Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Enzimoloģija Telomēri un Telomerāze Lit.: C.B.Harley, Nature Reviews, 2008, v.8, 167-179 R.E., Verdun, J.Karlseder, Nature, 2007, v.447, 924-931 S.B;.lCohen, R.R.Reddel, Nature Methods, 2008,v.5,355-360 Telomeres, Eds. T de Lange, V.Lundblad, E.Blackburn, Cold Spring Harbor Lab. Press, 2006,576 lpp http://www.telomere.net/ Molekulāras bioloģijas (bioķīmijas)katedra 2009/2010. akad.gads
katedra 2009/2010. akad.gads.")
2
C.W.Greider, E.H.Balckburn – Identification of a specific telomere terminal transferase enzyme with two kinds of primer specificity, Cell, 1985, v.51, p.405-413

3
Ko redz šūnas kodolā ?

5
Telomēri un telomerāze nosaka šūnu nemirstību Telomēri izvietoti hromosomu galos, to cilvēkam ir 2x23x2=92 To garums ir līdz 15 kbp

6
Kas ir telomēri? Telomēri ir specializēti nukleoproteīnu kompleksi, kuriem ir svarīga funkcija hromosomu aizsardzībā, replikācijā un hromosomu nobeigumu stabilizācijā; Telomēri ir fizikālie lineāro hromosomu nobeigumi eikariotu hromosomās; Vairumā izpētīto organismu telomēri satur garus posmus ar tandēmi atkārtotām, vienkāršām DNS sekvencēm, kas veidotas no G – un C- bagātiem posmiem (tā saucamie terminālie atkārtojumi); Šie terminālie atkārtojumi ir ļoti konservatīvi; patiesībā liekas visiem mugurkaulniekiem ir viens un tas pats vienkāršais atkārtojums (TTAGGG)n; Bieži sekvences, kas atrodas tuvu telomēru atkārtojumiem ir ļoti polimorfas un bagātas ar DNS atkārtojumu elementiem (subtelomēriskie atkārtojumi). Tomēr, dažos gadījumos ir atrasti gēni hromosomu proterminālajā reģionā;
; Šie terminālie atkārtojumi ir ļoti konservatīvi; patiesībā liekas visiem mugurkaulniekiem ir viens un tas pats vienkāršais atkārtojums (TTAGGG)n; Bieži sekvences, kas atrodas tuvu telomēru atkārtojumiem ir ļoti polimorfas un bagātas ar DNS atkārtojumu elementiem (subtelomēriskie atkārtojumi). Tomēr, dažos gadījumos ir atrasti gēni hromosomu proterminālajā reģionā;.")
7
Telomēri citos organismos GrupaOrganismsTelomēri MugurkaulniekiH. sapiens, M. musculis TTAGGG SēnesNeurospora crassa TTAGGG Augstākie augiArabidopsis thaliana TTTAGGG RaugsSaccharomyce s cerevisiae Schizosacchar omyces pombe TGTGGGTGTG GTG TTAGGG

