Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Προκαρυωτική Μεταγραφή ΙΙΙ: Trp operon – sporulation – phage λ

2
To οπερόνιο της τρυπτοφάνης στην E
To οπερόνιο της τρυπτοφάνης στην E. coli ρυθμίζεται με το μηχανισμό της εξασθένισης

3
Εναλλακτικές δευτεροταγείς δομές ελέγχουν την εξασθένηση
Οι εναλλακτικές δευτεροταγείς δομές που μπορεί να υιοθετήσει το RNA παρέχουν δυνατότητες για έλεγχο της έκφρασης. Αρκετά οπερόνια ρυθμίζονται με το μηχανισμό της εξασθένισης (attenuation). Η εξασθένιση είναι δυνατόν να ρυθμίζεται από πρωτεΐνες που προσδένονται στο RNA και σταθεροποιούν ή αποσταθεροποιούν το σχηματισμό της φουρκέτας που απαιτείται για τον τερματισμό.
. Η εξασθένιση είναι δυνατόν να ρυθμίζεται από πρωτεΐνες που προσδένονται στο RNA και σταθεροποιούν ή αποσταθεροποιούν το σχηματισμό της φουρκέτας που απαιτείται για τον τερματισμό.")
4
Η χρωμοσωμική σειρά των γονιδίων του οπερονίου trp της E
Η χρωμοσωμική σειρά των γονιδίων του οπερονίου trp της E. coli και η αλληλουχία των αντιδράσεων που καταλύονται από τα ενζυμικά προϊόντα των δομικών γονιδίων του οπερονίου

5
Αρνητικός έλεγχος του οπερονίου trp

6
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα O τερματισμός της μεταγραφής μπορεί να ελεγχθεί μέσω αλλαγών στη δευτεροταγή δομή του RNA. Οι αλλαγές αυτές ρυθμίζονται από την κίνηση του ριβοσώματος. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

7
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Tο οπερόνιο trp αποτελείται από πέντε συνεχή δομικά γονίδια και μία ρυθμιστική περιοχή που προηγείται και περιλαμβάνει τον υποκινητή, το χειριστή, μία περιοχή που κωδικοποιεί ένα πεπτίδιο-οδηγό και έναν εξασθενητή.

8
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Ένας εξασθενητής ρυθμίζει την προώθηση της RNA πολυμεράσης στα γονίδια του οπερονίου trp. Η RNA πολυμεράση ξεκινά τη μεταγραφή στον υποκινητή και προχωρά μέχρι τη θέση 90, όπου σταματά, πριν μετακινηθεί προς τον εξασθενητή που βρίσκεται στη θέση 140. Απουσία τρυπτοφάνης, η πολυμεράση προχωρά στα δομικά γονίδια (το trpΕ ξεκινά στο +163). Παρουσία τρυπτοφάνης, υπάρχει μεγάλη πιθανότητα τερματισμού (~90%), που ακολουθείται από την απελευθέρωση του, μήκους 140 βάσεων, RNA-οδηγού. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
. Παρουσία τρυπτοφάνης, υπάρχει μεγάλη πιθανότητα τερματισμού (~90%), που ακολουθείται από την απελευθέρωση του, μήκους 140 βάσεων, RNA-οδηγού. Genes VIII - Ακαδημαϊκές Εκδόσεις")
9
Όταν υπάρχει τρυπτοφάνη ο τερματισμός είναι αποτελεσματικός και ο εξασθενητής επιτρέπει μόνο το 10% των μορίων της RNA pol να περάσει. Απουσία τρυπτοφάνης πρακτικά όλα τα μόρια της RNA pol προχωρούν. Άρση εξασθένισης → ~10x επίδραση Άρση καταστολής → ~70x αύξηση ρυθμού έναρξης μεταγραφής => συνολικό εύρος ρύθμισης ~700x.

10
H εξασθένιση είναι δυνατόν να ρυθμιστεί από τη μετάφραση

11
Οι δυο πιθανές δομές της αλληλουχίας-οδηγού

12
Η αλληλουχία του οδηγού

13
Μοντέλα μηχανισμού εξασθένισης

14
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Παρουσία φορτισμένου tRNA της τρυπτοφάνης, τα ριβοσώματα μεταφράζουν το πεπτίδιο-οδηγό και μετά απελευθερώνονται. Αυτή η διαδικασία επιτρέπει το σχηματισμό της φουρκέτας τερματισμού και έτσι η RNA πολυμεράση τερματίζει. Απουσία φορτισμένου tRNA της τρυπτοφάνης, το ριβόσωμα σταματά, η φουρκέτα τερματισμού δε σχηματίζεται και η RNA πολυμεράση συνεχίζει τη μεταγραφή. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

