Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
09-Σύνθεση και επεξεργασία ευκαρυωτικού RNA

2
Το ευκαρυωτικό mRNA τροποποιείται κατά τη διάρκεια ή μετά το πέρας της μεταγραφής του

3
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 5.16 Το ευκαρυωτικό mRNA τροποποιείται με την προσθήκη μιας καλύπτρας στο 5΄ άκρο και μιας ουράς πολυ (Α) στο 3΄ άκρο. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
 στο 3΄ άκρο. Genes VIII - Ακαδημαϊκές Εκδόσεις")
4
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 5.17 Επισκόπηση: Η έκφραση του mRNA σε ζωικά κύτταρα απαιτεί μεταγραφή, τροποποίηση, επεξεργασία, πυρηνο-κυτταροπλασματική μεταφορά και μετάφραση. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

5
Η μεταγραφή στα ζωικά κύτταρα συμβαίνει με την ίδια περίπου ταχύτητα που συμβαίνει και στα βακτήρια (~40 b / sec). Για ένα γονίδιο 10,000 bp χρειάζονται ~5 λεπτά για να μεταγραφεί.

7
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 5.18 Η καλύπτρα ασφαλίζει το 5΄ άκρο του mRNA και μπορεί να μεθυλιωθεί σε αρκετές θέσεις.

8
Η καλύπτρα 0 υπάρχει σε όλους τους ευκαρυώτες
Η καλύπτρα 0 υπάρχει σε όλους τους ευκαρυώτες. Στους μονοκύτταρους ευκαρυώτες υπάρχει μόνο αυτή (Το υπεύθυνο ένζυμο είναι η 7-μεθυλο-τρανσφεράση της γουανίνης). Η καλύπτρα 1 είναι ο κύριος τύπος καλύπτρας εκτός από τους μονοκύτταρους. Η καλύπτρα 2 αντιπροσωπεύει λιγότερα από το 10-15% του συνολικού πληθυσμού του mRNA που είναι καλυμμένο. Οι ποσοστιαίες αναλογίες των διαφόρων τύπων καλύπτρας είναι χαρακτηριστικές για ένα συγκεκριμένο οργανισμό.
. Η καλύπτρα 1 είναι ο κύριος τύπος καλύπτρας εκτός από τους μονοκύτταρους. Η καλύπτρα 2 αντιπροσωπεύει λιγότερα από το 10-15% του συνολικού πληθυσμού του mRNA που είναι καλυμμένο. Οι ποσοστιαίες αναλογίες των διαφόρων τύπων καλύπτρας είναι χαρακτηριστικές για ένα συγκεκριμένο οργανισμό.")
9
Το 3’ άκρο πολυαδενυλιώνεται
CPSF: cleavage and polyadenylation specificity factor CstF: cleavage stimulation factor rich

