nosače i kanale, učestvuju u transportu kroz membranu kao npr. jonske pumpe Prema mestu koje zauzimaju u membrani proteini se mogu podeliti na: periferne koji se nalaze sa obe strane membrane (citoplazmatične i spoljašnje strane); integralne (nazivaju se još i transmembranski proteini) koji se delimično ili potpuno prostiru kroz membranu;
, kao i obrazovanje ćelijskih veza. enzime. receptore. nosače i kanale, učestvuju u transportu kroz membranu kao npr. jonske pumpe. Prema mestu koje zauzimaju u membrani proteini se mogu podeliti na: periferne koji se nalaze sa obe strane membrane (citoplazmatične i spoljašnje strane); integralne (nazivaju se još i transmembranski proteini) koji se delimično ili potpuno prostiru kroz membranu;")
80
Transport kroz ćelijsku membranu
Materije se kroz ćelijsku membranu mogu prenositi: pasivnim transportom – difuzijom nasumično kretanje čestica koje je uslovljeno njihovom kinetičkom energijom, kretanje se odvija u pravcu koncentracijskog gradijenta. aktivnim transportom Aktivni transport se odvija nasuprot koncentracijskog gradijenta, tako da je za ovaj transport potrebna hemijska energija i nosači.

81
Pasivni transport - difuzija
Difuzija kroz ćelijsku membranu se može odvijati: rastvaranjem u lipidima kroz pore membrane

82
Difuzija rastvaranjem u lipidima
Tako prolaze CO2, O2, masne kiseline i alkoholi. Kada ove materije dođu u dodir sa lipidnim dvoslojem membrane, odmah se rastvaraju, a zatim difunduju kroz lipidni dvosloj. faktor koji određuje brzinu difuzije kroz lipidni dvosloj membrane je rastvorljivost materije u lipidima. npr. O2 se lako rastvara u lipidima i on zato kroz lipidni dvosloj membrane prolazi nekoliko puta brže nego kroz vodu.

83
Difuzija kroz pore membrane
difunduju molekuli čiji je promer manji od promera pora i joni: molekuli H2O, uree i razni joni Pore membrane predstavljaju pukotine u membrani koje su oivičene proteinima. One su put za kretanje materija rastvorljivih u vodi. Proteini pore su pozitivno naelektrisani

84
Faktori koji određuju brzinu difuzije su veličina i naelektrisanje čestice
molekuli vode su manji od molekula uree, oni brže difunduju. Katjoni teže prolaze zbog svog pozitivnog naelektrisanja koje se odbija sa pozitivnim naelektrisanjem proteina pore. Katjoni sa manje pozitivnog naelektrisanja lakše prolaze (Na+ i K+ lakše prolaze od Mg2+ i Ca2+, a oni brže od feri jona: Fe3+). Anjoni brže prolaze od katjona jer ih privlači pozitivno naelektrisanje proteina pore.
. Anjoni brže prolaze od katjona jer ih privlači pozitivno naelektrisanje proteina pore.")
85
Olakšana difuzija Ovim procesom kroz membranu prolaze neki šećeri, pre svega glukoza i materije koje se slabo rastvaraju u lipidima. Da bi se materija kroz membranu prenosila na taj način, mora postojati koncentracijski gradijent i nosač u membrani za tu materiju.

