Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Μεταφορά μικρών μορίων διαμέσου κυτταρικών μεμβρανών
Φυσιολογία Μεμβρανών 2009 Μεταφορά μικρών μορίων διαμέσου κυτταρικών μεμβρανών ΜΕΡΟΣ III: Δομή και Φυσιολογική Λειτουργία των Μεμβρανικών Μεταφορέων Ε. Κωλέττας Μονάδα Κυτταρικής και Μοριακής Φυσιολογίας Εργαστήριο Φυσιολογίας Ιατρική Σχολή Πανεπιστήμιο Ιωαννίνων

2
Στόχοι Λειτουργία και δομή και οργάνωση των διαύλων (channels)
Ορισμός διαμόρφωσης/ενεργοποίησης πύλης διαύλου ( channel “gating”) Κατανόηση των διαφορών στα χαρακτηριστικά μεταφοράς των διαύλων, και μεταξύ πρωτογενών και δευτερογενών ενεργών μεταφορέων. Ορισμός και περιγραφή μιας αντλίας (pump) ή ενός πρωτογενούς συστήματος ενεργής μεταφοράς. Κατανόηση του ρόλου των ATPασών τύπου-P [P-type ATPases] [ή αντλίων ιόντων (ion pumps)] συμπεριλαμβανομένων των: ATPάση Na/K, ATPάση Ca, ATPάση H/K και ATPάση Cu (Na/K-ATPase, Ca-ATPase, H/K-ATPase, and Cu-ATPases). Κατανόηση της ενεργητικής βάσης της δευτερογενούς ενεργητικής μεταφοράς και το ρόλο των συστημάτων δευτεροταγούς ενεργητικής μεταφοράς σε κυτταρικά συστήματα.
 Κατανόηση των διαφορών στα χαρακτηριστικά μεταφοράς των διαύλων, και μεταξύ πρωτογενών και δευτερογενών ενεργών μεταφορέων. Ορισμός και περιγραφή μιας αντλίας (pump) ή ενός πρωτογενούς συστήματος ενεργής μεταφοράς. Κατανόηση του ρόλου των ATPασών τύπου-P [P-type ATPases] [ή αντλίων ιόντων (ion pumps)] συμπεριλαμβανομένων των: ATPάση Na/K, ATPάση Ca, ATPάση H/K και ATPάση Cu (Na/K-ATPase, Ca-ATPase, H/K-ATPase, and Cu-ATPases). Κατανόηση της ενεργητικής βάσης της δευτερογενούς ενεργητικής μεταφοράς και το ρόλο των συστημάτων δευτεροταγούς ενεργητικής μεταφοράς σε κυτταρικά συστήματα.")
3
ΔΙΑΥΛΟΙ

4
Δίαυλοι Βασικοί για την γρήγορη διάδοση διαφόρων σημάτων/μηνυμάτων και τη συστολή των μυών. Δυσλειτουργία των διαύλων οδηγεί σε μυοτονία, περιοδική παράλυση, καρδιακή αρρυθμία, ορισμένες μορφές επιληψίας και άλλες ανωμαλίες.

5
Διαδικασίες που εμπλέκονται οι Πρωτεϊνικοί Δίαυλοι
Οι δίαυλοι είναι ενσωματωμένες μεμβρανικές πρωτεΐνες που εμπλέκονται σε μια σειρά φυσιολογικές διαδικασίες: Μεταφορά ιόντων και νερού Ρύθμιση του μεμβρανικού δυναμικού Έκκριση ορμονών Έκκριση νευροδιαβιβαστών Μεταγωγή σήματος Νευρικές ώσεις Μυϊκή συστολή Καρδιακός κτύπος Όραση και Όσφρηση Πόνος Δυσλειτουργία των διαύλων μπορεί να οδηγήσει σε: • Επιληψία • Απώλεια ακοής • Απώλεια όρασης • Ημικρανίες • Καρδιακή αρρυθμία • Μυοτονία • Απώλεια μνήμης

6
Πρωτεϊνικοί Δίαυλοι Οι δίαυλοι ταξινομούνται σε δύο κύριες κατηγορίες:
Η αγωγιμότητα των διαύλων ιόντων συνήθως ρυθμίζεται από τη μετατροπή της διαμόρφωσης της δομής της πρωτεΐνης του διαύλου μεταξύ κυρίως δύο καταστάσεων: ‘ανοικτής’ και ‘κλειστής’. (conformational switching of the protein structure between “open” and “close” states). Μερικοί δίαυλοι διαμορφώνουν επιπλέον και μια ‘ανενεργό’ κατάσταση (inactive state). Οι δίαυλοι ταξινομούνται σε δύο κύριες κατηγορίες: I. Δίαυλοι χωρίς πύλη (Non-gated channels) ΙΙ. Δίαυλοι με πύλη (Gated channels)
. Μερικοί δίαυλοι διαμορφώνουν επιπλέον και μια ‘ανενεργό’ κατάσταση (inactive state). Οι δίαυλοι ταξινομούνται σε δύο κύριες κατηγορίες: I. Δίαυλοι χωρίς πύλη (Non-gated channels) ΙΙ. Δίαυλοι με πύλη (Gated channels)")
7
I. Δίαυλοι χωρίς πύλη (Non-gated channels)
Οι σημαντικότεροι δίαυλοι χωρίς πύλη είναι οι υδατοπορίνες (aquaporines) [Peter Agre, Nobel Χημείας 2003 (με Roderick MacKinnon για διαύλους K+)]. Οι υδατοπορίνες είναι ενσωματωμένες μεμβρανικές πρωτεΐνες που σχηματίζουν πόρους στην κυτταρική μεμβράνη και επιτρέπουν τη διάχυση μορίων νερού, αναφέρονται και ως δίαυλοι νερού. Το νερό μεταφέρεται με απλή διάχυση στην κατεύθυνση της ωσμωτικής πίεσης. Παίζουν σημαντικό ρόλο στη μεταφορά νερού στα ερυθροκύτταρα και νεφρικά κύτταρα (proximal renal tubule cells). Μολονότι οι υδατοπορίνες θεωρούνται ανοικτοί δίαυλοι δεν επιτρέπουν τη διάχυση ιόντων ή άλλων μορίων (εκτός από συγκεκριμένες κατηγορίες τις υδατο-γλυκεροπορίνες). Αυτό οφείλεται στη δομή τους που σχηματίζει ένα ηθμός (φίλτρο) επιλεκτικότητας (selectivity filter). Ένα μόριο (μονομερές) υδατοπορίνης αποτελείται από 6 α-έλικες που συνδέονται με βρόγχους. Οι βρόγχοι Β και Ε είναι υδρόφοβοι και φέρουν την αλληλουχία Asp-Pro-Ala (NPA) που εμπλέκονται στο σχηματισμό του διαύλου νερού. Οι δίαυλοι σχηματίζονται από 4 μονομερή υδατοπορινών.
 [Peter Agre, Nobel Χημείας 2003 (με Roderick MacKinnon για διαύλους K+)]. Οι υδατοπορίνες είναι ενσωματωμένες μεμβρανικές πρωτεΐνες που σχηματίζουν πόρους στην κυτταρική μεμβράνη και επιτρέπουν τη διάχυση μορίων νερού, αναφέρονται και ως δίαυλοι νερού. Το νερό μεταφέρεται με απλή διάχυση στην κατεύθυνση της ωσμωτικής πίεσης. Παίζουν σημαντικό ρόλο στη μεταφορά νερού στα ερυθροκύτταρα και νεφρικά κύτταρα (proximal renal tubule cells). Μολονότι οι υδατοπορίνες θεωρούνται ανοικτοί δίαυλοι δεν επιτρέπουν τη διάχυση ιόντων ή άλλων μορίων (εκτός από συγκεκριμένες κατηγορίες τις υδατο-γλυκεροπορίνες). Αυτό οφείλεται στη δομή τους που σχηματίζει ένα ηθμός (φίλτρο) επιλεκτικότητας (selectivity filter). Ένα μόριο (μονομερές) υδατοπορίνης αποτελείται από 6 α-έλικες που συνδέονται με βρόγχους. Οι βρόγχοι Β και Ε είναι υδρόφοβοι και φέρουν την αλληλουχία Asp-Pro-Ala (NPA) που εμπλέκονται στο σχηματισμό του διαύλου νερού. Οι δίαυλοι σχηματίζονται από 4 μονομερή υδατοπορινών.")