8
T cilpa Vienpavediena pārkare D cilpa

9
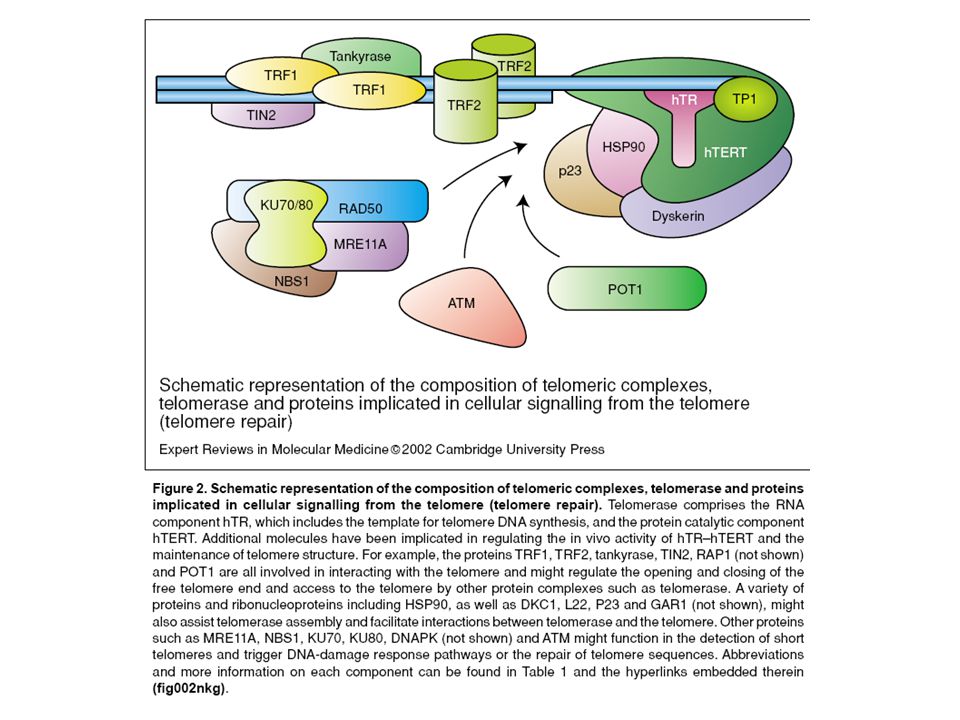
Ar telomēriem saistītos proteīnus mēģina dalīt grupās: 1.Telomērus ekranējošie (sheltering) proteīni: Šie proteīni tieši pazīst TTAGGG heksamēru TRF1, TRF2, Pot1 Šie saistās ar telomēriem caur pirmās grupas proteīniem Tin2, TPP1, RAP1 2. Ar ekranējošajiem proteīniem saistītie citi proteīni (telomeric proteins associated with the shelterin complex) Tankirāze 1 un 2, ANKYRIN- related ADP-ribose polymerase PINX1 WRN
 Tankirāze 1 un 2, ANKYRIN- related ADP-ribose polymerase PINX1 WRN.")
10
Ar telomēriem saistītos proteīnus mēģina dalīt grupās: 3. Telomēru proteīni, kuri saistīti ar DNS reparāciju: ATM (Ataxia teleangectasia mutated) u.c. 4. Telomerāzes komplekss: a) TERC TERT b) RNS stabilizējošie faktori DKS – diskerīns NOLA – nucleolar protein family A... HEST 1 A – human ever shorter telomeres c) heterogēni ar telomerāzi asociēti kodola ribonukleoproteīni, hn RNP... d) Šaperoni HSP90A P23 HSP-71-1
 u.c. 4. Telomerāzes komplekss: a) TERC TERT b) RNS stabilizējošie faktori DKS – diskerīns NOLA – nucleolar protein family A... HEST 1 A – human ever shorter telomeres c) heterogēni ar telomerāzi asociēti kodola ribonukleoproteīni, hn RNP... d) Šaperoni HSP90A P23 HSP")
11
TRF- telomeric repeat binding factor RAP- transcriptional repressor/activator protein POT- protection of telomeres TIN - TRF interacting protein TANC - TRF un RAP (PARP) pārveidotāji PARP- poly(ADP)-ribose polymerase MRE- meyotic recombination 11 homologue ERCC - excision repair cross complementing 1 BLM- Bloom syndrome protein WRN - Werner syndrome protein BLM- Bloom syndrome protein WRN - Werner syndrome protein ATM - ataxia- teleangiectasia mutated Rad - DNA-repair protein Telomērus ekranējošie (sheltering) proteīni
 pārveidotāji PARP- poly(ADP)-ribose polymerase MRE- meyotic recombination 11 homologue ERCC - excision repair cross complementing 1 BLM- Bloom syndrome protein WRN - Werner syndrome protein BLM- Bloom syndrome protein WRN - Werner syndrome protein ATM - ataxia- teleangiectasia mutated Rad - DNA-repair protein Telomērus ekranējošie (sheltering) proteīni")
12
Telomēru “cap” struktūra sastāv no “D-” un “T - cilpas”; Telomēru “D-cilpu” veido telomerāzes subvienības (TERT un TERC); Telomēru distālajā galā ir 300 nt vienpavediena secība, kura veido “T-cilpu” (T-loop); “T-cilpa” stabilizē telomēru struktūru, pasargājot telomēru galus no DNS reparācijas mašinērijas; “T-cilpa” tiek noturēta savā telpiskā konformācijā ar proteīnu kompleksu palīdzību (TRF1 un TRF2) Telomēru nukleoproteīna komplekss
; Telomēru distālajā galā ir 300 nt vienpavediena secība, kura veido T-cilpu (T-loop); T-cilpa stabilizē telomēru struktūru, pasargājot telomēru galus no DNS reparācijas mašinērijas; T-cilpa tiek noturēta savā telpiskā konformācijā ar proteīnu kompleksu palīdzību (TRF1 un TRF2) Telomēru nukleoproteīna komplekss")
13
Telomēru komponentu raksturojums Telomēru sintēzi veic RNS atkarīgā DNS polimerāze –telomerāze (TERT) par matricu izmantojot RNS (TERT), Telomēru struktūrā bez DNS, TERT un TERC ir iesaistīti proteīni: –T–TRF1 (Telomere repeat binding factor) – saistās pie DNS telomēru rajonā un regulē telomēru garumu atkarībā no telomerāzes; –T–TIN2 – telomerāzes-atkarīgais telomēru garuma regulētājs; –T–TANK1: TRF1 un PARP pārveidotājs un telomēru garuma regulētājs; –T–TRF2 (Telomere repeat binding factor 2) – saistās pie DNS telomēru rajonā ar (end capping) funkciju un no telomerāzes- neatkarīgais telomēru garuma regulētājs; –R–RAP1: mijiedarbojas ar TRF2 un ir telomēru garuma regulētājs; –T–TANK2: TRF1 un PARP pārveidotājs; –E–ERCC1/XPF: endonukleāze, kas darbojas kopā ar TRF2; –M–MRN komplekss: mijiedarbojas ar TRF2; –R–Rif1: mijiedarbojas ar TRF2 un in vivo komponents peļu telomēros; –P–POT1: mijiedarbojas ar TRF1 un saistās pie vienpavediena DNS telomēru rajonā ar telomēru garuma regulācijas funkciju; –K–Ku: in vivo telomēru komponents un telomēru garuma regulētājs (?); –D–DNS-PKcs: in vivo telomēru komponents ar telomēru capping funkciju; –E–EST1A/B: telomēru garuma regulētājs (?);
 par matricu izmantojot RNS (TERT), Telomēru struktūrā bez DNS, TERT un TERC ir iesaistīti proteīni: –T–TRF1 (Telomere repeat binding factor) – saistās pie DNS telomēru rajonā un regulē telomēru garumu atkarībā no telomerāzes; –T–TIN2 – telomerāzes-atkarīgais telomēru garuma regulētājs; –T–TANK1: TRF1 un PARP pārveidotājs un telomēru garuma regulētājs; –T–TRF2 (Telomere repeat binding factor 2) – saistās pie DNS telomēru rajonā ar (end capping) funkciju un no telomerāzes- neatkarīgais telomēru garuma regulētājs; –R–RAP1: mijiedarbojas ar TRF2 un ir telomēru garuma regulētājs; –T–TANK2: TRF1 un PARP pārveidotājs; –E–ERCC1/XPF: endonukleāze, kas darbojas kopā ar TRF2; –M–MRN komplekss: mijiedarbojas ar TRF2; –R–Rif1: mijiedarbojas ar TRF2 un in vivo komponents peļu telomēros; –P–POT1: mijiedarbojas ar TRF1 un saistās pie vienpavediena DNS telomēru rajonā ar telomēru garuma regulācijas funkciju; –K–Ku: in vivo telomēru komponents un telomēru garuma regulētājs ( ); –D–DNS-PKcs: in vivo telomēru komponents ar telomēru capping funkciju; –E–EST1A/B: telomēru garuma regulētājs ( );")
14
Telomerāze uzsintezē vienu DNS pavedienu, kā matricu izmantojot RNS, bet otru pavedienu sintezē DNS polimerāze uz tikko uzsintezētās DNS matricas