10
Η οργάνωση του γονιδίου της β-σφαιρίνης του ανθρώπου

11
Το μάτισμα και η επεξεργασία του RNA

12
Ασυνεχή γονίδια απαντώνται σε όλες τις τάξεις των οργανισμών:
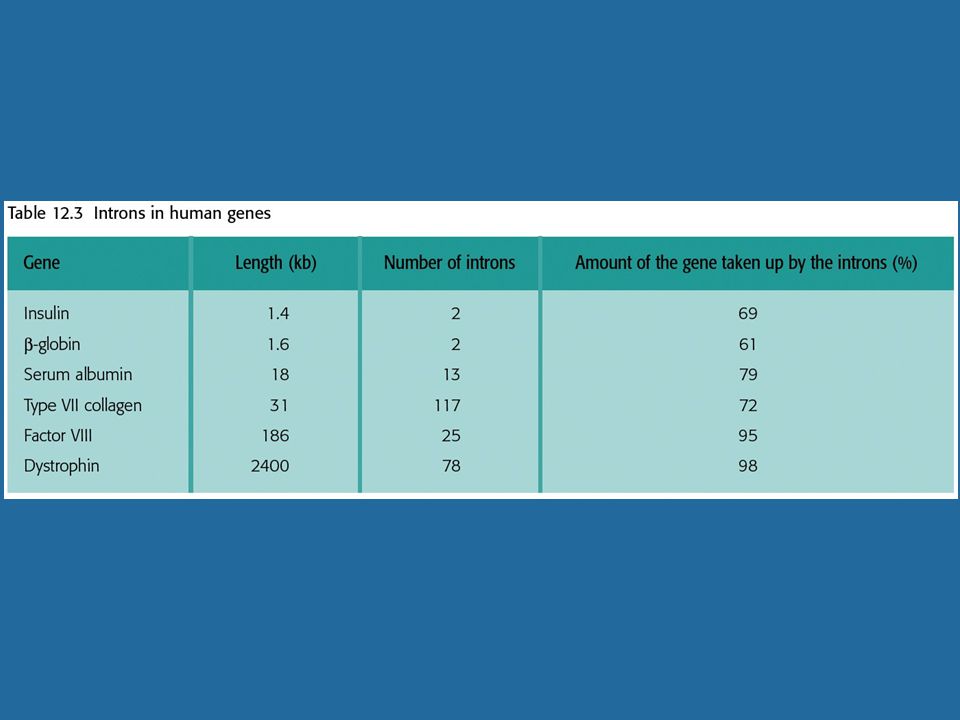
Μικρό ποσοστό σε κατώτερους ευκαρυώτες Πλειοψηφία στους ανώτερους ευκαρυώτες Τυπικό γονίδιο θηλαστικού: 7-8 εξόνια σε έκταση ~16kb. Εξόνια: ~ bp, ιντρόνια: >1kb.

14
Το μάτισμα εμπλέκεται στην παραγωγή μεγαλύτερου ποσοστού (50%) mRNA από το αναμενόμενο βάσει της ανάλυσης του γονιδιώματος. Το πυρηνικό RNA (hnRNA) έχει μέσο μέγεθος κατά πολύ μεγαλύτερο του mRNA, είναι ασταθές και η αλληλουχία του εμφανίζει μεγαλύτερη πολυπλοκότητα.
 έχει μέσο μέγεθος κατά πολύ μεγαλύτερο του mRNA, είναι ασταθές και η αλληλουχία του εμφανίζει μεγαλύτερη πολυπλοκότητα.")
15
Εικόνα 24.1 Το hnRNA απαντάται ως ριβονουκλεοπρωτεϊνικό σύμπλοκο με μορφή μιας σειράς από χάντρες.
Περίπου 20 πρωτεΐνες συμμετέχουν στη δημιουργία του hnRNA. Οι πρωτεΐνες αυτές τυπικά υπάρχουν σε ~108 αντίγραφα ανά πυρήνα, συγκρινόμενες με τα ~106 μόρια του hnRNA. Κάποιες μπορεί να έχουν δομικό ρόλο στο πακετάρισμα. Άλλες παλινδρομούν μεταξύ του πυρήνα και του κυτταροπλάσματος συμμετέχοντας στην εξαγωγή του RNA από τον πυρήνα.

16
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 24.2 Το RNA τροποποιείται στον πυρήνα με προσθήκες στα 5΄ και 3΄ άκρα και με μάτισμα, ώστε να αφαιρεθούν τα ιντρόνια. Η διαδικασία του ματίσματος απαιτεί τη διάσπαση των θέσεων συνένωσης εξονίου-ιντρονίου και τη συρραφή των άκρων των εξονίων. Το ώριμο mRNA μεταφέρεται μέσα από τους πυρηνικούς πόρους στο κυτταρόπλασμα, όπου μεταφράζεται. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

17
ΕΙΚΟΝΑ 5.1: Εξέταση της δομής των γονιδίων με ηλεκτρονική μικροσκοπία υβριδίων DNA-mRNA.