8
ΙΙ. Δίαυλοι με πύλη (Gated channels)
Η διαδικασία του ανοίγματος-κλεισίματος της πύλης του διαύλου εξαρτάται από τον τύπο και δομή του διαύλου και του ερεθίσματος: Δίαυλοι που εξαρτώνται από το ηλεκτρικό δυναμικό ή Δίαυλοι δυναμικού ή Τασεο-εξαρτώμενοι δίαυλοι (Voltage-gated channels): Δίαυλοι ιόντων K+, Na+, Ca2+ - Μεταβολές στο ηλεκτρικό δυναμικό κατά πλάτος των μεμβρανών Δίαυλοι που εξαρτώνται από προσδέτη ή Χημειο-εξαρτώμενοι δίαυλοι(Ligand-gated channels): Ακετυλοχολίνης, γλουταμινικού οξέος, GABA – Δέσμευση ενός προσδέτη (Ligand binding) Μηχανο-ευαίσθητοι Δίαυλοι (Mechanosensitive channels) Μηχανικές δυνάμεις δρουν σε κυτταρικά συστατικά
: Δίαυλοι ιόντων K+, Na+, Ca2+ - Μεταβολές στο ηλεκτρικό. δυναμικό κατά πλάτος των μεμβρανών. Δίαυλοι που εξαρτώνται από προσδέτη ή Χημειο-εξαρτώμενοι. δίαυλοι(Ligand-gated channels): Ακετυλοχολίνης, γλουταμινικού οξέος, GABA – Δέσμευση ενός προσδέτη (Ligand binding) Μηχανο-ευαίσθητοι Δίαυλοι (Mechanosensitive channels) Μηχανικές δυνάμεις δρουν σε κυτταρικά συστατικά.")
9
I. Δίαυλοι που ενεργοποιούνται από ηλεκτρικό δυναμικό ή δίαυλος δυναμικού (Voltage -gated channels)
● Παιζουν ςημαντικο ρολο ςτη νευροδιαβιβαςη και ςτη μυϊκη ςυςτολη. ● Είναι πολύ επιλεκτικοί σε σχέση με το ιόν που μεταφέρουν (ιοντο-εξειδικευμένοι δίαυλοι), σε αντίθεση με τους διαύλους που ενεργοποιούνται από έναν προσδέτη (ligand-gated channels) (για παράδειγμα ο υποδοχέας της ακετυλοχολίνης είναι ισοδύναμα διαπερατός στα ιόντα Na+ και K+, ενώ ο δίαυλος ιόντων καλίου είναι 100 φορές περισσότερο διαπερατός στα ιόντα K+ από ότι στα ιόντα Na+). ● Πολλοί δίαυλοι έχουν συγκεκριμένα φαρμακολογικά χαρακτηριστικά και αποτελούν στόχους διαφόρων τοξινών [τετραδοτοξίνης (tetrodotoxin), Σαξιτοξίνης (saxitoxin), Βερατριδίνης (veratridin)]. ● Παραμένουν ανοικτοί για ένα σύντομο χρονικό διάστημα και στη συνέχεια διαμορφώνουν μια ‘ανενεργό’ μορφή.
, σε αντίθεση με τους διαύλους που ενεργοποιούνται από έναν προσδέτη (ligand-gated channels) (για παράδειγμα ο υποδοχέας της ακετυλοχολίνης είναι ισοδύναμα διαπερατός στα ιόντα Na+ και K+, ενώ ο δίαυλος ιόντων καλίου είναι 100 φορές περισσότερο διαπερατός στα ιόντα K+ από ότι στα ιόντα Na+). ● Πολλοί δίαυλοι έχουν συγκεκριμένα φαρμακολογικά χαρακτηριστικά και αποτελούν στόχους διαφόρων τοξινών [τετραδοτοξίνης (tetrodotoxin), Σαξιτοξίνης (saxitoxin), Βερατριδίνης (veratridin)]. ● Παραμένουν ανοικτοί για ένα σύντομο χρονικό διάστημα και στη συνέχεια διαμορφώνουν μια ‘ανενεργό’ μορφή.")
10
ΔΥΝΑΜΙΚΟ ΜΕΜΒΡΑΝΗΣ Μεμβρανικό δυναμικό (membrane potential) ή ηλεκτροχημικό δυναμικό (electrochemical potential) ορίζεται η διαφορά δυναμικού μεταξύ του εσωτερικού και του εξωτερικού ενός κυττάρου. Το μεμβρανικό δυναμικό όλων των κυττάρων οφείλεται στη διαφορά των συγκεντρώσεων των ιόντων (K+, Na+, Cl-, Ca2+) μεταξύ της ενδοκυττάριας και εξωκυττάριας πλευράς της κυτταρικής μεμβράνης. Τα σημαντικότερα ιόντα που συμβάλλουν στο μεμβρανικό δυναμικό είναι K+ και Na+
 ή ηλεκτροχημικό δυναμικό (electrochemical potential) ορίζεται η διαφορά δυναμικού μεταξύ του εσωτερικού και του εξωτερικού ενός κυττάρου. Το μεμβρανικό δυναμικό όλων των κυττάρων οφείλεται στη διαφορά των συγκεντρώσεων των ιόντων (K+, Na+, Cl-, Ca2+) μεταξύ της ενδοκυττάριας και εξωκυττάριας πλευράς της κυτταρικής μεμβράνης. Τα σημαντικότερα ιόντα που συμβάλλουν στο μεμβρανικό δυναμικό είναι K+ και Na+")
11
ΡΟΛΟΣ ΤΟΥ μεμβρανικΟΥ δυναμικΟΥ
Το μεμβρανικό δυναμικό μη-διεγέρσιμων (non-excitable) και διεγέρσιμων (ecitable) κυττάρων έχει δύο βασικές λειτουργίες: (i) Επιτρέπει στο κύτταρο να λειτουργεί ως μια μπαταρία που παρέχει ενέργεια για τη δράση κυρίως των ενσωματωμένων πρωτεϊνών στην κυτταρική μεμβράνη. ii) Σε ηλεκτρικά διεγέρσιμα κύτταρα όπως τα μυϊκά και νευρικά κύτταρα, το μεμβρανικό δυναμικό χρησιμοποιείται για τη μετάδοση σημάτων κατά μήκος της μεμβράνης. Οι κυτταρικές μεμβράνες των διεγέρσιμων κυττάρων έχουν την ικανότητα να παράγουν ηλεκτροχημικές ώσεις, οι οποίες μπορούν να μεταδίδουν σήματα, όπως τα νευρικά σήματα, κατά μήκος της μεμβράνης. Το άνοιγμα ή το κλείσιμο των διαύλων ιόντων σε οποιοδήποτε σημείο της κυτταρικής μεμβράνης οδηγεί σε τοπικές μεταβολές του μεμβρανικού δυναμικού προκαλώντας την αύξηση και τη ροή της ηλεκτρικής δραστηριότητας σε άλλα σημεία της μεμβράνης. Επομένως, το μεμβρανικό δυναμικό χαρακτηρίζεται σε: (α) Δυναμικό Ηρεμίας (Resting membrane potential) (β) Δυναμικό ενέργειας (Action potential)
 και διεγέρσιμων (ecitable) κυττάρων έχει δύο βασικές λειτουργίες: (i) Επιτρέπει στο κύτταρο να λειτουργεί ως μια μπαταρία που παρέχει ενέργεια για τη δράση κυρίως των ενσωματωμένων πρωτεϊνών στην κυτταρική μεμβράνη. ii) Σε ηλεκτρικά διεγέρσιμα κύτταρα όπως τα μυϊκά και νευρικά κύτταρα, το μεμβρανικό δυναμικό χρησιμοποιείται για τη μετάδοση σημάτων κατά μήκος της μεμβράνης. Οι κυτταρικές μεμβράνες των διεγέρσιμων κυττάρων έχουν την ικανότητα να παράγουν ηλεκτροχημικές ώσεις, οι οποίες μπορούν να μεταδίδουν σήματα, όπως τα νευρικά σήματα, κατά μήκος της μεμβράνης. Το άνοιγμα ή το κλείσιμο των διαύλων ιόντων σε οποιοδήποτε σημείο της κυτταρικής μεμβράνης οδηγεί σε τοπικές μεταβολές του μεμβρανικού δυναμικού προκαλώντας την αύξηση και τη ροή της ηλεκτρικής δραστηριότητας σε άλλα σημεία της μεμβράνης. Επομένως, το μεμβρανικό δυναμικό χαρακτηρίζεται σε: (α) Δυναμικό Ηρεμίας (Resting membrane potential) (β) Δυναμικό ενέργειας (Action potential)")
12
(α) ΔυναμιΚΟ ΗρεμΙας Το δυναμικό ηρεμίας μη-διεγέρσιμων και διεγέρσιμων κυττάρων ορίζεται ως η βασική, σταθερή ηλεκτρική δραστηριότητα που ανιχνεύεται στη κυτταρική μεμβράνη. Το δυναμικό ηρεμίας των περισσοτέρων, δηλ. των μη-διεγέρσιμων κυττάρων είναι ~ -40 mV (-40 εως -60 mV), ενώ των διεγέρσιμων κυττάρων (μυϊκών, νευρικών και ορισμένων εκκριτικών κυττάρων αδένων) είναι ~ -60 mV (-60 εως -80 mV). Το δυναμικό ηρεμίας οφείλεται στη μη-ισοδύναμη κατανομή κυρίως των ιόντων K+ και Na+ μέσα και έξω από το κύτταρο. Το δυναμικό ηρεμίας δεν επιτρέπει τη καθαρή διάχυση ιόντος προς οποιαδήποτε κατεύθυνση και δίνεται από την εξίσωση του Nernst.