86
veličina jona (stepen hidratacije)
Faktori od kojih zavisi dinamika olakšane difuzije: Količina materije koja se prenosi olakšanom difuzijom zavisi od koncentracijskog gradijenta za tu materiju i količine raspoloživog nosača. hormon insulin prenos glukoze ubrzava 7-10 puta, mada se ne zna kako insulin deluje: da li ubrzava vezivanje nosača i glukoze ili povećava količinu nosača u membrani. veličina jona (stepen hidratacije) Npr. hidrirani jon Na+ je za 30% veći od hidriranog jona K+, pa zato oko 100 puta teže difunduje. manjak Ca u vanćelijskoj tečnosti povećava propustljivost pora. hormon ADH (antidiuretični hormon) povećava promer pora.
 Npr. hidrirani jon Na+ je za 30% veći od hidriranog jona K+, pa zato oko 100 puta teže difunduje. manjak Ca u vanćelijskoj tečnosti povećava propustljivost pora. hormon ADH (antidiuretični hormon) povećava promer pora.")
87
Aktivni transport Aktivni transport je transport kroz ćelijsku membranu nasuprot koncentracijskog gradijenta zahteva energiju. Kroz ćelijsku membranu se aktivnim transportom razmenjuju: Na, K, Ca, Fe, H, neki šećeri i aminokiseline.

88
Opšti mehanizam aktivnog transporta:
neka materija T iz vanćelijske tečnosti treba da se ubaci u ćeliju, koncentracija T u ćeliji mnogo je veća nego u vanćelijskoj tečnosti, Materija T se na spoljašnjoj strani membrane vezuje za nosač N nastaje kompleks NT zatim se na unutrašnjoj strani membrane materija otpusti u ćeliju nosač se vrati ka spoljašnjoj strani membrane.

89
Zavisnost aktivnog transporta od energije
Ovom transportnom sistemu treba dodati energiju, jer se vrši nasuprot koncentracijskog gradijenta. Bez obzira na način korišćenja energije, ona se transportnom sistemu mora dodati na unutrašnjoj strani membrane, jer se tu nalazi ATP koji daje energiju za transport. Da bi ATP dao energiju, mora postojati enzim ATPaza koji razgrađuje ATP. Nosači membrane su u isto vreme i enzimi koji razgrađuju ATP. Nosači su ili proteini membrane ili lipoproteini. Protein uvek sadrži receptorsko mesto za materiju koja se prenosi aktivnim transportom

90
Aktivni transport Na+ i K+ “pumpa za Na+ i K+”
Na+ je ekstracelularni jon i njegova koncentracija u vanćelijskoj tečnosti je jako visoka. Na+ se aktivnim transportom stalno izbacuje u vanćelijsku tečnost. K+ je intracelularni jon i njegova koncentracija u ćeliji je mnogo veća nego u vanćelijskoj tečnosti. K+ se stalno iz vanćelijske tečnosti ubacuje u ćeliju.

91
Mehanizam Na+ / K+ pumpe:
na unutrašnjoj strani membrane se Na+ vezuje za nosač Y i nastaje kompleks Na+ Y. Nosač Y ima tri domena i tri mesta za vezivanje Na+ kompleks Na+ Y se okreće ka spoljašnjoj strani membrane gde se Na+ otpušta u vanćelijsku tečnost, nosač Y delimično menja svoju konformaciju i postaje nosač X. X na spoljašnjoj strani membrane veže K+, nastaje kompleks K+X koji se kreće ka unutrašnjoj strani membrane. ATP daje energiju za otpuštanje K+ u ćeliju, pri čemu će enzim ATP-aza izgrađivati ATP, a za ovaj proces neophodni su i joni Mg2+. Za 3 jona Na prenese se 1 jon K. Nosač je lipoprotein koji igra ulogu ATP-aze

94
Mehanizam prenosa Na i K važan je u mnogim ćelijama i funkcionalnim sistemima organizma.
Poseban značaj ima u nervnim i mišićnim ćelijama u prenosu nervnog impulsa. U drugim ćelijama aktivni transport Na i K sprečava bubrenje ćelije jer se zajedno sa Na iz ćelije izbacuje i voda. Aktivni transport Na i K označen je kao K-Na pumpa.

95
Aktivni transport glukoze
Kroz neke membrane aktivnim transportom se prenosi i glukoza: kroz epitel tankog creva i epitel bubrežnih tubula. Iz lumena creva, glukoza se aktivnim transportom prebacuje u krv, gde je njena koncentracija mnogo veća. Zbog takvog aktivnog transporta, glukoza se ne izbacuje fecesom i mokraćom.