18
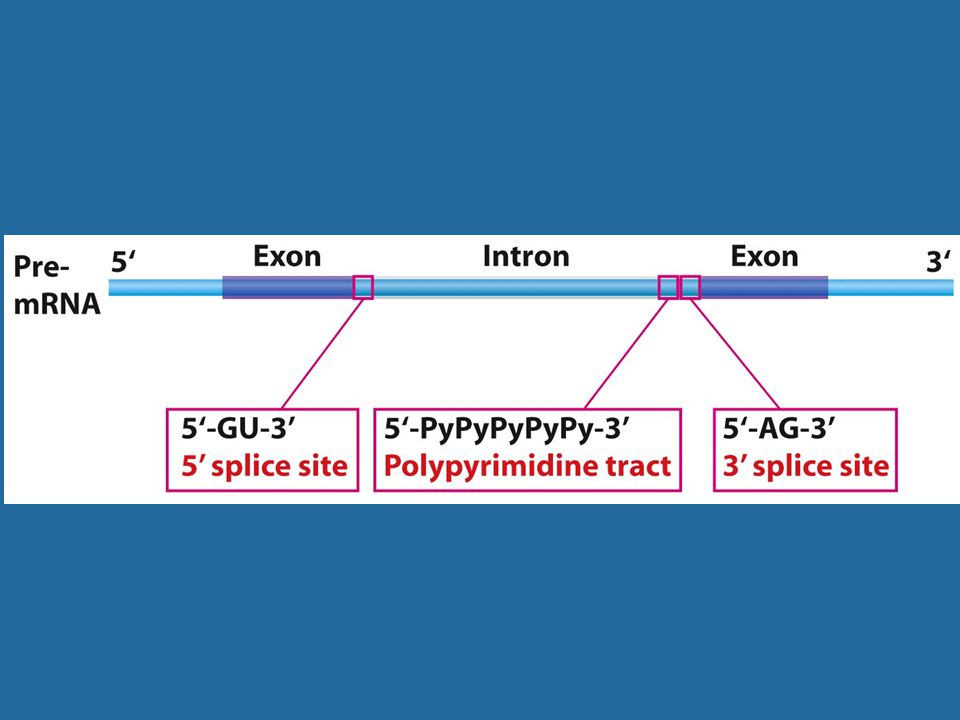
Οι θέσεις ματίσματος είναι βραχείες αλληλουχίες
Τα σημεία συνένωσης διαθέτουν καλά συντηρημένες, αν και μικρές σε μήκος, πρότυπες αλληλουχίες.

19
Εικόνα 24.3 Τα άκρα των πυρηνικών ιντρονίων καθορίζονται από τον κανόνα GU-AG.
Οι δύο θέσεις ματίσματος έχουν διαφορετική αλληλουχία και έτσι προσδιορίζουν τη φορά του ιντρονίου. Σημειακές μεταλλάξεις στις θέσεις ματίσματος (GT-AG) παρεμποδίζουν το μάτισμα είτε in vivo είτε in vitro, αποδεικνύοντας ότι οι συντηρημένες αλληλουχίες είναι πράγματι αυτές που αναγνωρίζονται κατά το μάτισμα.
 παρεμποδίζουν το μάτισμα είτε in vivo είτε in vitro, αποδεικνύοντας ότι οι συντηρημένες αλληλουχίες είναι πράγματι αυτές που αναγνωρίζονται κατά το μάτισμα.")
20
Οι θέσεις ματίσματος διαβάζονται κατά ζεύγη

21
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα 24.4 Μόνο οι σωστοί συνδυασμοί θέσεων ματίσματος αναγνωρίζονται ανά ζεύγη. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

22
Όλες οι 5' θέσεις ματίσματος είναι λειτουργικά ισοδύναμες και όλες οι 3' θέσεις είναι επίσης λειτουργικά ισοδύναμες. Παρόλα αυτά, το μάτισμα πραγματοποιείται μεταξύ των 5' και 3' θέσεων του ίδιου ιντρονίου.