, ενώ των διεγέρσιμων κυττάρων (μυϊκών, νευρικών και ορισμένων εκκριτικών κυττάρων αδένων) είναι ~ -60 mV (-60 εως -80 mV). Το δυναμικό ηρεμίας οφείλεται στη μη-ισοδύναμη κατανομή κυρίως των ιόντων K+ και Na+ μέσα και έξω από το κύτταρο. Το δυναμικό ηρεμίας δεν επιτρέπει τη καθαρή διάχυση ιόντος προς οποιαδήποτε κατεύθυνση και δίνεται από την εξίσωση του Nernst.")
13
Ιόν mM Ενδοκυττάρια Εξωκυττάρια Λόγος (Εντός/εκτός)
Η μη-ισοδύναμη κατανομή ιόντων σε ένα κύτταρο δημιουργεί ένα ‘δυναμικό ηρεμίας’ (“resting potential”) Ιόν mM Ενδοκυττάρια Εξωκυττάρια Λόγος (Εντός/εκτός) Na ~ 0.1 K ~ 30 Mg Ca H x 10-5 (pH 7.2) 4 x 10-5 (pH 7.4) Cl DY = RT ln Ionin= -60 mV ZF Ionout Ηλεκτρικό Χημικό δυναμικό Σ’ αυτό το δυναμικό το ιόν K+ είναι πιο κοντά σε ισορροπία από ότι το ιόν Na+ (ΔΨK = -75 mV, ενώ ΔΨNa = +55 mV ). Επομένως, αν η μεμβράνη γίνονταν πλήρως διαπερατή στα ιόντα, τότε το κύριο αποτέλεσμα θα ήταν η μαζική εισροή ιόντων Na+. (Σημείωση: Λαμβάνοντας υπόψη το πλάτος της κυτταρικής μεμβράνης ~3.5 nm, η διαφορά του μεμβρανικού δυναμικού αντιστοιχεί σε: 0.06 V ανά 3.5 nm ή ~ V ανά cm). R (gas const) =8.28 J/degree.mol T=293 K F(Faraday const)=96 C/mol.V Z (valency) ΔΨ= ηλεκτρεγερτική δύναμη Εξίσωση του Nernst (Ηλεκτρο- χημικής Ισορροπίας)
 Ιόν mM Ενδοκυττάρια Εξωκυττάρια Λόγος (Εντός/εκτός) Na ~ 0.1. K ~ 30. Mg Ca H+ 7 x 10-5 (pH 7.2) 4 x 10-5 (pH 7.4) Cl DY = - RT ln Ionin= -60 mV. ZF Ionout. Ηλεκτρικό Χημικό δυναμικό. Σ’ αυτό το δυναμικό το ιόν K+ είναι πιο κοντά σε ισορροπία από ότι το ιόν Na+ (ΔΨK = -75 mV, ενώ ΔΨNa = +55 mV ). Επομένως, αν η μεμβράνη γίνονταν πλήρως διαπερατή στα ιόντα, τότε το κύριο αποτέλεσμα θα ήταν η μαζική εισροή ιόντων Na+. (Σημείωση: Λαμβάνοντας υπόψη το πλάτος της κυτταρικής μεμβράνης ~3.5 nm, η διαφορά του μεμβρανικού δυναμικού αντιστοιχεί σε: 0.06 V ανά 3.5 nm ή ~ V ανά cm). R (gas const) =8.28 J/degree.mol. T=293 K. F(Faraday const)=96 C/mol.V. Z (valency) ΔΨ= ηλεκτρεγερτική δύναμη. Εξίσωση του. Nernst. (Ηλεκτρο- χημικής. Ισορροπίας)")
14
Πως διατηρείται το Δυναμικό Ηρεμίας των Μεμβρανών
Πως δημιουργείται η μη-ισοδύναμη κατανομή των ιόντων K+ και Na+ στις δύο πλευρές της μεμβράνης; Η μη-ισοδύναμη κατανομή των ιόντων K+ και Na+ στις δύο πλευρές της μεμβράνης οφείλεται στην αντλία ιόντων K+/Na+ (K+/Na+ Pump) που μεταφέρει ενεργά ιόντα Na+ από μέσα προς τα έξω, και ιόντα K+ απ’ έξω προς τα μέσα, με τη δαπάνη ενέργειας υπό τη μορφή ATP .
 που μεταφέρει ενεργά ιόντα Na+ από μέσα προς τα έξω, και ιόντα K+ απ’ έξω προς τα μέσα, με τη δαπάνη ενέργειας υπό τη μορφή ATP .")
15
η μεμβράνη φέρει επίσης διαύλους ιόντων K+ και Na+ από τους οποίους διαχέονται αυτά τα ιόντα. Σε ένα κύτταρο σε κατάσταση ηρεμίας, οι δίαυλοι Na+ είναι κλειστοί ενώ οι δίαυλοι K+ είναι συνήθως ανοικτοί. Σαν αποτέλεσμα, τα ιόντα Na+ δεν διαχέονται διαμέσου της μεμβράνης και παραμένουν έξω, ενώ τα ιόντα K+ μπορούν να διαχυθούν από τους διαύλους τους. Επειδή η αντλία Na-K αντλεί 3 ιόντα Na+ από μέσα προς τα έξω και 2 ιόντα K+ απ’ έξω προς τα μέσα το αποτέλεσμα είναι η αύξηση της συγκέντρωσης των Na+ στο εξωτερικό και των Κ+ στο εσωτερικό του κυττάρου. Επειδή οι δίαυλοι Na+ είναι κλειστοί και οι δίαυλοι K+ επιτρέπουν την ελεύθερη διάχυση των ιόντων K+, τότε καθώς αυξάνεται η συγκέντρωση τους στο εσωτερικό τα ιόντα K+ θα διαχέονται προς το εξωτερικό του κυττάρου προς τη διαβάθμιση της συγκέντρωσης τους (για να διατηρηθεί η χημική ισορροπία). Το αποτέλεσμα αυτής της διάχυσης των ιόντων K+ θα οδηγήσει σε ένα πιο αρνητικό φορτίο στο εσωτερικό του κυττάρου και αυτή η ηλεκτρική διαφορά μέσα και έξω από το κύτταρο θα οδηγήσει στην διάχυση των ιόντων K+ μέσα στο κύτταρο (τα K+ ελκύονται από το αρνητικό φορτίο στο εσωτερικό). Η καθαρή μετακίνηση (διάχυση) των ιόντων K+ θα σταματήσει όταν επέλθει ισορροπία μεταξύ της χημικής και ηλεκτρική δύναμης (Ηλεκτροχημική ισορροπία). Γενικά, επομένως, υπάρχουν άφθονα θετικά φορτισμένα ιόντα K+ μέσα στο κύτταρο και άφθονα θετικά φορτισμένα ιόντα Na+ και μερικά ιόντα K+ έξω από το κύτταρο. Αυτό σημαίνει ότι υπάρχουν περισσότερα θετικά φορτία στο εξωτερικό απ’ ότι στο εσωτερικό του κυττάρου. Επομένως υπάρχει μια μη-ισοδύναμη κατανομή ιόντων στις δύο πλευρές τις μεμβράνης που οδηγεί σε μια αρνητική διαφορά δυναμικού της τάξης των -60 mV που ονομάζεται δυναμικό ηρεμίας. Αυτό το δυναμικό διατηρείται μέχρι να διαταραχθεί ή να διεγερθεί η κυτταρική μεμβράνη. ( 44
. Το αποτέλεσμα αυτής της διάχυσης των ιόντων K+ θα οδηγήσει σε ένα πιο αρνητικό φορτίο στο εσωτερικό του κυττάρου και αυτή η ηλεκτρική διαφορά μέσα και έξω από το κύτταρο θα οδηγήσει στην διάχυση των ιόντων K+ μέσα στο κύτταρο (τα K+ ελκύονται από το αρνητικό φορτίο στο εσωτερικό). Η καθαρή μετακίνηση (διάχυση) των ιόντων K+ θα σταματήσει όταν επέλθει ισορροπία μεταξύ της χημικής και ηλεκτρική δύναμης (Ηλεκτροχημική ισορροπία). Γενικά, επομένως, υπάρχουν άφθονα θετικά φορτισμένα ιόντα K+ μέσα στο κύτταρο και άφθονα θετικά φορτισμένα ιόντα Na+ και μερικά ιόντα K+ έξω από το κύτταρο. Αυτό σημαίνει ότι υπάρχουν περισσότερα θετικά φορτία στο εξωτερικό απ’ ότι στο εσωτερικό του κυττάρου. Επομένως υπάρχει μια μη-ισοδύναμη κατανομή ιόντων στις δύο πλευρές τις μεμβράνης που οδηγεί σε μια αρνητική διαφορά δυναμικού της τάξης των -60 mV που ονομάζεται δυναμικό ηρεμίας. Αυτό το δυναμικό διατηρείται μέχρι να διαταραχθεί ή να διεγερθεί η κυτταρική μεμβράνη. ( 44.")