96
KOTRANSPORT aktivni transport glukoze kroz neke membrane, on zavisi od akivnog transporta Na (Na/K- pumpe) jer taj proces daje energiju za aktivni transport glukoze. Smatra se da nosač koji aktivno prenosi Na ima još jedno receptorsko mesto za glukozu. Nosač neće prenositi glukozu ako se za njega ne veže Na.
 jer taj proces daje energiju za aktivni transport glukoze. Smatra se da nosač koji aktivno prenosi Na ima još jedno receptorsko mesto za glukozu. Nosač neće prenositi glukozu ako se za njega ne veže Na.")
97
Aktivni transport aminokiselina
Aminokiseline se takođe aktivnim transportom prenose kroz neke organe, epitel creva i bubrežnih tubula. Aktivni transport aminokiselina takođe zavisi od aktivnog transporta Na (tj. od energije tog sistema). Zavisi i od nekih hormona kao što su: hormon rasta, insulin, glikokortikoidi (oni ubrzavaju aktivni transport aminokiselina u ćeliji).
. Zavisi i od nekih hormona kao što su: hormon rasta, insulin, glikokortikoidi (oni ubrzavaju aktivni transport aminokiselina u ćeliji).")
98
Endocitoza To je oblik ulaska velikih molekula i krupnih čestica u ćeliju. Postoje 2 oblika endocitoze: pinocitoza fagocitoza

99
Pinocitoza ćelija iz vanćelijske tečnosti uzima krupne molekule, uglavnom proteine. Molekuli proteina koji su u vanćelijskoj tečnosti se apsorbuju na membranu, zatim se membrana na tom delu gde su se apsorbovali proteini ugiba i taj ugnuti deo membrane se odvaja u vidu pinocitoznog mehurića. Pinocitozni mehurić se zatim spaja sa lizozomom. Lizozom sadrži enzime koji razgrađuju proteine do aminokiselina; zatim aminokiseline pređu u citoplazmu gde se dalje koriste.

100
Fagocitoza To je mehanizam kojim ćelija iz vanćelijske tečnosti uzima krupne čestice: bakterije drugih ćelija ili čestice oštećenog tkiva. Mehanizam fagocitoze je sličan mehanizmu pinocitoze. Čestice koje se fagocitozom uzimaju apsorbuju se na površini membrane, koja se tu ugiba i ugnuti deo se odvaja u vidu fagocitoznog mehurića. Fagocitozni mehurić se spaja sa lizozomom koji sadrži enzim za razgradnju čestica fagocitoznog mehurića

101
Egzocitoza To je proces suprotan endocitozi.
Karakterističan je za sekretorne ćelije koje izlučuju određene materije: ćelije endokrinih i egzokrinih žlezda. Egzocitoza uključuje granulirani endoplazmatični retikulum (GER) i goldži-aparat. Ribozomi GER-a sintetišu proteine koji trebaju da budu sekretovani (izlučeni) iz ćelije. Ti proteini onda dospevaju u kanaliće endoplazmatičnog retikuluma i putem njih do goldži-aparata. U goldži-aparatu se ti proteini okruže membranom i u vidu sekretorne granule se odvoje od goldži-aparata. Sekretorna granula citoplazmom putuje do ćelijske membrane, gde se membrana granule spaja sa ćelijskom membranom, a proteini se izluče u vanćelijsku tečnost.
 i goldži-aparat. Ribozomi GER-a sintetišu proteine koji trebaju da budu sekretovani (izlučeni) iz ćelije. Ti proteini onda dospevaju u kanaliće endoplazmatičnog retikuluma i putem njih do goldži-aparata. U goldži-aparatu se ti proteini okruže membranom i u vidu sekretorne granule se odvoje od goldži-aparata. Sekretorna granula citoplazmom putuje do ćelijske membrane, gde se membrana granule spaja sa ćelijskom membranom, a proteini se izluče u vanćelijsku tečnost.")






>")

