15
Telomerāzes darbības mehanisms

16
Telomēru garuma noteikšanas metodes Southern hybridization/Southern blot Hybridization protection assay Fluorescent in situ hybridization –F–FISH –Q–Q-FISH –Q–Q-FISH FCM –F–Flow FISH

17
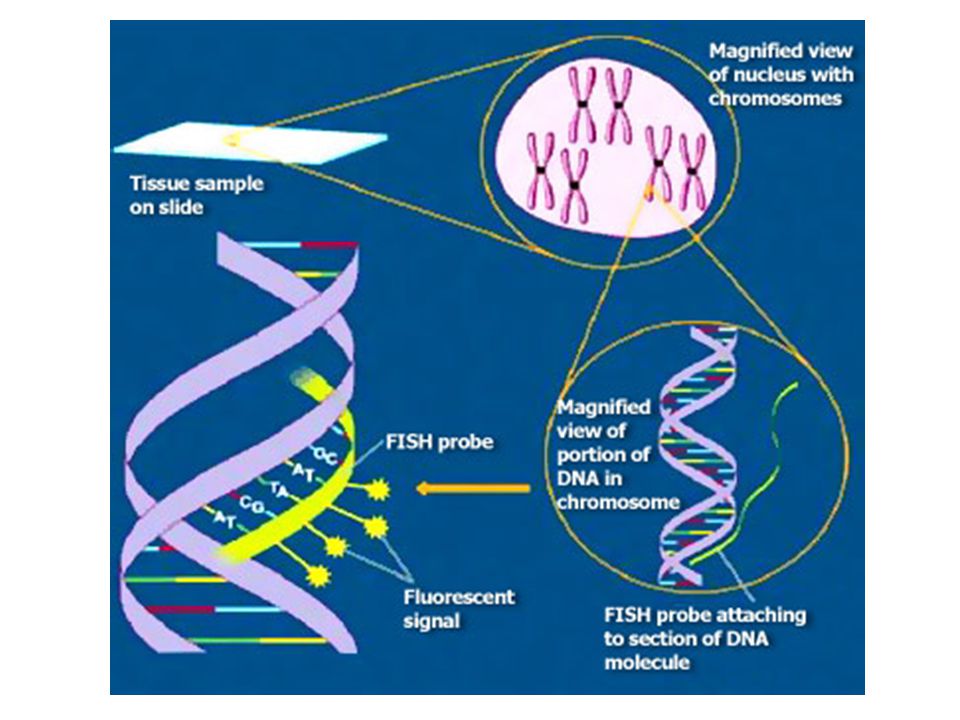
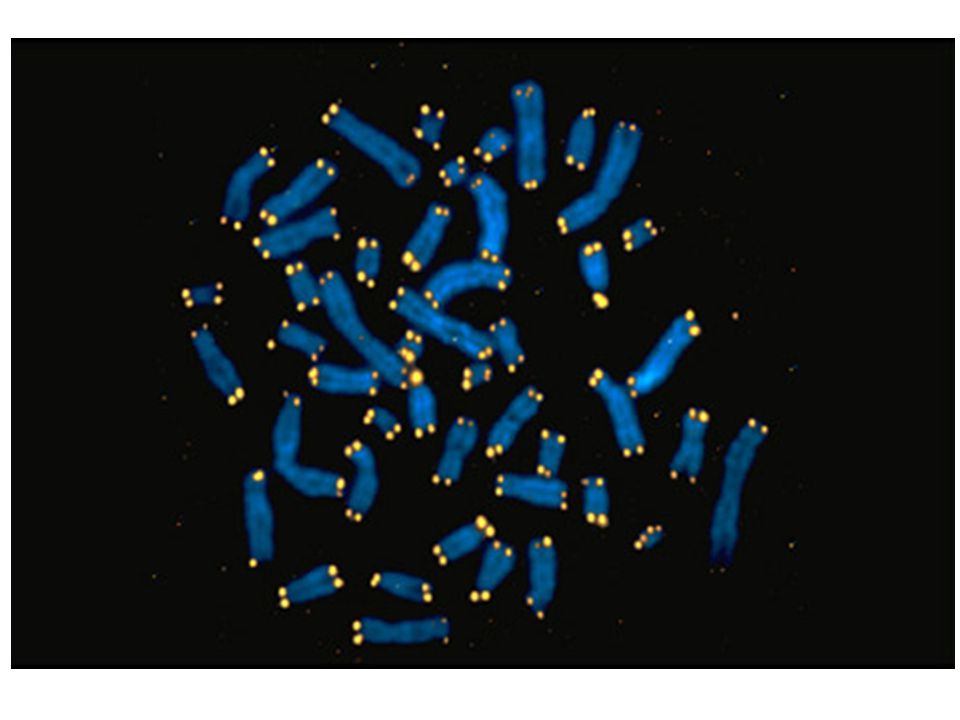
FISH FISH metode ietver šūnu fiksāciju, kam seko apstrādāšana ar hibridizācijas materiālu, kas satur formamīdu, fluorescentās nukleīnskābes/peptīda (PNA) zondi, kas ir komplementāra telomēru atkārtojumiem. Flourescentā iezīmēšana dod iespēju tieši izmērīt telomēru garumu ar kvantitatīvu noteikšanas metodi, kas ir Q-FISH. PNA zondēm ir neitrāls glicīna pamats, kas nodrošina ciešu PNA-DNS saistību, atšķirībā no citām zondēm. Flourescentais signāls ko izdala telomērs atbilst tā garumam un ar speciālu programmatūru, izanalizējot šo attēlu var izrēķināt individuālu telomēru signāla stiprumu. Telomēru garumu izsaka telomēru fluorescentajās vienībās (TFU), 1TFU atbilst 1kb lielam TTAGGG atkārtojumam.
, 1TFU atbilst 1kb lielam TTAGGG atkārtojumam..")
19
Iespējamais protokols eksperimentālai leikocitāro telomēru struktūras noteikšanai 1.Pacientu leikocītu izdalīšana –L–Leikocītu izdalīšana no svaigām ar EDTA apstrādātām asinīm ar DS (dekstrāna šķīdums) –L–Leikocītu izdalīšana no svaigām lizētām asinīm (DL metode) 2.Pacientu leikocītu kultivēšana –A–Audzē, piemēram, barotnē Glutamax-DMEM (Gibco/Invitrogen, Carlsband, CA) ar 15% FBS, 0,1mM dažādām aminoskābēm, un 100 vienības/ml penicilīna/streptomicīna. Kultivēšana: pie 7,5% CO 2 un 3% O 2 koncentrācijas. 3.Leikocītu dalīšanās indukcija –I–Inficē ar retrovīrusu vektoru HPV16-E6 4.Telomēru analīze ar FISH metodi metafāzes hromosomās –4–40% leu konfluences inkubē ar 0,1μg/ml demekolhīnu 3h –Š–Šūnas apstrādā ar tripsīnu, resuspendē 7,5mM KCl, inkubē 37 0 C 7min –Š–Šūnas fiksē ar metanola:etiķskābes (3:1) šķīdumu inkubē 4 0 C pa nakti –F–Fiksētās šūnas pārnes uz priekšmetstikliņa, skalo ar metanola/ etiķskābes šķīdumu un žāvē paaugstinātā mitrumā 80 0 C pa nakti –A–Apstrādā ar FITC-saistīto PNA zondi FITC-OO-[CCCTAA] 3 (Applied Biosystems) –D–DNS krāso ar DAPI, un kodolus aplūko mikroskopā (Zeiss Axioplan II) 5.Telomēru garuma noteikšana –I–Izmantojot Q-FISH metodi (Leica), attēli tiek apstrādāti ar MCK programmu (Leica Microsystems Imaging Solutions)
 šķīdumu inkubē 4 0 C pa nakti –F–Fiksētās šūnas pārnes uz priekšmetstikliņa, skalo ar metanola/ etiķskābes šķīdumu un žāvē paaugstinātā mitrumā 80 0 C pa nakti –A–Apstrādā ar FITC-saistīto PNA zondi FITC-OO-[CCCTAA] 3 (Applied Biosystems) –D–DNS krāso ar DAPI, un kodolus aplūko mikroskopā (Zeiss Axioplan II) 5.Telomēru garuma noteikšana –I–Izmantojot Q-FISH metodi (Leica), attēli tiek apstrādāti ar MCK programmu (Leica Microsystems Imaging Solutions).")
21