16
Πλασματική μεμβράνη ζωικών κυττάρων έχει πολλούς ανοιχτούς διαύλους K+ αλλά ελάχιστους Na+, Cl- ή Ca2+. Όταν ένα κύτταρο βρίσκεται σε ηρεμία η κυτταρική μεμβράνη είναι πιο διαπερατή στα ιόντα K+ από ότι στα ιόντα Na+ (ή Cl-). Επομένως K+ κινούνται από μέσα προς τα έξω, δημιουργώντας ένα –ve φορτίο μέσα και +ve έξω. Αυτή η ροή των K+ από μέσα προς τα έξω διαμέσου των διαύλων των ιόντων K+ είναι υπεύθυνη για το αρνητικό μεμβρανικό δυναμικό στο εσωτερικό του κυττάρου. Άρα, η ηλεκτροχημική ισορροπία που προέρχεται από την κατανομή αυτών των ιόντων στις δυο πλευρές της μεμβράνης, μαζί με τις σχετικές διαπερατότητες αυτών των ιόντων είναι υπεύθυνη για το φορτίο των –60 mV το οποίο ονομάζεται δυναμικό ηρεμίας.
. Επομένως K+ κινούνται από μέσα προς τα έξω, δημιουργώντας ένα –ve φορτίο μέσα και +ve έξω. Αυτή η ροή των K+ από μέσα προς τα έξω διαμέσου των διαύλων των ιόντων K+ είναι υπεύθυνη για το αρνητικό μεμβρανικό δυναμικό στο εσωτερικό του κυττάρου. Άρα, η ηλεκτροχημική ισορροπία που προέρχεται από την κατανομή αυτών των ιόντων στις δυο πλευρές της μεμβράνης, μαζί με τις σχετικές διαπερατότητες αυτών των ιόντων είναι υπεύθυνη για το φορτίο των –60 mV το οποίο ονομάζεται δυναμικό ηρεμίας.")
17
Δυναμικό ενέργειας (Action potential)
Δυναμικό ενέργειας (Action potential) ορίζεται ως η ταχεία μεταβολή του μεμβρανικού δυναμικού όταν διεγείρεται ένα νευρικό ή μυϊκό κύτταρο. Ειδικότερα, το μεμβρανικό δυναμικό μεταβάλλεται από το δυναμικό ηρεμίας (~ -60 mV) σε θετικές τιμές (~ mV) σε ένα σύντομο χρονικό διάστημα (ολίγων ms) Na+ εισέρχεται, K+ εξέρχεται Το δυναμικό ενέργειας παράγεται όταν η μεμβράνη εκπολώνεται (depolarized) τοπικά κατά ~20 mV Για να λάβει χώρα η διαδικασία, θα πρέπει οι δίαυλοι δυναμικού (voltage-gated channels) να: (α) είναι πολύ επιλεκτικοί c) είναι ευαίσθητοι στο ηλεκτρικό δυναμικό (voltage sensitive) (b) είναι πολύ γρήγοροι d) έχουν ένα μηχανισμό γρήγορης απενεργοποίησης
 ορίζεται ως η ταχεία μεταβολή του μεμβρανικού δυναμικού όταν διεγείρεται ένα νευρικό ή μυϊκό κύτταρο. Ειδικότερα, το μεμβρανικό δυναμικό μεταβάλλεται από το δυναμικό ηρεμίας (~ -60 mV) σε θετικές τιμές (~ mV) σε ένα σύντομο χρονικό διάστημα (ολίγων ms) Na+ εισέρχεται, K+ εξέρχεται. Το δυναμικό ενέργειας παράγεται όταν η μεμβράνη εκπολώνεται (depolarized) τοπικά κατά ~20 mV. Για να λάβει χώρα η διαδικασία, θα πρέπει οι δίαυλοι δυναμικού (voltage-gated channels) να: (α) είναι πολύ επιλεκτικοί c) είναι ευαίσθητοι στο ηλεκτρικό δυναμικό (voltage. sensitive) (b) είναι πολύ γρήγοροι d) έχουν ένα μηχανισμό γρήγορης απενεργοποίησης.")
18
Τι προκαλεί τη μεταβολή του δυναμικού;
Το ερέθισμα προκαλεί το σύντομο άνοιγμα των διαύλων Na+, και επειδή η [Na+] είναι μεγαλύτερη στο εξωτερικό από ότι το εσωτερικό του κυττάρου, τα Na+ διαχέονται γρήγορα μέσα στο κύτταρο. Η είσοδος των θετικά φορτισμένων ιόντων Na+ καθιστά το ΜΔ πιο θετικό (μέσα από ότι έξω). Όταν το ΜΔ γίνει θετικότερο κατά ~20 mV του ΔΗ (ουδός πυροδότησης) τότε σηματοδοτείται η έναρξη ενός δυναμικού ενέργειας. Η καθαρή είσοδος ιόντων Na+ αυξάνει το ΜΔ προκαλώντας το άνοιγμα περισσοτέρων τασο- εξαρτώμενων διαύλων Na+ (θετική ανατροφοδότηση) και η μεμβράνη υφίσταται εκπόλωση (Na+in>K+out) και παρατηρείται υπερακόντιση – περίοδος κατά την οποία το ΜΔ είναι πάνω από 0. ΤΟ ΜΒ κορυφώνεται στα ~+40 mV που ορίζεται ως Δυναμικό Ενέργειας. Οι δίαυλοι Na+ κλείνουν. Οι δίαυλοι ιόντων K+ ανοίγουν και επειδή η [K+] είναι μεγαλύτερη στο εσωτερικό από ότι το εξωτερικό του κυττάρου, τα K+ διαχέονται γρήγορα έξω από το κύτταρο. Καθώς τα K+ εξέρχονται, το εσωτερικό της μεμβράνης γίνεται πιο αρνητικό σε σχέση με το εξωτερικό και η μεμβράνη εισέρχεται στη φάση της επαναπόλωσης. Η ταχεία είσοδος των K+ προκαλεί υπακόντιση (θετικό μεταδυναμικό) όπου το ΜΔ γίνεται πιο ηλεκτραρνητικό (~-80 mV) σε σχέση με ΔΗ.
. Όταν το ΜΔ γίνει θετικότερο κατά ~20 mV του ΔΗ. (ουδός πυροδότησης) τότε σηματοδοτείται η έναρξη ενός δυναμικού ενέργειας. Η καθαρή είσοδος ιόντων Na+ αυξάνει το ΜΔ προκαλώντας το άνοιγμα περισσοτέρων τασο- εξαρτώμενων διαύλων Na+ (θετική ανατροφοδότηση) και η μεμβράνη υφίσταται εκπόλωση (Na+in>K+out) και παρατηρείται υπερακόντιση – περίοδος κατά την οποία το ΜΔ είναι πάνω από 0. ΤΟ ΜΒ κορυφώνεται στα ~+40 mV που ορίζεται ως Δυναμικό Ενέργειας. Οι δίαυλοι Na+ κλείνουν. Οι δίαυλοι ιόντων K+ ανοίγουν και επειδή η [K+] είναι μεγαλύτερη στο εσωτερικό από ότι το εξωτερικό του κυττάρου, τα K+ διαχέονται γρήγορα έξω από το κύτταρο. Καθώς τα K+ εξέρχονται, το εσωτερικό της μεμβράνης γίνεται πιο αρνητικό σε σχέση με το εξωτερικό και η μεμβράνη εισέρχεται στη φάση της επαναπόλωσης. Η ταχεία είσοδος των K+ προκαλεί υπακόντιση (θετικό μεταδυναμικό) όπου το ΜΔ γίνεται πιο ηλεκτραρνητικό (~-80 mV) σε σχέση με ΔΗ.")
19
Κατά την περίοδο της υπο-ακόντισης (~1 ms), δεν μπορεί να παραχθεί ένα δεύτερο δυναμικό ενέργειας, καθώς οι δίαυλοι ιόντων Na+ είναι ανενεργοί. Στην πραγματικότητα, οι τασο-εξαρτώμενοι δίαυλοι ιόντων Na+ εναλλάσσονται μεταξύ 3 καταστάσεων: ανοικτοί, κλειστοί και ανενεργοί. Η ηλεκτροχημική ισορροπία αποκαθίσταται κα με τη βοήθεια της αντλίας ιόντων Na+/K+ και το μεμβρανικό δυναμικό επιστρέφει στο δυναμικό ηρεμίας.