23
Με ποια σειρά αφαιρούνται τα ιντρόνια από ένα μόριο RNA;

24
Εικόνα 24.5 Στύπωμα τύπου Northern πυρηνικού RNA χρησιμοποιώντας το γονίδιο μιας ωοβλεννοειδούς πρωτεΐνης ως ιχνηθέτη δείχνει διακριτά ενδιάμεσα μόρια κατά το μάτισμα. Αναγράφονται τα συστατικά τμήματα των κυρίαρχων ζωνών. Η φωτογραφία είναι ευγενική προσφορά του Bert O’Malley. Εάν τα ιντρόνια αφαιρούνταν με τυχαία σειρά θα υπήρχαν περισσότερα από 300 πρόδρομα μόρια. Εν τούτοις, δεν υπάρχει ένα μοναδικό μονοπάτι, καθώς ανιχνεύονται ενδιάμεσα μόρια από τα οποία έχουν αφαιρεθεί διαφορετικοί συνδυασμοί ιντρονίων.

25
Το πρόδρομο mRNA σχηματίζει δομή θηλιάς κατά το μάτισμα

27
Splicing in outline

28
Η θέση διακλάδωσης διαδραματίζει σημαντικό ρόλο στην αναγνώριση της 3' θέσης ματίσματος.
Στο ζυμομύκητα είναι συντηρημένη. Ελλείμματα στη θέση διακλάδωσης αποτρέπει το μάτισμα. Στους ανώτερους ευκαρυώτες είναι λιγότερο συντηρημένη => σε περίπτωση ελλείμματος χρησιμοποιούνται κρυφές θέσεις, που όμως βρίσκονται στη γειτονιά. => Ο ρόλος της θέσης διακλάδωσης είναι να αναγνωρίζει την πλησιέστερη 3' θέση ματίσματος.

29
Στο μάτισμα είναι απαραίτητα ορισμένα snRNA
Εικόνα 24.8 Το σωμάτιο ματίσματος έχει μοριακό βάρος ~12 MDa. Τα πέντε snRNP αντιστοιχούν σχεδόν στο μισό της μάζας του. Οι υπόλοιπες πρωτεΐνες περιλαμβάνουν γνωστούς παράγοντες ματίσματος, καθώς και πρωτεΐνες που εμπλέκονται σε άλλα στάδια της γονιδιακής έκφρασης.

30
Οι ρόλοι των snRNPs και των συνδεόμενων πρωτεϊνών κατά το μάτισμα

31
Μια εναλλακτική συσκευή ματίσματος χρησιμοποιεί διαφορετικά snRNP
98% των ιντρονίων του ανθρώπινου γονιδιώματος είναι GU-AG. Ένα <1% χρησιμοποιεί GC-AG. ~0,1% είναι AU-AC. Χρησιμοποιούν εναλλακτικές συσκευές, U11 και U12 (συγγενικά των U1 και U2).
.")
32
Το μάτισμα συνδέεται με την εξαγωγή του mRNA από τον πυρήνα
Οι πρωτεΐνες που ευθύνονται για τη μεταφορά ματισμένων προϊόντων παλινδρομούν ανάμεσα στον πυρήνα και το κυτταρόπλασμα. Πώς αναγνωρίζουν τα RNA-υποστρώματα και τι εγγυάται ότι μόνο τα πλήρως επεξεργασμένα mRNA εξάγονται;

33
Εικόνα Το σύμπλοκο EJC (Exon Junction Complex) προσδένεται στο RNA αναγνωρίζοντας το σωμάτιο ματίσματος. Εάν αφαιρέσουμε τα ιντρόνια από ένα γονίδιο, τότε το RNA προϊόν εξάγεται πολύ πιο αργά από το κυτταρόπλασμα

34
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Η πρωτεΐνη REF προσδένεται σε έναν παράγοντα ματίσματος και παραμένει με το ματισμένο προϊόν. Η REF προσδένεται σε έναν παράγοντα εξαγωγής από τον πυρήνα, ο οποίος με τη σειρά του συνδέεται με τον πυρηνικό πόρο. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