Kad telomēru struktūru būtu mērķtiecīgi pētīt Telomēru struktūras pētījumi (atkārtojumu skaits) dažādās vecuma grupās: –I–Ilgdzīvotāji (vecumā virs 90 gadiem) –I–Indivīdi vecumā no 60 līdz 80 gadiem –K–Kontroles grupa (vecumā no 18 līdz 40 gadiem) Telomēru garuma un struktūras pētījumi šūnās ar dažādu replikācijas potenciālu – tumoru šūnas, fibroblasti, hematopoētiskās sistēmas šūnas, neironi, cilmes šūnas Telomēru garuma atšķirību meklējumi dažādās hromosomās Telomēru garuma un struktūras pētījumi pacientiem ar progeroīdiem sindromiem
 dažādās vecuma grupās: –I–Ilgdzīvotāji (vecumā virs 90 gadiem) –I–Indivīdi vecumā no 60 līdz 80 gadiem –K–Kontroles grupa (vecumā no 18 līdz 40 gadiem) Telomēru garuma un struktūras pētījumi šūnās ar dažādu replikācijas potenciālu – tumoru šūnas, fibroblasti, hematopoētiskās sistēmas šūnas, neironi, cilmes šūnas Telomēru garuma atšķirību meklējumi dažādās hromosomās Telomēru garuma un struktūras pētījumi pacientiem ar progeroīdiem sindromiem")
22
Progeroīdais - Vernera (Werner) sindroms (VS) WS ir priekšlaicīgas novecošanās slimība, kas sākas pusaudža vecumā vai agrā pieaugušā vecumā un manifestējas 30-40 gadu vecumā Pacienti izskatās daudz vecāki par saviem gadiem. Fiziskās izpausmes var būt kā īss augums (bieži jau kopš bērnības) un citi simptomi, kas attīstās laika gaitā: krunkaina āda, plikpaurība, katarakta, muskuļu atrofija un tendence uz cukura diabētu Šī ir autosomāli recesīva patoloģija. Kultivējot VS pacientu šūnas, tām ir īsāka dzīvildze nekā šūnām no kontroles grupas. Gēns (WRN), kas ir iesaistīts šīs patoloģijas attīstībā lokalizējas 8. hromosomā un kodē helikāzi no RecQ helikāžu saimes. VS pacientiem papildus novecošanās fenotipam ir raksturīga arī genoma nestabilitāte, kas savukārt paaugstina risku izveidoties vēzim. Jaunākie pētījumi liecina par novecošanās fenotipa saistību ar telomēru disfunkciju. Pašlaik vēl nav noskaidrota saistība starp palielināto saslimstību ar vēzi un genoma nestabilitāti. Ir pierādīts ka VS gadījumā ar replikāciju saistītais telomēru zudums tieši ir atbildīgs par hromosomu saplūšanu ko novēro VS gadījumā.
 un citi simptomi, kas attīstās laika gaitā: krunkaina āda, plikpaurība, katarakta, muskuļu atrofija un tendence uz cukura diabētu Šī ir autosomāli recesīva patoloģija. Kultivējot VS pacientu šūnas, tām ir īsāka dzīvildze nekā šūnām no kontroles grupas. Gēns (WRN), kas ir iesaistīts šīs patoloģijas attīstībā lokalizējas 8. hromosomā un kodē helikāzi no RecQ helikāžu saimes. VS pacientiem papildus novecošanās fenotipam ir raksturīga arī genoma nestabilitāte, kas savukārt paaugstina risku izveidoties vēzim. Jaunākie pētījumi liecina par novecošanās fenotipa saistību ar telomēru disfunkciju. Pašlaik vēl nav noskaidrota saistība starp palielināto saslimstību ar vēzi un genoma nestabilitāti. Ir pierādīts ka VS gadījumā ar replikāciju saistītais telomēru zudums tieši ir atbildīgs par hromosomu saplūšanu ko novēro VS gadījumā..")
23
VS paciente 15 gadu vecumā VS paciente 48 gadu vecumā