20
Ποια είναι η σημασία και ο ρόλος ενός δυναμικού ενέργειας για τη μετάδοση ενός σήματος που ενεργοποιεί ένα ‘διεγέρσιμο’ κύτταρο όπως ένα νευρικό ή μυϊκό κύτταρο; Τα δυναμικά ενέργειας ακολουθούν τον Νόμο Όλα-ή-Ουδέν (All-or-None): Τα δυναμικά ενέργειας φθάνουν στο μέγιστο ή δεν πραγματοποιούνται καθόλου. Δεν υπάρχουν οι όροι μερικό ή ασθενές δυναμικό ενέργειας. Επομένως όταν το μεμβρανικό δυναμικό φθάσει στην ουδό πυροδότησης (δυναμικό ουδού ή δυναμικό πυροδότησης) τότε υφίσταται δυναμικό ενέργειας, ενώ όταν δεν φθάσει τότε δεν υπάρχει δυναμικό ενέργειας. Ένα τοπικό δυναμικό ενέργειας μπορεί να διαδοθεί σε παρακείμενες θέσεις της κυτταρικής μεμβράνης και αυτός είναι ένας τρόπος με τον οποίο διαδίδονται τα αρχικά ερεθίσματα. Αυτή η διάδοση των δυναμικών ενέργειας είναι σημαντική στις νευρικές ώσεις και τη μυϊκή συστολή.
: Τα δυναμικά ενέργειας φθάνουν στο μέγιστο ή δεν πραγματοποιούνται καθόλου. Δεν υπάρχουν οι όροι μερικό ή ασθενές δυναμικό ενέργειας. Επομένως όταν το μεμβρανικό δυναμικό φθάσει στην ουδό πυροδότησης (δυναμικό ουδού ή δυναμικό πυροδότησης) τότε υφίσταται δυναμικό ενέργειας, ενώ όταν δεν φθάσει τότε δεν υπάρχει δυναμικό ενέργειας. Ένα τοπικό δυναμικό ενέργειας μπορεί να διαδοθεί σε παρακείμενες θέσεις της κυτταρικής μεμβράνης και αυτός είναι ένας τρόπος με τον οποίο διαδίδονται τα αρχικά ερεθίσματα. Αυτή η διάδοση των δυναμικών ενέργειας είναι σημαντική στις νευρικές ώσεις και τη μυϊκή συστολή.")
21
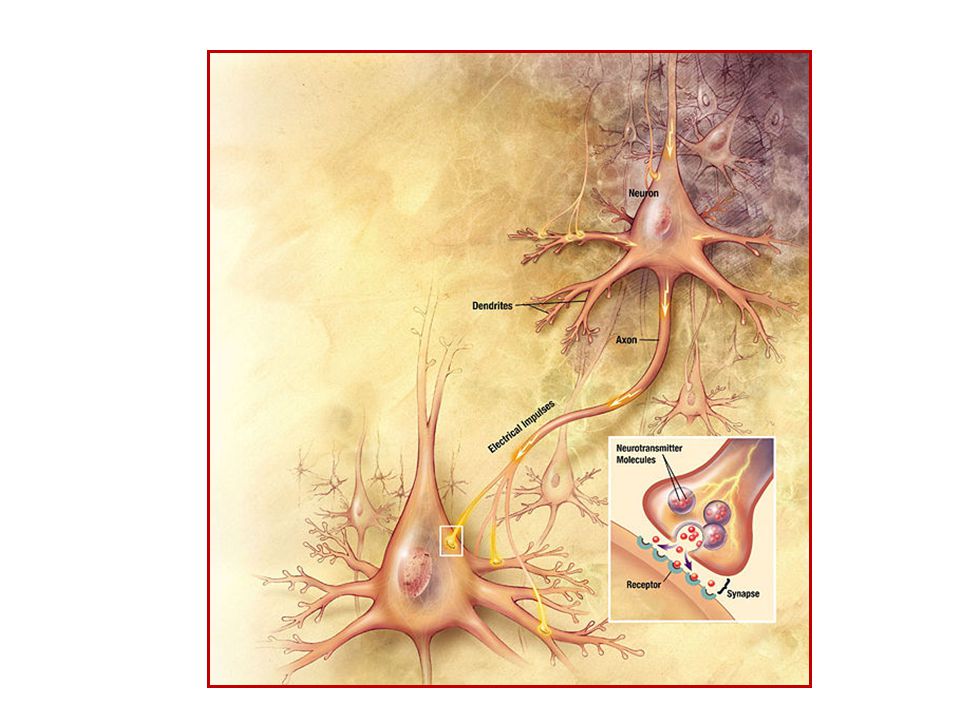
Διάδοση ενός δυναμικού ενέργειας σε ένα ‘διεγέρσιμο’ κύτταρο
Ένας νευρώνας είναι ένα νευρικό κύτταρο φέρει μικρές προεξοχές, τους δενδρίτες, και μια μακριά προεξοχή που ονομάζεται νευράξονας. Οι απολήξεις των δενδριτών ονομάζονται συνάψεις και λαμβάνουν χημικά σήματα από άλλους νευρώνες. Οι απολήξεις του νευράξονα απελευθερώνουν χημικά σήματα τα οποία λαμβάνουν οι συνάψεις γειτονικών νευρώνων. Η διάμετρος ενός νευράξονα διαφέρει από νευρώνα σε νευρώνα και από είδος σε είδος. Ο νευράξονας κινητικών (κινητήριων) νευρώνων είναι ~10 μm. Οι νευράξονες που διασυνδέονται με άλλους νευρώνες επικαλύπτονται από ένα έλυτρο (θήκη) μυελίνης (εκκρίνεται από νευρογλοιακά κύτταρα, τα κύτταρα Schwann). Η μυελίνη δρα ως μονωτής παρεμποδίζοντας τη διαρροή ιόντων και επομένως αυξάνουν την ταχύτητα διάδοσης των δυναμικών ενέργειας (και κατ’ ακολουθία τις νευρικές ώσεις). Σ’ αυτές τις εμμύελες νευρικές ίνες, το έλυτρο μυελίνης διακόπτεται ανά 1-3 mm από τους κόμβους του Ranvier (πλάτος ~1 μm), όπου λαμβάνει χώρα η διάχυση των ιόντων. Η πυκνότητα των διαύλων ιόντων είναι πολύ μεγαλύτερη σ’ αυτούς τους κόμβους: Ενώ υπάρχουν ~20 δίαυλοι ιόντων Na+/μm2 σε έναν αμύελο νευράξονα, υπάρχουν ~104 δίαυλοι Na+/μm2 σε κάθε κόμβο του Ranvier σ’ έναν εμμύελο νευράξονα. Η διάδοση μια νευρικής ώσης (διάδοση δυναμικού ενέργειας) κατά μήκος ενός νευράξονα αρχίζει όταν οι συνάψεις λαμβάνουν νευροδιαβιβαστές από παρακείμενες (γειτονικές) νευρικές απολήξεις. Η μυελίνη αποτελείται από 80% λιπίδια: σφιγγομυελίνη και 20% πρωτεΐνη: myelin basic protein (MBP), myelin oligodendrocyte glycoprotein (MOG), and proteolipid protein (PLP)].
 νευρώνων είναι ~10 μm. Οι νευράξονες που διασυνδέονται με άλλους νευρώνες επικαλύπτονται από ένα έλυτρο (θήκη) μυελίνης (εκκρίνεται από νευρογλοιακά κύτταρα, τα κύτταρα Schwann). Η μυελίνη δρα ως μονωτής παρεμποδίζοντας τη διαρροή ιόντων και επομένως αυξάνουν την ταχύτητα διάδοσης των δυναμικών ενέργειας (και κατ’ ακολουθία τις νευρικές ώσεις). Σ’ αυτές τις εμμύελες νευρικές ίνες, το έλυτρο μυελίνης διακόπτεται ανά 1-3 mm από τους κόμβους του Ranvier (πλάτος ~1 μm), όπου λαμβάνει χώρα η διάχυση των ιόντων. Η πυκνότητα των διαύλων ιόντων είναι πολύ μεγαλύτερη σ’ αυτούς τους κόμβους: Ενώ υπάρχουν ~20 δίαυλοι ιόντων Na+/μm2 σε έναν αμύελο νευράξονα, υπάρχουν ~104 δίαυλοι Na+/μm2 σε κάθε κόμβο του Ranvier σ’ έναν εμμύελο νευράξονα. Η διάδοση μια νευρικής ώσης (διάδοση δυναμικού ενέργειας) κατά μήκος ενός νευράξονα αρχίζει όταν οι συνάψεις λαμβάνουν νευροδιαβιβαστές από παρακείμενες (γειτονικές) νευρικές απολήξεις. Η μυελίνη αποτελείται από 80% λιπίδια: σφιγγομυελίνη και 20% πρωτεΐνη: myelin basic protein (MBP), myelin oligodendrocyte glycoprotein (MOG), and proteolipid protein (PLP)].")