35
Το εναλλακτικό μάτισμα

36
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Το εναλλακτικό μάτισμα μπορεί να παραγάγει μια ποικιλία πρωτεϊνικών προϊόντων από το ίδιο γονίδιο. Η αλλαγή των θέσεων ματίσματος μπορεί να εισαγάγει κωδικόνια τερματισμού (αστερίσκοι) ή να αλλάζει το πλαίσιο ανάγνωσης. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
 ή να αλλάζει το πλαίσιο ανάγνωσης. Genes VIII - Ακαδημαϊκές Εκδόσεις")
37
Alternative splicing

38
Σκιαγράφηση του φυλοκαθοριστικού μονοπατιού στη Drosophila

39
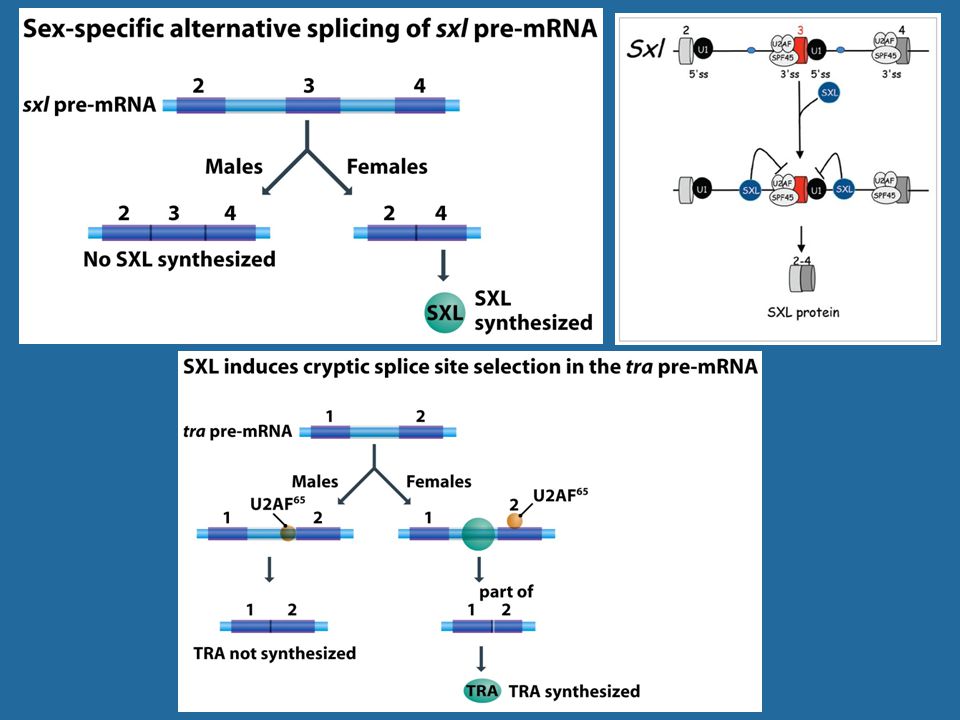
Η διαφορική ενεργοποίηση του γονιδίου sxl σε θηλυκά και αρσενικά