15
Η σποριογονία ελέγχεται από παράγοντες σ

16
Τα δύο είδη κυττάρων του B. subtilis
Βλαστικά κύτταρα (Vegetative cells) Δημιουργία ενδοσπορίου (στο αριστερό άκρο του κυττάρου) κατά τη σποριογένεση
 Δημιουργία ενδοσπορίου (στο αριστερό άκρο του κυττάρου) κατά τη σποριογένεση.")
17
Η σποριογονία ενέχει τη διαφοροποίηση ενός ασεξουαλικώς αναπτυσσόμενου βακτηρίου σε ένα μητρικό κύτταρο που λύεται και σε ένα σπόριο που απελευθερώνεται.

18
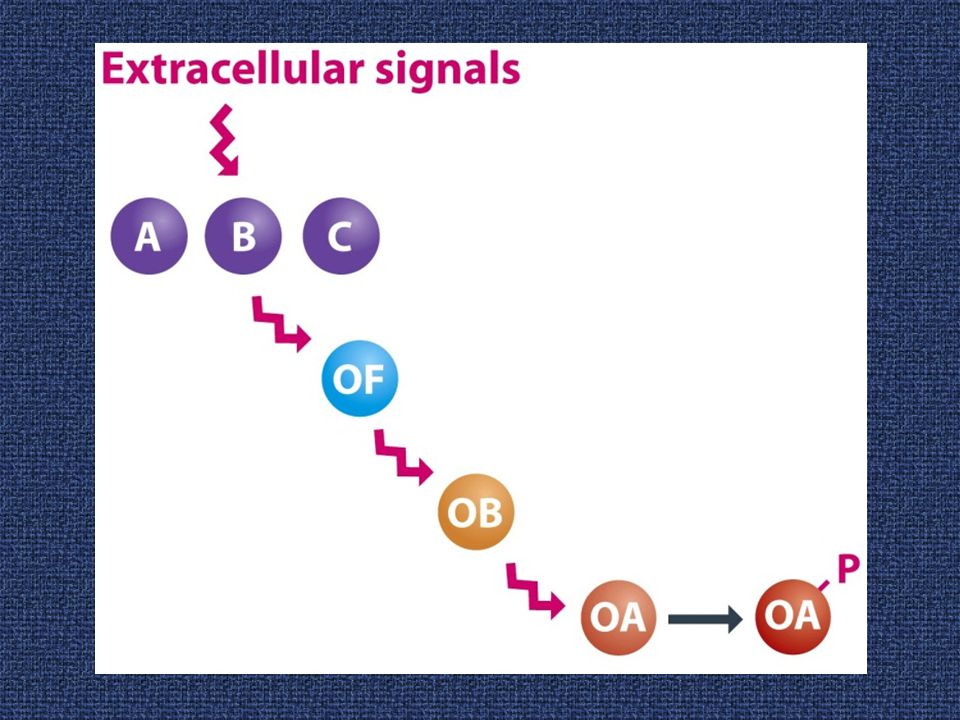
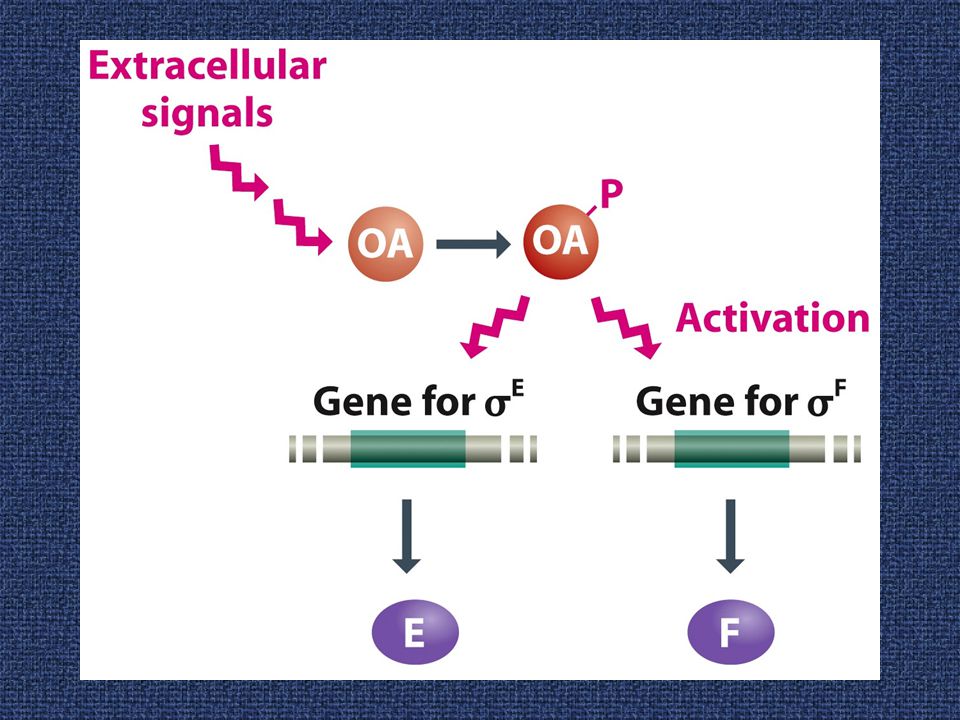
Ειδικές υπομονάδες σ ελέγχουν τη δραστηριότητα του γονιδιώματος κατά τη σποριογένεση