22
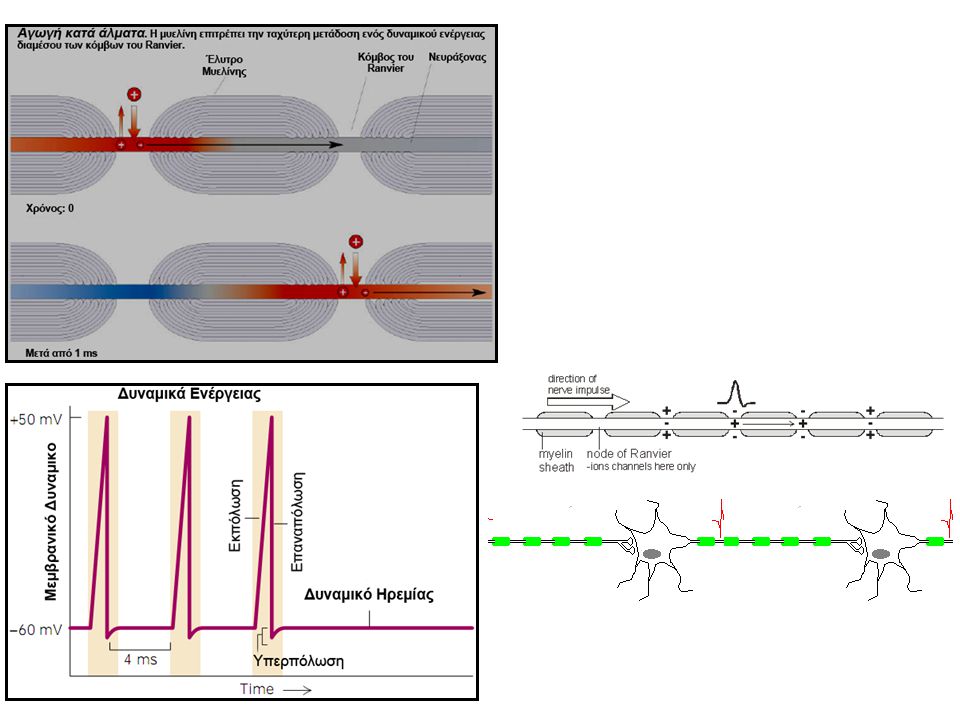
Πως διαδίδεται (άγεται) ένα δυναμικό ενέργειας σε ένα ‘διεγέρσιμο’ κύτταρο
Αμύελη Ίνα Εμμύελη Ίνα Απεικονίζεται μια αμύελη νευρική ίνα που έχει διεγερθεί στο μέσο της (αύξηση της διαπερατότητας σε Na+) (κόκκινο). Τα βέλη απεικονίζουν ένα ‘τοπικό κύκλωμα’ ροής ρευμάτων μεταξύ των εκπολωμένων περιοχών της μεμβράνης και των αμέσως γειτονικών περιοχών που βρίσκονται σε ηρεμία (μπλέ). Θετικά ηλεκτρικά φορτία (+) μεταφέρονται προς τα έσω με τα διαχεόμενα ιόντα Na+ από την εκπολωμένη μεμβράνη και στη συνέχεια μετακινούνται για μερικά mm κατά μήκος του εσωτερικού του νευράξονα. Αυτά τα (+) φορτία αυξάνουν το δυναμικό 1-3 mm μέσα στην ίνα πάνω από το δυναμικό πυροδότησης. Επομένως, οι δίαυλοι Na+ στις νέες αυτές περιοχές (μπλέ) ενεργοποιούνται και το δυναμικό ενέργειας επεκτείνεται μ’ αυτόν τον τρόπο εκατέρωθεν της διεγερθείσης περιοχής. Απεικονίζεται μια εμμύελη ίνα που έχει διεγερθεί σε έναν κόμβο του Ranvier (αύξηση της διαπερατότητας σε Na+) (κόκκινο), καθώς είναι οι περιοχές διάχυσης ιόντων. Θετικά ηλεκτρικά φορτία (+) μεταφέρονται προς τα έσω με τα διαχεόμενα ιόντα Na+ από την εκπολωμένη μεμβράνη που αυξάνουν το δυναμικό μέσα στην ίνα πάνω από το δυναμικό πυροδότησης, και στη συνέχεια μετακινούνται στον επόμενο κόμβο του Ranvier (μπλέ). Οι δίαυλοι Na+ στις νέες αυτές περιοχές (μπλέ) ενεργοποιούνται και το δυναμικό ενέργειας επεκτείνεται μ’ αυτόν τον τρόπο εκατέρωθεν της διεγερθείσης περιοχής. Αυτός ο τρόπος μετάδοσης του δυναμικού ενέργειας (και κα’ ακολουθία μιας νευρικής ώσης) διαμέσου των κόμβων του Ranvier ονομάζεται κατά άλματα αγωγή. Η ταχύτητα διάδοσης μιας νευρικής ώσης εξαρτάται από τη διάμετρο μιας νευρικής ίνας και από την παρουσία μυελίνης. Η ταχύτητα σε κινητήριες (εμμύελες) νευρικές ίνες κυμαίνεται από m/s, ενώ σε αμύελες ίνες από m/s.
 (κόκκινο). Τα βέλη απεικονίζουν ένα ‘τοπικό κύκλωμα’ ροής ρευμάτων μεταξύ των εκπολωμένων περιοχών της μεμβράνης και των αμέσως γειτονικών περιοχών που βρίσκονται σε ηρεμία (μπλέ). Θετικά ηλεκτρικά φορτία (+) μεταφέρονται προς τα έσω με τα διαχεόμενα ιόντα Na+ από την εκπολωμένη μεμβράνη και στη συνέχεια μετακινούνται για μερικά mm κατά μήκος του εσωτερικού του νευράξονα. Αυτά τα (+) φορτία αυξάνουν το δυναμικό 1-3 mm μέσα στην ίνα πάνω από το δυναμικό πυροδότησης. Επομένως, οι δίαυλοι Na+ στις νέες αυτές περιοχές (μπλέ) ενεργοποιούνται και το δυναμικό ενέργειας επεκτείνεται μ’ αυτόν τον τρόπο εκατέρωθεν της διεγερθείσης περιοχής. Απεικονίζεται μια εμμύελη ίνα που έχει διεγερθεί σε έναν κόμβο του Ranvier (αύξηση της διαπερατότητας σε Na+) (κόκκινο), καθώς είναι οι περιοχές διάχυσης ιόντων. Θετικά ηλεκτρικά φορτία (+) μεταφέρονται προς τα έσω με τα διαχεόμενα ιόντα Na+ από την εκπολωμένη μεμβράνη που αυξάνουν το δυναμικό μέσα στην ίνα πάνω από το δυναμικό πυροδότησης, και στη συνέχεια μετακινούνται στον επόμενο κόμβο του Ranvier (μπλέ). Οι δίαυλοι Na+ στις νέες αυτές περιοχές (μπλέ) ενεργοποιούνται και το δυναμικό ενέργειας επεκτείνεται μ’ αυτόν τον τρόπο εκατέρωθεν της διεγερθείσης περιοχής. Αυτός ο τρόπος μετάδοσης του δυναμικού ενέργειας (και κα’ ακολουθία μιας νευρικής ώσης) διαμέσου των κόμβων του Ranvier ονομάζεται κατά άλματα αγωγή. Η ταχύτητα διάδοσης μιας νευρικής ώσης εξαρτάται από τη διάμετρο μιας νευρικής ίνας και από την παρουσία μυελίνης. Η ταχύτητα σε κινητήριες (εμμύελες) νευρικές ίνες κυμαίνεται από m/s, ενώ σε αμύελες ίνες από m/s.")
24
ΜΟΡΙΑΚΗ ΔΟΜΗ ΔΙΑΥΛΩΝ

25
Μοριακή Αρχιτεκτονική Διαύλων
Τετραμερές: Ο δίαυλος ιόντων K+ είναι ένα τετραμερές 4 πανομοιότυπων υπομονάδων (I-IV), που κάθε μια φέρει 6 διαμεμβρανικούς α-έλικες (S1-S6). Ο S4 φέρει θετικά φορτισμένα αμινοξέα και θεωρείται ως ο τασεο-ευαίσθητος α-έλικας. Το Ν-τελικό άκρο κάθε μιας υπομονάδας έχει σφαιρική μορφή και είναι απαραίτητο για την απενεργοποίηση του διαύλου. Οι 4 υπομονάδες σχηματίζουν τον δίαυλο ιόντων K+. Ο πόρος του διαύλου σχηματίζεται από του έλικες S5 & S6 και το συνδετικό βρόγχο P. Πορεία αγωγιμότητας ιόντων Μήκος διαύλου: Å Εσωτερικός πόρος: Å Εσωτερική κοιλότητα: 10 Å με υδρόφοβα αμινοξέα Ηθμός επιλεκτικότητας (Gly-Tyr-Gly) διαχωρίζει την κεντρική κοιλότητα από τον εξωκυττάριο χώρο Σε αντίθεση, οι δίαυλοι ιόντων Na+ και Ca++ αποτελούνται από ένα πολυπεπτίδιο με 4 ομόλογες διαμεμβρανικές επαναλαμβανόμενες υποπεριοχές (I-IV) σε μια πρωτεΐνη (Μονομερή). Καθεμιά περιοχή φέρει 6 διαμεμβρανικούς α-έλικες (S1-S6), όμοιες με αυτές των διαύλων K+.