24
Blūma sindroms - defekts 15q 26.1 hromosomā, DNA helikāzes grupas proteīni Pirmoreiz apraksta Dr. David Bloom 1954.g. Blūma sindroms ir autosomāla slimība, ko izsauc mutācija 15. hromosomā. Tādēļ šī slimība izpaužas jau zīdaiņa vecumā. Biežāk mutāciju sastop starp Aškenazi ebrejiem, kur 1 no 110 personām ir nēsātājs, kam gan gēna defekts neizpaužas. Slimību reģistrē dažādās etniskās grupās – amerikāņos, indiāņos, etiopos, turkos u.c. Molekulārais pamats Defektīvs gēns 15. hromosomā, kā rezultātā notiek nepareiza hromosomu reorganizācija un novēro hromosomas pārrāvumus. Fenotips Indivīdi ir īsa auguma (~1.5 m), bet var attīstīties normāli. Šaura seja, izteikts deguns. Sejas krāsa sārta, cieš no saules stariem, kuru ietekmē rodas raksturīgi izsitumi uz plakstiņiem, vaigiem, lūpām, ap degunu. Indivīdi cieš no elpošanas ceļu, ausu infekcijām. Viņiem biežāk sastop diabētu, leikozes, vēzi, infertilitāti abiem dzimumiem. Dzīves ilgums var pārsniegt 27 gadus
, bet var attīstīties normāli. Šaura seja, izteikts deguns. Sejas krāsa sārta, cieš no saules stariem, kuru ietekmē rodas raksturīgi izsitumi uz plakstiņiem, vaigiem, lūpām, ap degunu. Indivīdi cieš no elpošanas ceļu, ausu infekcijām. Viņiem biežāk sastop diabētu, leikozes, vēzi, infertilitāti abiem dzimumiem. Dzīves ilgums var pārsniegt 27 gadus.")
25
Telomēru fizioloģija

26
Audzējos telomēru garums var variēt. Telomēri ir īsāki, bet permenentā telomerāzes aktivitāte nodrošina audzēju sūnu nemirstību

27
Telomerāze EC. 2.7.7.49 Telomerāze ir reversā transkriptāze, kas kontrolē telomēru sintēzi, pievienojot hromosomām telomērus. Reversā transkriptāze - telomerāze satur savu RNS matricu un ir RNS-atkarīgā DNS polimerāze. Telomerāzes darbības mehānisma pamatā ir DNS pagarināšana, izmantojot tās sastāvā esošo RNS kā matricu un veidojot atbilstošo komplementāro DNS