20
Φωσφο-ρυθμιστικά κυκλώματα στη σποριογένεση του B. subtilis

22
Οι εμπλεκόμενοι μηχανισμοί στη ρύθμιση των παραγόντων σ κατά τη διαμερισματο-ποίηση στη σποριογένεση
Η σποριογονία περιλαμβάνει διαδοχικές αλλαγές των παραγόντων σίγμα, οι οποίοι ελέγχουν την εξειδίκευση της RNA πολυμεράσης στο επίπεδο την έναρξης της μεταγραφής. Οι καταρράκτες στο προσπόριο (δεξιά) και στο μητρικό κύτταρο (αριστερά) συντονίζονται με σήματα που διέρχονται μέσω του διαφράγματος.
 και στο μητρικό κύτταρο (αριστερά) συντονίζονται με σήματα που διέρχονται μέσω του διαφράγματος.")
23
Η σύνθεση των πρωτεϊνών r ρυθμίζεται με αυτογενή έλεγχο

24
Εικόνα 11.17 Τα γονίδια που κωδικοποιούν τις ριβοσωμικές πρωτεΐνες, τους παράγοντες της πρωτεϊνοσύνθεσης και τις υπομονάδες της RNA πολυμεράσης (~70) απαντώνται διάσπαρτα σε λίγα οπερόνια που ρυθμίζονται αυτόνομα. O ρυθμιστής αναγράφεται με κόκκινο και οι πρωτεΐνες που ρυθμίζονται είναι σκιασμένες με ροζ.
 απαντώνται διάσπαρτα σε λίγα οπερόνια που ρυθμίζονται αυτόνομα. O ρυθμιστής αναγράφεται με κόκκινο και οι πρωτεΐνες που ρυθμίζονται είναι σκιασμένες με ροζ.")
25
Σχεδόν όλες οι ριβοσωμικές πρωτεΐνες απαιτούνται σε ισομοριακές ποσότητες που πρέπει να ρυθμιστούν σε σχέση με τα επίπεδα του rRNA Κοινό χαρακτηριστικό όλων των οπερονίων (που κωδικοποιούν τις ριβοσωμικές πρωτεΐνες) είναι ότι ένα από τα προϊόντα τους ρυθμίζει μερικά από τα γονίδιά τους Το γονίδιο που κωδικοποιεί το ρυθμιστικό προϊόν αποτελεί ταυτόχρονα και έναν από τους στόχους του = αυτογενής έλεγχος
 είναι ότι ένα από τα προϊόντα τους ρυθμίζει μερικά από τα γονίδιά τους. Το γονίδιο που κωδικοποιεί το ρυθμιστικό προϊόν αποτελεί ταυτόχρονα και έναν από τους στόχους του = αυτογενής έλεγχος.")
26
Σε όλες τις περιπτώσεις, η συσσώρευση της πρωτεΐνης αναστέλλει τη σύνθεσή της
Κάθε ένας από τους ρυθμιστές είναι μια ριβοσωμική πρωτεΐνη που προσδένεται άμεσα στο rRNA, Eπιπλέον, όμως, διαθέτει και την ιδιότητα να προσδένεται στο δικό της mRNA

27
Πώς εξασφαλίζεται ότι οι r πρωτεΐνες συντίθενται σε ποσότητες αντίστοιχες με αυτές των rRNA;
Εικόνα H μετάφραση των οπερονίων των πρωτεϊνών r ρυθμίζεται με αυτογενή έλεγχο και αποκρίνεται στα επίπεδα του rRNA.