42
Exonic Splicing Enhancer: Ενισχυτής ματίσματος για την παρακείμενη 3' θέση

43
Dsx alternative splicing in males and females
Pre-mRNAs from the D. melanogaster gene dsx contain 6 exons. In males, exons 1,2,3,5,and 6 are joined to form the mRNA, which encodes a transcriptional regulatory protein required for male development. The exon 4 is eliminated. In females, Tra1 and Tra2 in splicing between 3 and 4 th exons, thus exons 1,2,3, and 4 are joined, and a polyadenylation signal in exon 4 causes cleavage of the mRNA at that point. The resulting mRNA is a transcriptional regulatory protein required for female development. This is an example of exon skipping alternative splicing. The intron upstream from exon 4 has a weak-consensus polypyrimidine tract, to which U2AF proteins bind poorly without assistance from splicing activators. This 3' splice acceptor site is therefore not used in males. Females, however, produce the splicing activator Transformer (Tra). The SR protein Tra2 is produced in both sexes and binds to an ESE in exon 4; if Tra is present, it binds to Tra2 and, along with another SR protein, forms a complex that assists U2AF proteins in binding to the weak polypyrimidine tract. U2 is recruited to the associated branch point, and this leads to inclusion of exon 4 in the mRNA. The Dsx protein in female prevents male sex organ differentiation and development and the Dsx protein in females blocks male sex organ development.
. The SR protein Tra2 is produced in both sexes and binds to an ESE in exon 4; if Tra is present, it binds to Tra2 and, along with another SR protein, forms a complex that assists U2AF proteins in binding to the weak polypyrimidine tract. U2 is recruited to the associated branch point, and this leads to inclusion of exon 4 in the mRNA. The Dsx protein in female prevents male sex organ differentiation and development and the Dsx protein in females blocks male sex organ development.")
44
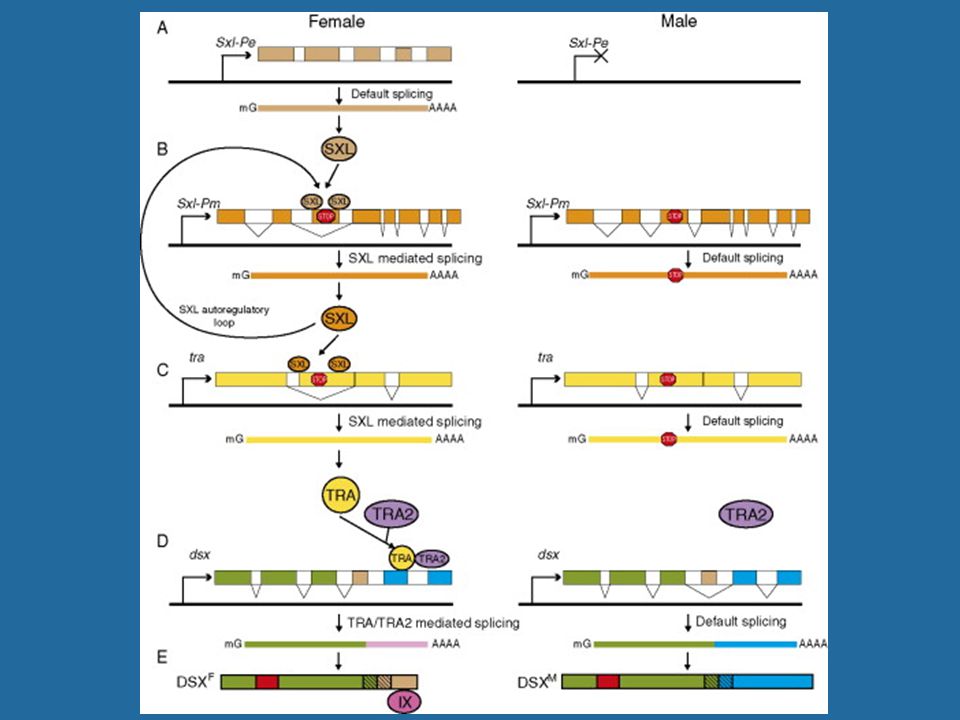
The pattern of sex-specific RNA splicing in three major Drosophila sex-determining genes

45
Details of proposed regulation cascade for Drosophila somatic sex determination

46
Τα 3' άκρα των μεταγράφων polI και polIΙI καθορίζονται κατά τον τερματισμό της μεταγραφής τους

47
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Κατά τη δημιουργία του 3΄ άκρου με τερματισμό της μεταγραφής, η RNA πολυμεράση και το RNA αποδεσμεύονται σε μια διακριτή αλληλουχία τερματισμού του DNA. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

48
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Κατά τη δημιουργία του 3΄ άκρου με αποκοπή, η RNA πολυμεράση συνεχίζει τη μεταγραφή, και μια ενδονουκλεάση αποκόπτει το RNA σε μια καθορισμένη θέση. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

49
Το προϊόν της μεταγραφής της polI είναι ένα μεγάλο πρόδρομο rRNA.
Το μόριο αυτό υφίσταται εκτεταμένη επεξεργασία. Ο τερματισμός συμβαίνει σε απόσταση ~1000 βάσεων καθοδικά του 3' άκρου του ώριμου rRNA. Ο τερματισμός προϋποθέτει την αναγνώριση αλληλουχίας τερματισμού 18 βάσεων από βοηθητικό παράγοντα.