28
Abas subvienības kodē atšķirīgi gēni. TERT gēna kodējošais rajons ir 3396 bp, un to translē 1131 aminoskābju garā proteīnā. Polipeptīds savienojas ar TERC (451 nukleotīdu), kurš translēts netiek un paliek kā RNS. TERT ir dūrainim līdzīga telpiskā struktūra un tā atļauj aptīties ap hromosomu un pievienot vienpavediena telomēriskos atkārtojumus. TERT ir reversā transkriptāze, kurs intezē DNS uz RNS matricas. TERT ir cieši saistīta ar savu RNS matric, kuru izmanto reversajai transkripcijai t.i. telomēru sintēzei. Telomerāzes uzbūve: 2 subvienības – 1)telomerāzes reversā transkriptāze – TERT-1131 aminoskābju atlieku 2)451 nukleotīdus gara RNS – TERC (saīsina arī hTR)
, kurš translēts netiek un paliek kā RNS. TERT ir dūrainim līdzīga telpiskā struktūra un tā atļauj aptīties ap hromosomu un pievienot vienpavediena telomēriskos atkārtojumus. TERT ir reversā transkriptāze, kurs intezē DNS uz RNS matricas. TERT ir cieši saistīta ar savu RNS matric, kuru izmanto reversajai transkripcijai t.i. telomēru sintēzei. Telomerāzes uzbūve: 2 subvienības – 1)telomerāzes reversā transkriptāze – TERT-1131 aminoskābju atlieku 2)451 nukleotīdus gara RNS – TERC (saīsina arī hTR).")
29
Telomerāze Masspektrometrijas analīzes rezultāti: Cilvēka telomerāze sastāv no divām TERT molekulām, divām TERC molekulām un divām diskerīna (dyskerin) molekulām TERT ir polipeptīds (1131 aa), kurš salocās ar TERC (451 nt garu) RNS molekulu, kura netiek translēta
 molekulām TERT ir polipeptīds (1131 aa), kurš salocās ar TERC (451 nt garu) RNS molekulu, kura netiek translēta")
30
TERT gēns Gēns (TERT): 5. hromosomā, 40.09 Kb, 16 eksoni; Ekspresijas regulācija: Inducējas ar MYC Inhibēts ar MAD1L1 Supresēts ar MEN1, SIP1 Promotera rajona DNS metilēšana, FRK, USF2

32
Proteīns (telomerāzes apgriezta transkriptāze): 127 kDa, 1132 aa Telomerāzes kompleksa komponents, saistās ar NOLA1, NOLA2, NOLA3, NOLA4 proteīniem (nucleolar protein family A) Savstarpēja mijiedarbība: TERC, netieši saistās ar PINX1 kodoliņā TERT gēna produkts un tā funkcijas
: 127 kDa, 1132 aa Telomerāzes kompleksa komponents, saistās ar NOLA1, NOLA2, NOLA3, NOLA4 proteīniem (nucleolar protein family A) Savstarpēja mijiedarbība: TERC, netieši saistās ar PINX1 kodoliņā TERT gēna produkts un tā funkcijas")
35
TERC gēns un tā produkts - RNS Gēns: 3. hromosomā, 1.2 Kb, 1 eksons RNS: 1000 bp, viens transkripts Funkcijas: telomerāzes RNS komponents; piedalās telomēru struktūras saglabāšanā; būtiska loma šūnu nobriešanā un imortalitātes nodrošināšanā Telomerāzes kompleksa komponents, iesaistās ar NOLA2, NOLA3, DKC1 Savstarpēja mijiedarbība:TERT; saistās ar DNS un RNS Pieci literatūrā aprakstīti polimorfismi TERC gēnā

36
Ar telomēriem saistītos proteīnus mēģina dalīt grupās: 3. Telomēru proteīni, kuri saistīti ar DNS reparāciju: ATM (Ataxia teleangectasia mutated) u.c. 4. Telomerāzes komplekss: a) TERC TERT b) RNS stabilizējošie faktori DKS – diskerīns NOLA – nucleolar protein family A... HEST 1 A – human ever shorter telomeres c) heterogēni ar telomerāzi asociēti kodola ribonukleoproteīni, hn RNP... d) Šaperoni HSP90A P23 HSP-71-1
 u.c. 4. Telomerāzes komplekss: a) TERC TERT b) RNS stabilizējošie faktori DKS – diskerīns NOLA – nucleolar protein family A... HEST 1 A – human ever shorter telomeres c) heterogēni ar telomerāzi asociēti kodola ribonukleoproteīni, hn RNP... d) Šaperoni HSP90A P23 HSP")
37
Diskerīns (dyskerin): H/ACA ribonucleoprotein complex subunit 4 Funkcijas: daļa no H/ACA (small nucleolar ribonucleoprotein (H/ACA snoRNP) kompleksa, kas sastāv no NOLA2/NHP2, NOLA1/GAR1, NOLA3/NOP10 un DKC1/NOLA4 (iespējams katalītiskā subvienība); augstāk minētais komplekss saistās ar TERC, kas satur 3’- terminālo domēnu līdzīgu H/ACA snoRNAs boksam. DKC1 specifiskā mijiedarbība ar snoRNAs vai TERC tiek veidota ar NOLA1 un NOLA2 palīdzību Biežākie polimorfismi DKC1 gēnā (7 no 43 aprakstītiem) Gēns: DKC1, Xq28, 15 eksoni, 16 Kb. Proteīns: 514 aa; 58 kDa
 Gēns: DKC1, Xq28, 15 eksoni, 16 Kb. Proteīns: 514 aa; 58 kDa.")
38
Telomerāzes darbības mehānisms