28
ΣΤΡΑΤΗΓΙΚΕΣ ΦΑΓΩΝ

29
Εικόνα 12.1 H λυτική ανάπτυξη περιλαμβάνει την αναπαραγωγή των σωματίων του φάγου και την ακόλουθη καταστροφή του βακτηρίου-ξενιστή. Η λυσιγονία επιτρέπει στο γονιδίωμα του φάγου να μεταφερθεί ως τμήμα της γενετικής πληροφορίας του βακτηρίου.

31
Η λυτική ανάπτυξη ρυθμίζεται από έναν καταρράκτη γεγονότων

32
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 12.4 H λυτική ανάπτυξη προχωρά μέσω ενός ρυθμιστικού καταρράκτη, κατά τον οποίο ένα από τα γονιδιακά προϊόντα κάθε σταδίου απαιτείται για την έκφραση των γονιδίων του επόμενου σταδίου. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

33
Υπάρχουν δύο μηχανισμοί ρύθμισης του λυτικού καταρράκτη
Οι ρυθμιστικές πρωτεΐνες που χρησιμοποιούνται στους καταρράκτες των φάγων είτε: προωθούν την έναρξη της μεταγραφής σε νέους υποκινητές του φάγου επιτρέπουν στην πολυμεράση του ξενιστή να προσπεράσει αλληλουχίες τερματισμού

34
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 12.5 Όταν ο έλεγχος γίνεται στο επίπεδο της έναρξης της μεταγραφής, τα εμπλεκόμενα γονίδια συνιστούν ανεξάρτητες μεταγραφικές μονάδες που, διαθέτοντας η καθεμία το δικό της υποκινητή και τη δική της αλληλουχία τερματισμού, παράγουν ανεξάρτητα μόρια mRNA. Στην περίπτωση αυτή, δε χρειάζεται οι μεταγραφικές μονάδες να βρίσκονται κοντά. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

35
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 12.6 Όταν ο έλεγχος γίνεται στο επίπεδο του τερματισμού της μεταγραφής, τα εμπλεκόμενα γονίδια πρέπει να είναι παρακείμενα, έτσι ώστε να είναι δυνατόν να προχωρήσει η μεταγραφή από την πρώτη ομάδα γονιδίων στην επόμενη. Με τον τρόπο αυτό, και οι δύο ομάδες γονιδίων μεταγράφονται στο ίδιο μόριο mRNA. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

36
Τα αμέσως πρώιμα και τα καθυστερημένα πρώιμα γονίδια του φάγου λ απαιτούνται τόσο για τη λυσιγονία όσο και για το λυτικό κύκλο

37
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα O λυτικός καταρράκτης του φάγου λ διαπλέκεται με το λυσιγονικό κύκλωμα. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

38
Εικόνα Στο χάρτη του φάγου λ παρατηρείται ομαδοποίηση των σχετιζόμενων λειτουργιών. Το γονιδίωμα του φάγου λ έχει μήκος 48,514 bp.

39
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα O φάγος λ διαθέτει δύο πρώιμες μεταγραφικές μονάδες. Στην «αριστερή» μονάδα μεταγράφεται η «επάνω» αλυσίδα προς τα αριστερά, ενώ στη «δεξιά» μονάδα μεταγράφεται η «κάτω» αλυσίδα προς τα δεξιά. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

40
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Το DNA του φάγου λ κυκλοποιείται κατά τη μόλυνση, με αποτέλεσμα η όψιμη συστοιχία γονιδίων να σχηματίζει μια ενιαία μεταγραφική μονάδα. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

41
Ο αντιτερματισμός αποτελεί ρυθμιστικό μηχανισμό γονιδιακής έκφρασης

42
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.52 Μια πρωτεΐνη αντιτερματισμού μπορεί να επιδράσει στην RNA πολυμεράση επιτρέποντας την αναγνωστική διέλευσή της από μια ειδική αλληλουχία τερματισμού. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

43
Εικόνα 9.53 Η RNA πολυμεράση του ξενιστή μεταγράφει τα γονίδια του φάγου λ και τερματίζει τη μεταγραφή στις θέσεις t. Η πρωτεΐνη pN επιτρέπει την αναγνωστική διέλευση της RNA πολυμεράσης από τις αλληλουχίες τερματισμού tL και tR. Η πρωτεΐνη pQ επιτρέπει την αναγνωστική διέλευση της RNA πολυμεράσης από την αλληλουχία τερματισμού tR. Τα σημεία στα οποία δρα η pN (nut) και η pQ (qut) εντοπίζονται αντίστοιχα σε διαφορετικές σχετικές θέσεις στις μεταγραφικές μονάδες. Ο αντι-τερματισμός απαιτεί θέσεις που είναι ανεξάρτητες από τις αλληλουχίες τερματισμού
 και η pQ (qut) εντοπίζονται αντίστοιχα σε διαφορετικές σχετικές θέσεις στις μεταγραφικές μονάδες. Ο αντι-τερματισμός απαιτεί θέσεις που είναι ανεξάρτητες από τις αλληλουχίες τερματισμού.")
44
Πώς συμβαίνει ο αντιτερματισμός;
Εικόνα 9.54 Βοηθητικοί παράγοντες προσδένονται στην RNA πολυμεράση καθώς διέρχεται από τη θέση nut. Αυτοί εμποδίζουν τον Rho να προκαλέσει τερματισμό, όταν η πολυμεράση φτάσει στην αλληλουχία τερματισμού.