, που κάθε μια φέρει 6 διαμεμβρανικούς α-έλικες (S1-S6). Ο S4 φέρει θετικά φορτισμένα αμινοξέα και θεωρείται ως ο τασεο-ευαίσθητος α-έλικας. Το Ν-τελικό άκρο κάθε μιας υπομονάδας έχει σφαιρική μορφή και είναι απαραίτητο για την απενεργοποίηση του διαύλου. Οι 4 υπομονάδες σχηματίζουν τον δίαυλο ιόντων K+. Ο πόρος του διαύλου σχηματίζεται από του έλικες S5 & S6 και το συνδετικό βρόγχο P. Πορεία αγωγιμότητας ιόντων. Μήκος διαύλου: 45 Å. Εσωτερικός πόρος: 18 Å. Εσωτερική κοιλότητα: 10 Å. με υδρόφοβα αμινοξέα. Ηθμός επιλεκτικότητας (Gly-Tyr-Gly) διαχωρίζει την κεντρική κοιλότητα από τον εξωκυττάριο χώρο. Σε αντίθεση, οι δίαυλοι ιόντων Na+ και Ca++ αποτελούνται από ένα πολυπεπτίδιο με 4 ομόλογες διαμεμβρανικές επαναλαμβανόμενες υποπεριοχές (I-IV) σε μια πρωτεΐνη (Μονομερή). Καθεμιά περιοχή φέρει 6 διαμεμβρανικούς α-έλικες (S1-S6), όμοιες με αυτές των διαύλων K+.")
26
Δίαυλος ιόντων K+ είναι το ιδανικό παράδειγμα:
τουλάχιστον 10,000 φορές υψηλότερη από των ιόντων Na+) Η αγωγιμότητα των ιόντων είναι πολύ αποδοτική (κοντά στο όριο της ελεύθερης διάχυσης, 108 ιόντα/sec) Έχει αισθητήρα δυναμικού (voltage- sensor) Απενεργοποιείται γρήγορα
 Η αγωγιμότητα των ιόντων είναι πολύ αποδοτική (κοντά. στο όριο της ελεύθερης διάχυσης, 108 ιόντα/sec) Έχει αισθητήρα δυναμικού (voltage- sensor) Απενεργοποιείται γρήγορα.")
27
Δίαυλος ιόντων K+ Η πορεία αγωγιμότητας του ιόντος αρχίζει στη εσωτερική πλευρά όπου η πύλη του διαύλου είναι ευρεία και μπορεί να δεχθεί το ιόν στην ενυδατωμένη σφαιρική μορφή. Καθώς εισέρχεται διαμέσου της μεμβράνης, η πύλη του διαύλου στενεύει με αποτέλεσμα το νερό να απομακρύνεται από το ιόν. Η απομάκρυνση του νερού οφείλεται στον ηθμό επιλεκτικότητας του διαύλου που δεσμεύει το ιόν K+ σχηματίζοντας μια σφαίρα συντονισμού που επιτρέπει τη μετακίνηση του. Τα ιόντα Na+ δεν μπορούν να αλληλεπιδράσουν τέλεια με τον ηθμό καθώς είναι μικρότερα, και επομένως τα ιόντα K+ διαχέονται διαμέσου της πύλης του διαύλου. Καθεμιά από τις 4 πανομοιότυπες υπομονάδες φέρει 6 διαμεμβρανικούς α- έλικες, S1-S6. S4 - Τασεο-ευαίσθητος έλικας (Voltage sensor) Βρόγχος P (Segment P) – Εντοπίζεται μεταξύ S5-S6 στην πύλη του διαύλου και δρα ως ηθμός επιλεκτικότητας του πόρου. Ν-τελικό άκρο κάθε πολυπεπτιδίου έχει σφαιρική δομή και είναι απαραίτητο για την απενεργοποίηση του διαύλου Οξυγόνα από τις καρβονυλικές ομάδες της Gly στην υποπεριοχή-P δεσμεύουν το ιόν K+ Roddrick MacKinnon Nobel Χημείας 2003
 Βρόγχος P (Segment P) – Εντοπίζεται μεταξύ S5-S6 στην πύλη του διαύλου και δρα ως ηθμός επιλεκτικότητας του πόρου. Ν-τελικό άκρο κάθε πολυπεπτιδίου έχει σφαιρική δομή και είναι απαραίτητο για την απενεργοποίηση του διαύλου. Οξυγόνα από τις καρβονυλικές ομάδες της Gly στην υποπεριοχή-P δεσμεύουν το ιόν K+ Roddrick MacKinnon. Nobel Χημείας")
28
Μηχανισμός λειτουργίας της πύλης διαύλων ιόντων που εξαρτώνται από το ηλεκτρικό δυναμικό (Voltage-gated ion Channel) Μοντέλο ‘σφαίρας’ και ‘αλύσου’ (‘Ball’ and ‘Chain’ Model) Ο τασεο-εξαρτώμενος έλικας S4 ή ‘αισθητήρας του δυναμικού’ ρυθμίζει την πύλη του διαύλου ιόντων K+ καθώς φέρει το πρότυπο X-A-A, όπου X είναι ένα φορτισμένο αμινοξύ (Lys ή Arg), και A είναι ένα υδρόφοβο κατάλοιπο. Σε κατάσταση ηρεμίας το ήμισυ του έλικα S4 εντοπίζεται στο κυτταρόπλασμα. Όταν η μεμβράνη εκπολώνεται τα αμινοξέα Χ-Α-Α του S4 μετακινούνται προς τα έξω και εκτίθενται στην εξωκυτταρική πλευρά της πύλης του διαύλου με αποτέλεσμα την είσοδο ενός ενυδατωμένου ιόντος K+. Οι 4 υποπεριοχές-P, μια από κάθε υπομονάδα σχηματίζουν τον ιοντο-επιλεκτικό ηθμό στην πύλη του διαύλου ιόντων K+. Το πρότυπο Gly-Tyr-Gly προεξέχει προς τα μέσα απομακρύνει το νερό και δεσμεύει το ιόν K+ το οποίο μετακινείται διαμέσου της πύλης. Ένα χαρακτηριστικό της πύλης είναι η απενεργοποίηση της: Σύντομα μετά το άνοιγμα, η πύλη κλείνει αυθόρμητα προκαλώντας την απενεργοποίηση του διαύλου. Η πύλη δεν ανοίγει μέχρι την επαναπόλωση της μεμβράνης. Τα Ν-τελικά άκρα καθενός πολυπεπτιδίου σχηματίζουν μια σφαιρική δομή (το αποφρακτικό σωματίδιο) και ένα απ’ αυτά ταλαντεύεται προς την ανοικτή πύλη του διαύλου και την απενεργοποιεί. Να σημειωθεί ότι στην κατάσταση ηρεμίας ο δίαυλος ιόντων K+ δεν λειτουργεί ως τασεο-εξαρτώμενος δίαυλος.
 Ο τασεο-εξαρτώμενος έλικας S4 ή ‘αισθητήρας του δυναμικού’ ρυθμίζει την πύλη του διαύλου ιόντων K+ καθώς φέρει το πρότυπο X-A-A, όπου X είναι ένα φορτισμένο αμινοξύ (Lys ή Arg), και A είναι ένα υδρόφοβο κατάλοιπο. Σε κατάσταση ηρεμίας το ήμισυ του έλικα S4 εντοπίζεται στο κυτταρόπλασμα. Όταν η μεμβράνη εκπολώνεται τα αμινοξέα Χ-Α-Α του S4 μετακινούνται προς τα έξω και εκτίθενται στην εξωκυτταρική πλευρά της πύλης του διαύλου με αποτέλεσμα την είσοδο ενός ενυδατωμένου ιόντος K+. Οι 4 υποπεριοχές-P, μια από κάθε υπομονάδα σχηματίζουν τον ιοντο-επιλεκτικό ηθμό στην πύλη του διαύλου ιόντων K+. Το πρότυπο Gly-Tyr-Gly προεξέχει προς τα μέσα απομακρύνει το νερό και δεσμεύει το ιόν K+ το οποίο μετακινείται διαμέσου της πύλης. Ένα χαρακτηριστικό της πύλης είναι η απενεργοποίηση της: Σύντομα μετά το άνοιγμα, η πύλη κλείνει αυθόρμητα προκαλώντας την απενεργοποίηση του διαύλου. Η πύλη δεν ανοίγει μέχρι την επαναπόλωση της μεμβράνης. Τα Ν-τελικά άκρα καθενός πολυπεπτιδίου σχηματίζουν μια σφαιρική δομή (το αποφρακτικό σωματίδιο) και ένα απ’ αυτά ταλαντεύεται προς την ανοικτή πύλη του διαύλου και την απενεργοποιεί. Να σημειωθεί ότι στην κατάσταση ηρεμίας ο δίαυλος ιόντων K+ δεν λειτουργεί ως τασεο-εξαρτώμενος δίαυλος.")