40
Zīdītāju telomeru komplekss

41
Telomerāze un... Katru gadu telomēru pētījumos parādās jaunas detaļas

42
TERT gēna analīzes metodes 1.RNS izdalīšana no perifērām asinīm 2.kDNS iegūšana ar RT-PCR metodi 3.Alternatīvā splaisinga pētīšana ar specifiskiem praimeriem 4.Iegūtos PCR produktus klonē vektorā, lai pārbaudītu sekvenci 5.Sekvenēšana

43
hTERT gēna mutācijas ietekme uz TERT subvienības sekundāro struktūru A – attēlots hTERT konservatīvie domēni ar divām “missensa” mutācijām B – attēlota telomerāzes TERT subvienības sekundārā struktūra un ar zvaigznīti iezīmēta mutācija

44
Telomerāzes aktivitātes noteikšanas princips TRAP (Telomeric repeat amplification protocol) Metode pamatojas uz fluorescējošu praimeru un Q-PCR izmantošanu. 1.Šūnu lizāta sagatavošana TRAP analīzei 2.TRAP analīze –V–Veselu šūnu uz gēlu balstītais fluorescentais (Cy5) TRAP –R–Reālā laika jeb Q-TRAP 3.Relatīvās telomerāzes aktivitātes (RTA) noteikšana –G–Gēlu apstrādā ar Phosphor Imager programmu, kas spēj atpazīt Cy5 fluorescenci –N–Nosaka telomerāzes produkta un 36-bp IC joslu intensitāti ar ImageQuant programmu (Molecular Dynamics) –A–Aprēķina RTA kā telomerāzes produkta intensitātes attiecību pret IC (internal control)
 TRAP –R–Reālā laika jeb Q-TRAP 3.Relatīvās telomerāzes aktivitātes (RTA) noteikšana –G–Gēlu apstrādā ar Phosphor Imager programmu, kas spēj atpazīt Cy5 fluorescenci –N–Nosaka telomerāzes produkta un 36-bp IC joslu intensitāti ar ImageQuant programmu (Molecular Dynamics) –A–Aprēķina RTA kā telomerāzes produkta intensitātes attiecību pret IC (internal control).")
45
Telomerāzes aktivitātes noteikšana ar Cy5 – TRAP (telomēri=TTAGGG) a – cilvēka audzēja šūnu lizāts (MCF-7 krūts karcinomas šūnas) 3-5 ; LB (lizējošais buferis) 6 ; ar RNāzi apstrādāts 1 un termiski inaktivēts 2 šūnu lizāts (negatīvās kontroles); IC – 36-bp iekšējā kontrole b – RTA atkarība no eksperimentā izmantojamā šūnu skaita c – Cy5-TRAP analīze, izmantojot dažādu šūnu skaitu un kultūras: BJ – normāli fibroblasti; MCF-7 (endogēnais telomerāzes aktivātors); HMS+hTERT – cilvēka saistaudu šūnas (eksogēnais telomerāzes aktivātors). 1 2 3 4 5 6
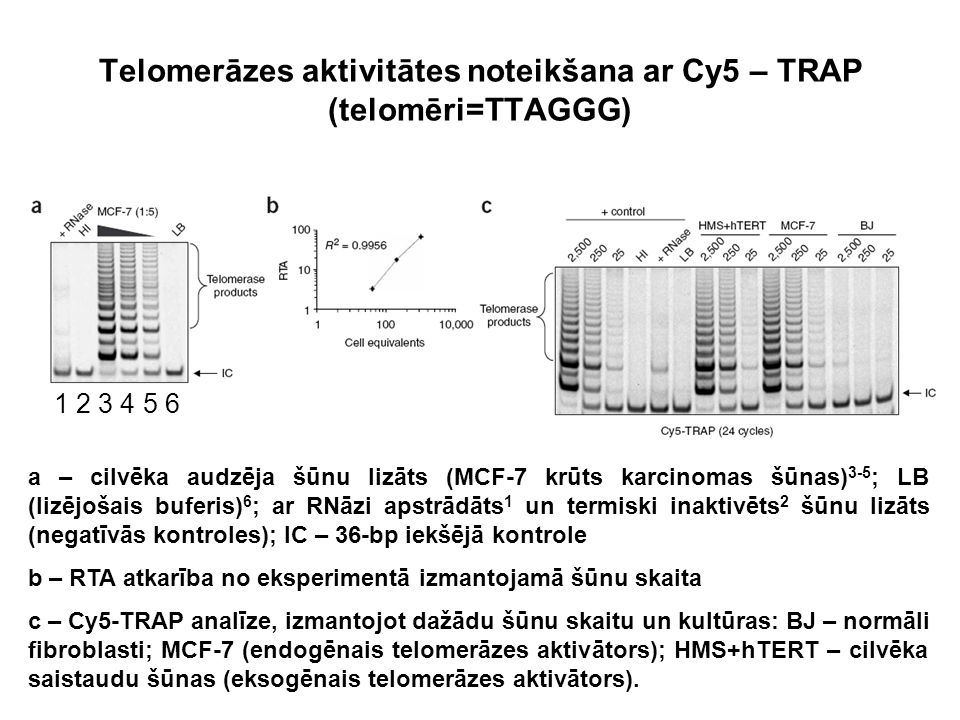
46
Telomerāzes aktivitāte atkarībā no mutācijas vietas D - telomerāzes aktivitātes noteikšana atkarībā no mutācijām hTERT gēnā E – RT-PCR