45
Οι παράγοντες τερματισμού και αντι-τερματισμού αλληλεπιδρούν με την RNA πολυμεράση
Μεταλλάξεις που καθιστούν την pN αναποτελεσματική ταυτοποιούν πρωτεΐνες που σχετίζονται με τον τερματισμό: Στο γονίδιο rpoB η pN αλληλεπιδρά με την υπομονάδα β του κεντρικού ενζύμου Στους γενετικούς τόπους nus (N utilization substance): nusA, nusB, nusE, nusG.
: nusA, nusB, nusE, nusG.")
46
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 9.55 Βοηθητικοί παράγοντες προσδένονται στην RNA πολυμεράση καθώς διέρχεται από συγκεκριμένες θέσεις. Η θέση nut αποτελείται από δύο αλληλουχίες. Ο παράγοντας NusB-S10 ενώνεται με το κεντρικό ένζυμο καθώς αυτό διέρχεται από το boxA. Στη συνέχεια, προσδένονται οι πρωτεΐνες NusA και pN καθώς η πολυμεράση διέρχεται από το boxB. Η παρουσία της pN επιτρέπει την αναγνωστική διέλευση του ενζύμου από την αλληλουχία τερματισμού. Έτσι, παράγεται ένα ενιαίο mRNA που περιέχει αμέσως πρώιμες αλληλουχίες ενωμένες με καθυστερημένες πρώιμες αλληλουχίες.

47
Άραγε η pN αναγνωρίζει την αλληλουχία boxB στο DNA ή στο mRNA;
H pN δεν μπορεί να συνδεθεί από μόνη της σε κανένα από τα δύο είδη αλληλουχιών Συνδέεται με το μεταγραφικό σύμπλοκο καθώς το κεντρικό ένζυμο περνάει από τη θέση boxB

48
Η στάση σε αλληλουχίες τερματισμού είναι προϋπόθεση για να συμβεί τερματισμός
Υπό την επίδραση παραγόντων όπως ο NusA, η στάση παρατείνεται, αυξάνοντας την αποτελεσματικότητα του τερματισμού Υπό την επίδραση των παραγόντων pN ή pQ η στάση γίνεται πιο σύντομη, μειώνοντας έτσι την αποτελεσματικότητα του τερματισμού

49
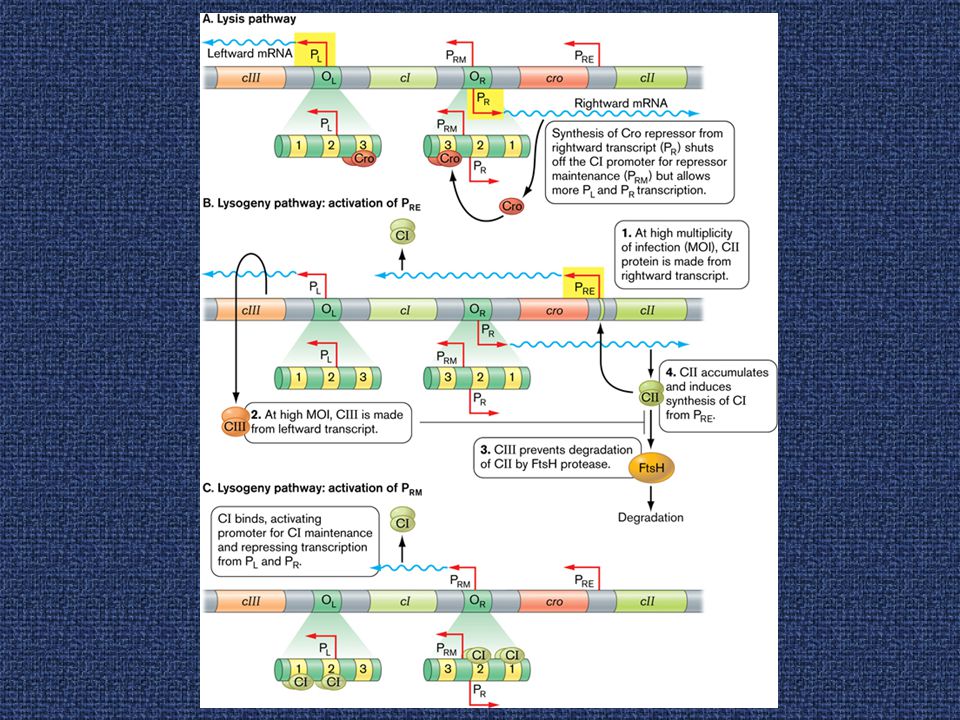
Η λυσιγονία διατηρείται μέσω μιας πρωτεΐνης-καταστολέα