50
Η αντίδραση τερματισμού της polIII (5S rRNA, tRNAs, small RNAs) μοιάζει με αυτή των προκαρυωτών.
Ο τερματισμός συμβαίνει συνήθως στη 2η U σε σειρά 4 U σε περιοχή πλούσια σε GC.

51
Το 3' άκρο του mRNA δημιουργείται με αποκοπή και πολυαδενυλίωση
Η αλληλουχία AAUAA είναι ένα σήμα αποκοπής για να δημιουργηθεί το 3' άκρο του mRNA, το οποίο κατόπιν πολυαδενυλιώνεται.

52
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Η αλληλουχία AAUAAA είναι απαραίτητη για την αποκοπή που δημιουργεί το 3΄ άκρο για την πολυαδενυλίωση. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

53
Εικόνα Το σύμπλοκο επεξεργασίας του 3΄ άκρου περιλαμβάνει αρκετές ενζυμικές ενεργότητες. Οι παράγοντες CPSF και CstF αποτελούνται από αρκετές υπομονάδες. Τα άλλα συστατικά είναι μονομερή. Η συνολική μάζα του συμπλόκου ξεπερνάει τα 900 kD. Cleavage and Polyadenylation Specificity Factor (CPSF) Cleavage Factor I and II (CFI & CFII) PolyA Polymerase (PAP) Cleavage stimulatory Factor (CstF)
 Cleavage Factor I and II (CFI & CFII) PolyA Polymerase (PAP) Cleavage stimulatory Factor (CstF)")
54
Δεν είναι όλα τα mRNA πολυαδενυλιωμένα.

55
Εικόνα 5.19 Το πολυ(Α)+ RNA μπορεί να διαχωριστεί από τα άλλα RNA με κλασμάτωση σε στήλη σεφαρόζης-ολιγο(dT).
+ RNA μπορεί να διαχωριστεί από τα άλλα RNA με κλασμάτωση σε στήλη σεφαρόζης-ολιγο(dT).")
56
Τα μεγάλα και τα μικρά rRNA αποδεσμεύονται με αποκοπή από ένα κοινό πρόδρομο RNA

57
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Τα ώριμα ευκαρυωτικά rRNA δημιουργούνται από ένα πρωτογενές μετάγραφο με φαινόμενα αποκοπής και περιορισμού των άκρων. To 5S rRNA μεταγράφεται από ξεχωριστά γονίδια μέσω της RNA pol III. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004

58
Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
Εικόνα Τα οπερόνια rrn στο βακτήριο E. coli περιέχουν γονίδια που κωδικοποιούν τόσο για rRNA όσο και για tRNA. Το ακριβές μήκος των μεταγράφων εξαρτάται από τους υποκινητές (P) και τις αλληλουχίες τερματισμού (t) που χρησιμοποιούνται. Κάθε προϊόν RNA πρέπει να απελευθερωθεί από το μετάγραφο με αποκοπή στα δύο του άκρα. Genes VIII - Ακαδημαϊκές Εκδόσεις 2004
 και τις αλληλουχίες τερματισμού (t) που χρησιμοποιούνται. Κάθε προϊόν RNA πρέπει να απελευθερωθεί από το μετάγραφο με αποκοπή στα δύο του άκρα. Genes VIII - Ακαδημαϊκές Εκδόσεις")
59
Τι να μελετήσετε Από Genes VIII 5.8 έως 5.10 24.1 έως 24.3 24.12
24.18 και 24.19 24.21

Παρόμοιες παρουσιάσεις