47
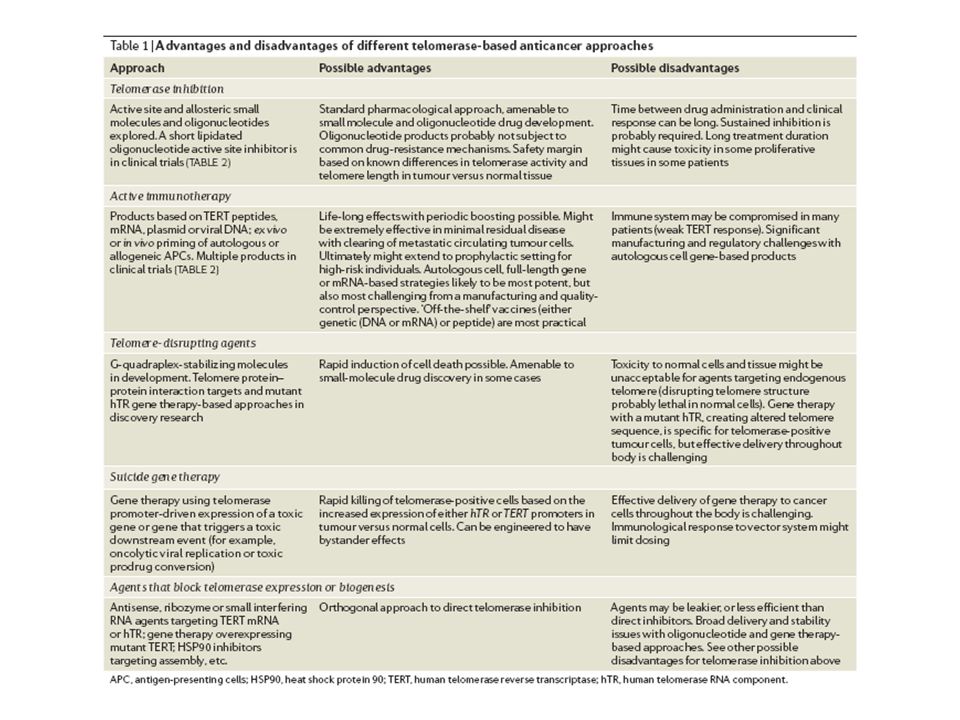
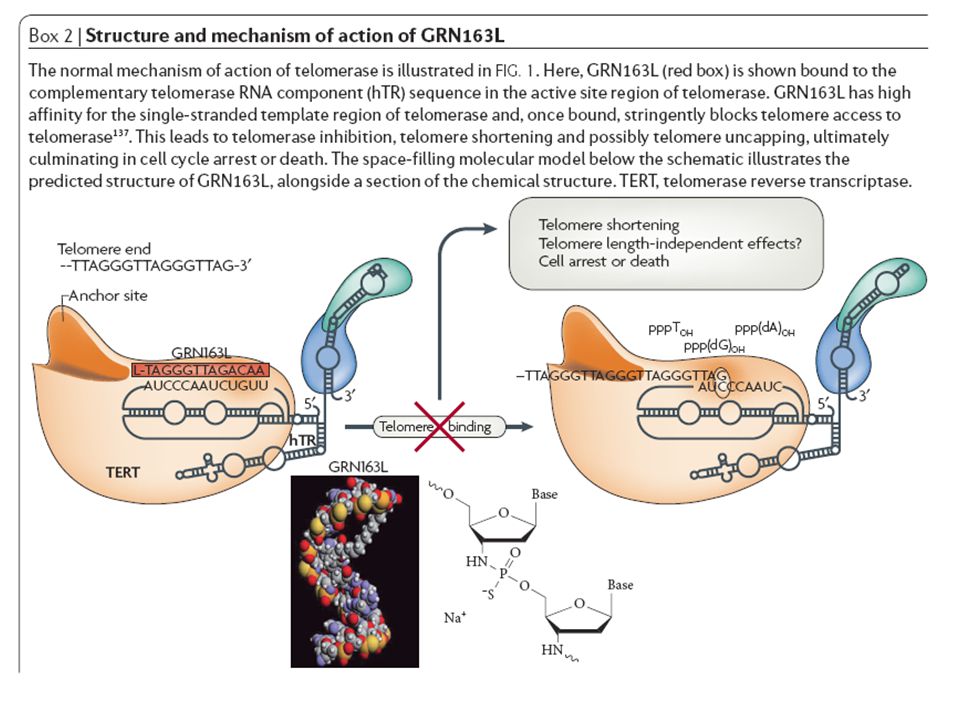
Telomerāzes aktivitātes regulēšanai tiek meklēti farmakopreparāti: 85-90% gadījumos šodien tie ir telomerāzes inhibitori, kuri paredzēti ļaundabīgo audzēju terapijai anti-informācijas oligonukleotidi (antisens oligos) peptido-oligonukleotidi 2-O-metīl - tRNS kvadrupleksa DNS fragmenti
 peptido-oligonukleotidi 2-O-metīl - tRNS kvadrupleksa DNS fragmenti")
48
Telomerāzes aktivitātes regulācijas iespējas

Παρόμοιες παρουσιάσεις









, καθεμία από τις οποίες σχηματίζει.>")



>")