50
Εικόνα H ρυθμιστική περιοχή του φάγου λ περιλαμβάνει cis-δραστικά στοιχεία και μια συστοιχία ρυθμιστικών γονιδίων. Φάγοι με μεταλλάξεις στο γονίδιο cI είναι ανίκανοι να υποστηρίξουν λυσιγονία προχωρούν πάντοτε στο λυτικό μονοπάτι. Ο cI μεταγράφεται από τον PRM. Επειδή απουσιάζει η συνήθης θέση πρόσδεσης του ριβοσώματος, ο ρυθμός μετάφρασης του cI είναι μειωμένος. Έτσι, τα επίπεδα του cI διατηρούνται χαμηλά.

51
Ο καταστολέας συντηρεί ένα αυτογενές κύκλωμα

52
Εικόνα 12. 16 Ένα αυτογενές κύκλωμα διατηρεί τη λυσιγονία (επάνω)
Εικόνα Ένα αυτογενές κύκλωμα διατηρεί τη λυσιγονία (επάνω). Aν το κύκλωμα αυτό διακοπεί, τότε ξεκινάει ο λυτικός κύκλος (κάτω). Η πρόσδεση του CI στον OL δεν επιτρέπει στην RNA pol να ξεκινήσει μεταγραφή από τον PL. Η πρόσδεση του CI στον OR παρεμποδίζει τη χρήση του PR. Η πρόσδεση του CI στον OR συνεισφέρει στην αποτελεσματική πρόσδεση της RNA pol στον PRM.
. Aν το κύκλωμα αυτό διακοπεί, τότε ξεκινάει ο λυτικός κύκλος (κάτω). Η πρόσδεση του CI στον OL δεν επιτρέπει στην RNA pol να ξεκινήσει μεταγραφή από τον PL. Η πρόσδεση του CI στον OR παρεμποδίζει τη χρήση του PR. Η πρόσδεση του CI στον OR συνεισφέρει στην αποτελεσματική πρόσδεση της RNA pol στον PRM.")
53
Αν το DNA ενός δεύτερου φάγου λ εισέλθει σε ένα κύτταρο που βρίσκεται ήδη σε λυσιγονία, η CI του πρώτου φάγου θα προσδεθεί αμέσως στους OR και OL του νέου γονιδιώματος. Αυτό θα εμποδίσει το δεύτερο φάγο να ξεκινήσει το λυτικό κύκλο Ανοσία Τι θα συμβεί αν ο δεύτερο φάγος έχει μεταλλαγμένους του OR και OL ;

54
Ο καταστολέας προσδένεται στο DNA ως διμερές

55
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα H N-τελική και η C-τελική περιοχή του καταστολέα συνιστούν διακριτές επικράτειες. Οι C-τελικές επικράτειες αλληλεπιδρούν για το σχηματισμό του διμερούς, ενώ με τις Ν-τελικές επικράτειες το διμερές προσδένεται στο DNA. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

56
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα O χειριστής αποτελείται από μια αλληλουχία 17 bp που είναι μερικώς συμμετρική ως προς έναν άξονα ο οποίος περνά από το κεντρικό ζεύγος βάσεων. Με πράσινη σκίαση σημειώνεται η κάθε ημιθέση. Με κόκκινο σημειώνονται οι βάσεις που είναι όμοιες σε κάθε ημιθέση του χειριστή. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

57
Τα διμερή του καταστολέα προσδένονται με συνέργεια στο χειριστή

58
Εικόνα Kάθε χειριστής περιλαμβάνει τρεις θέσεις πρόσδεσης του καταστολέα και επικαλύπτεται με τον υποκινητή στον οποίο προσδένεται η RNA πολυμεράση. O προσανατολισμός του OL έχει αντιστραφεί, ώστε να διευκολυνθεί η σύγκριση με τον OR.

59
Εικόνα Όταν δύο διμερή του καταστολέα του φάγου λ προσδένονται στο DNA με συνέργεια, καθεμία από τις υπομονάδες του ενός διμερούς αλληλεπιδρά με μία υπομονάδα του άλλου. Σε κάθε χειριστή, η θέση 1 εμφανίζει τη μεγαλύτερη συγγένεια πρόσδεσης, περίπου 10x των άλλων. Η παρουσία ενός διμερούς στη θέση 1 αυξάνει κατά πολύ τη συγγένεια με την οποία ένα δεύτερο διμερές μπορεί να προσδεθεί στη θέση 2. Όταν όμως οι θέσεις 1 και 2 είναι κατειλημμένες, η πρόσδεση στη θέση 3 δεν διευκολύνεται.

60
Ο καταστολέας στη θέση OR2 αλληλεπιδρά με την RNA pol στον υποκινητή PRM

61
Ο καταστολέας που έχει προσδεθεί στη θέση OR2 αλληλεπιδρά, μέσω της περιοχής πρόσδεσής του στο DNA, με την RNA pol και σταθεροποιεί την αλληλεπίδραση της πολυμεράσης με τον PRM. Η διαδικασία αυτή αποτελεί τη βάση για τον αυτογενή έλεγχο που εξασφαλίζει τη διατήρηση της σύνθεσης του καταστολέα.

62
Τι συμβαίνει αν ένα διμερές του καταστολέα προσδεθεί στη θέση OR3;
cI down → απελευθέρωση OR3 → cI up Ο μηχανισμός αυτός εμποδίζει τη συγκέντρωση του καταστολέα στο λυσιγονικό βακτήριο να φτάσει σε ασυνήθιστα υψηλά επίπεδα. Ο καταστολέας, λοιπόν, αποτελεί έναν αυτογενή ρυθμιστή της έκφρασής του, ο οποίος λειτουργεί θετικά σε χαμηλές συγκεντρώσεις και αρνητικά σε υψηλές

63
Τα γονίδια cII και cIII απαιτούνται για την εγκαθίδρυση της λυσιγονίας
Η παρουσία του καταστολέα είναι απαραίτητη για τη διατήρηση της σύνθεσής του από τον PRM. Πώς ξεκινά de novo η σύνθεση του καταστολέα;

64
Εικόνα Όταν το DNA ενός φάγου εισέλθει σε ένα νέο κύτταρο-ξενιστή, η RNA pol δεν μπορεί να μεταγράψει το cI, διότι δεν υπάρχει ο καταστολέας (CI) να υποβοηθήσει την πρόσδεση της πολυμεράσης στον PRM. H σύνθεση του καταστολέα εγκαθιδρύεται χάρη στη δράση της πρωτεΐνης cII και της RNA πολυμεράσης στον υποκινητή PRΕ, από τον οποίο μεταγράφεται η αντισημαίνουσα αλυσίδα του cro και στη συνέχεια το γονίδιο cI.

65
H cII υποβοηθά την RNA pol να αναγνωρίσει τον PRE.
H cIII εμποδίζει την αποικοδόμηση της cII από την HflA (του ξενιστή). παράγεται η cI. συντίθεται το αντισημαίνον cro μπλοκάροντας τη σύνθεση της Cro που χρειάζεται για την είσοδο στο λυτικό κύκλο. H cII διεγείρει επίσης τη μεταγραφή της ιντεγκράσης από τον PI. Η ιντεγκράση καταλύει την ενσωμάτωση του DNA του φάγου στον ξενιστή, βασική προϋπόθεση για τη λυσιγονία
. παράγεται η cI. συντίθεται το αντισημαίνον cro μπλοκάροντας τη σύνθεση της Cro που χρειάζεται για την είσοδο στο λυτικό κύκλο. H cII διεγείρει επίσης τη μεταγραφή της ιντεγκράσης από τον PI. Η ιντεγκράση καταλύει την ενσωμάτωση του DNA του φάγου στον ξενιστή, βασική προϋπόθεση για τη λυσιγονία.")
66
Ένας αδύναμος υποκινητής λειτουργεί με τη βοήθεια της cII

67
Εικόνα H RNA πολυμεράση προσδένεται στον υποκινητή PRΕ μόνο παρουσία της πρωτεΐνης cII, η οποία αλληλεπιδρά με τον υποκινητή αυτό γύρω από τη θέση -35.

68
H λυσιγονία εγκαθιδρύεται μέσω μιας διαδοχής γεγονότων

69
Εικόνα Για την εγκαθίδρυση της λυσιγονίας απαιτείται αρχικά ένας καταρράκτης. Στη συνέχεια, το κύκλωμα αυτό διακόπτεται και αντικαθίσταται από το αυτογενές κύκλωμα διατήρησης του καταστολέα.

70
Ο καταστολέας Cro απαιτείται για τη λυτική μόλυνση
Παρεμποδίζει τη σύνθεση του cI αναστέλλοντας τη μεταγραφή από τον PRM. Καταστέλλει την έκφραση των πρώιμων γονιδίων από τους PL και PR.

71
Η δομή και των δύο πρωτεϊνών είναι παρεμφερής.
H Cro επιτυγχάνει τη δράση της μέσω πρόσδεσης στους ίδιους χειριστές όπου προσδένεται και ο cI. Η δομή και των δύο πρωτεϊνών είναι παρεμφερής. Πώς γίνεται δύο πρωτεΐνες να αναγνωρίζουν τις ίδιες θέσεις στο DNA και να έχουν αντίθετα αποτελέσματα; Διαφορετική συγγένεια πρόσδεσης για τις ξεχωριστές θέσεις των χειριστών.

72
Εικόνα Για το λυτικό καταρράκτη απαιτείται η πρωτεΐνη Cro, η οποία εμποδίζει την ενεργοποίηση του κυκλώματος διατήρησης της λυσιγονίας που λειτουργεί μέσω του PRM και παράλληλα καταστέλλει την έκφραση των καθυστερημένα πρώιμων γονιδίων, αποτρέποντας έτσι έμμεσα την εγκαθίδρυση του καταστολέα.

73
Πώς καθορίζεται η ισορροπία ανάμεσα στη λυσιγονία και στο λυτικό κύκλο;
Το κρίσιμο γεγονός για τη λυσιγονία είναι η πρόσδεση του cI στις θέσεις OL1 και OR1. Το κρίσιμο γεγονός για το λυτικό κύκλο είναι η πρόσδεση της Cro στη θέση OR3.

74
Εικόνα Η κρίσιμη φάση στην οποία λαμβάνεται η απόφαση για το αν θα ακολουθηθεί το μονοπάτι της λυσιγονίας ή του λυτικού κύκλου είναι όταν εκφραστούν τα καθυστερημένα πρώιμα γονίδια. Εάν τα επίπεδα της cII επαρκούν για να υποστηρίξουν την παραγωγή ικανής ποσότητας καταστολέα, τότε ακολουθείται το μονοπάτι της λυσιγονίας, γιατί ο καταστολέας καταλαμβάνει τους χειριστές. Διαφορετικά, οι χειριστές καταλαμβάνονται από την Cro και ακολουθείται το μονοπάτι του λυτικού κύκλου

75
Το βασικό στοιχείο που καθορίζει την έκβαση της μόλυνσης είναι η δράση της cII.
Αν η cII είναι αρκετή, τότε η σύνθεση του καταστολέα (cI) μέσω του PRE είναι αποτελεσματική και κατά συνέπεια ο καταστολέας καταλαμβάνει τους χειριστές. Αν η cII δεν είναι αρκετή τότε η Cro προσδένεται στους χειριστές. Η δράση της cIII είναι έμμεση: βοηθά στην προστασία της cII έναντι των πρωτεασών.
 μέσω του PRE είναι αποτελεσματική και κατά συνέπεια ο καταστολέας καταλαμβάνει τους χειριστές. Αν η cII δεν είναι αρκετή τότε η Cro προσδένεται στους χειριστές. Η δράση της cIII είναι έμμεση: βοηθά στην προστασία της cII έναντι των πρωτεασών.")
77
Ο ξενιστής ενέχεται στη διαδικασία λήψης της απόφασης:
Η cII είναι ευαίσθητη στις πρωτεάσες του ξενιστή. Οι πρωτεάσες αυτές ενεργοποιούνται κατά την αύξηση σε πλούσιο θρεπτικό υλικό. Ο φάγος τείνει να λύει τα κύτταρα που πολλαπλασιάζονται με γοργό ρυθμό, αλλά είναι πιο πιθανό να ακολουθήσει τη λυσιγονία όταν τα κύτταρα βρίσκονται σε συνθήκες πείνας.

78
Genes VIII, Κεφάλαιο 11: 11.11 11.16 και 11.17 Genes VIII, Κεφάλαιο 9:
9.19 Genes VIII, Κεφάλαιο 12: 12.1 12.3 έως 12.4 12.6 έως 12.9 12.14 έως 12.20

Παρόμοιες παρουσιάσεις




, είναι δύο μέλη της οικογένειας του ιού του έρπητα, herpesviridae, που μολύνει.>")