29
Δίαυλος ιόντων Na+ Η δομή και η λειτουργία των διαύλων Na+ είναι όμοια μ’ αυτή των διαύλων ιόντων K+, αλλά πιο πολύπλοκη. Δομή Διαύλου ιόντων Na+: Ο δίαυλος αποτελείται από 3 πρωτεϊνικές υπομονάδες: μια μεγάλη γλυκοπρωτεΐνη α και 2 μικρότερα πολυπεπτίδια β1 και β2. Η α-υπομονάδα σχηματίζει την πύλη του διαύλου, ενώ οι μικρότερες β-υπομονάδες αλληλεπιδρούν και ρυθμίζουν την α-υπομονάδα. Όπως και στο δίαυλο ιόντων K+, το φορτίο της υποπεριοχής S4 επηρεάζει το άνοιγμα και το κλείσιμο του τασεο-εξαρτώμενου διαύλου ιόντων Na+. Σε κατάσταση ηρεμίας, το καθαρό αρνητικό φορτίο μέσα από τη μεμβράνη ελκύει τη θετικά φορτισμένη υποπεριοχή S4 μέσα στη μεμβράνη και επομένως η πύλη είναι κλειστή. Όμως κατά την εκπόλωση της μεμβράνης, η μεταβολή στο ηλεκτρικό φορτίο κατά πλάτος της μεμβράνης ωθεί την υποπεριοχή S4 να κινηθεί προς την εξωκυττάρια πλευρά της μεμβράνης με αποτέλεσμα το άνοιγμα της πύλης του διαύλου. (Α) α-έλικες S5 & S6 επενδύουν την πύλη του διαύλου, ενώ ο S4 είναι ο τασεο-ευαίσθητος α-έλικας. Οι υποπεριοχές III & IV εμπλέκονται στην απενεργοποίηση του διαύλου. Απεικονίζεται επίσης η περιοχή ρύθμισης με φωσφορυλίωση. (Β) 3-D Δομή της α-υπομονάδας
 α-έλικες S5 & S6 επενδύουν την πύλη του διαύλου, ενώ ο S4 είναι ο τασεο-ευαίσθητος α-έλικας. Οι υποπεριοχές III & IV εμπλέκονται στην απενεργοποίηση του διαύλου. Απεικονίζεται επίσης η περιοχή ρύθμισης με φωσφορυλίωση. (Β) 3-D Δομή της α-υπομονάδας.")
30
Λειτουργία Πύλης Διαύλου ιόντων Na+: Όπως και ο δίαυλος ιόντων K+, και ο δίαυλος ιόντων Na+ αποκτά 3 διαμορφώσεις: κλειστή ή απενεργοποιημένη (closed ή deactivated), ανοικτή (open) και ανενεργό (inactivated). Η ανενεργός διαμόρφωση (inactivated state) οφείλεται στο αποφρακτικό σωματίδιο που σχηματίζεται από τις υποπεριοχές III και IV της α-υπομονάδας, που ονομάζεται ‘πύλη απενεργοποίησης’ (inactivation gate), η οποία παρεμποδίζει το εσωτερικό του διαύλου λίγο μετά το άνοιγμα του διαύλου. Κατά τη διάρκεια ενός δυναμικού ενέργειας ο δίαυλος παραμένει ανενεργός (inactive) για λίγα ms μετά την εκπόλωση της μεμβράνης. Η ανενεργός διαμόρφωση μεταβάλλεται προς την κλειστή διαμόρφωση όταν η μεμβράνη επαναπολώνεται κατά τη φθίνουσα φάση του δυναμικού ενέργειας. Αυτή η μεταβολή επιτρέπει την επανεργοποίηση του διαύλου κατά το δεύτερο δυναμικό ενέργειας. Η λειτουργία του διαύλου των ιόντων Na+ είναι πολύπλοκη και μπορεί να περιγραφεί με το Μοντέλο Markov ή το Μοντέλο Hodgkin-Huxley. Γενετικές ασθένειες που επηρεάζουν τη λειτουργία του διαύλου προκαλούν μεταξύ άλλων μυοτονία και επιληψία.
 οφείλεται στο αποφρακτικό σωματίδιο που σχηματίζεται από τις υποπεριοχές III και IV της α-υπομονάδας, που ονομάζεται ‘πύλη απενεργοποίησης’ (inactivation gate), η οποία παρεμποδίζει το εσωτερικό του διαύλου λίγο μετά το άνοιγμα του διαύλου. Κατά τη διάρκεια ενός δυναμικού ενέργειας ο δίαυλος παραμένει ανενεργός (inactive) για λίγα ms μετά την εκπόλωση της μεμβράνης. Η ανενεργός διαμόρφωση μεταβάλλεται προς την κλειστή διαμόρφωση όταν η μεμβράνη επαναπολώνεται κατά τη φθίνουσα φάση του δυναμικού ενέργειας. Αυτή η μεταβολή επιτρέπει την επανεργοποίηση του διαύλου κατά το δεύτερο δυναμικό ενέργειας. Η λειτουργία του διαύλου των ιόντων Na+ είναι πολύπλοκη και μπορεί να περιγραφεί με το Μοντέλο Markov ή το Μοντέλο Hodgkin-Huxley. Γενετικές ασθένειες που επηρεάζουν τη λειτουργία του διαύλου προκαλούν μεταξύ άλλων μυοτονία και επιληψία.")
31
Οι τασεο-εξαρτώμενοι δίαυλοι ιόντων Na+ [Voltage-gated sodium channels (VGSCs)] μεταφέρουν ηλεκτρικό δυναμικό προς τα μέσα και είναι υπεύθυνοι για τη δημιουργία δυναμικών ενέργειας στις νευρικές ίνες, σκελετικό και καρδιακό μυ. Οι VGSCs είναι συνήθως κλειστοί σε φυσιολογικά δυναμικά ηρεμίας Όταν η μεμβράνη εκπολωθεί, οι VGSCs ανοίγουν (ενεργοποιούνται) και μετά απενεργοποιούνται και αποκτούν μια κλειστή διαμόρφωση η οποία δεν μπορεί να ανοίξει πάλι ακόμη και αν η μεμβράνη εκπολωθεί σε μεγαλύτερο βαθμό. Για να καταστεί δυνατή η ενεργοποίηση της πύλης, οι δίαυλοι θα πρέπει να ανακάμψουν από την απενεργοποιημένη διαμόρφωση. Η ανάκτηση της ικανότητας του διαύλου επιτυγχάνεται κατά την επαναπόλωση της μεμβράνης στο δυναμικό ηρεμίας. Στα περισσότερα κύτταρα, η ανάκτηση από την απενεργοποιημένη διαμόρφωση είναι ηλεκτρικά σιωπηλή (δεν υπάρχει ροή ιόντων Na+) (Σχήμα). Κλασσικό μοντέλο διαύλων ιόντων Na+ σε νευρικές ίνες. Οι δίαυλοι είναι κλειστοί στα –90 mV. Όταν η μεμβράνη εκπολώνεται στα +30 mV, οι δίαυλοι ανοίγουν και μεταφέρουν ιόντα προς τα μέσα. Μετά από ελάχιστα ms, οι δίαυλοι απενεργοποιούνται λόγω της παρεμπόδισης της πύλης του διαύλου (α-υπομονάδα) από το αποφρακτικό σωματίδιο. Καθώς η μεμβράνη επαναπολώνεται στα –50 mV, οι δίαυλοι επανέρχονται μερικώς από την απενεργοποιημένη μορφή, πίσω στην κλειστή διαμόρφωση τους που δεν επιτρέπει τη ροή ιόντων.
![Οι τασεο-εξαρτώμενοι δίαυλοι ιόντων Na+ [Voltage-gated sodium channels (VGSCs)] μεταφέρουν ηλεκτρικό δυναμικό προς τα μέσα και είναι υπεύθυνοι για τη δημιουργία δυναμικών ενέργειας στις νευρικές ίνες, σκελετικό και καρδιακό μυ. Οι VGSCs είναι συνήθως κλειστοί σε φυσιολογικά δυναμικά ηρεμίας Όταν η μεμβράνη εκπολωθεί, οι VGSCs ανοίγουν (ενεργοποιούνται) και μετά απενεργοποιούνται και αποκτούν μια κλειστή διαμόρφωση η οποία δεν μπορεί να ανοίξει πάλι ακόμη και αν η μεμβράνη εκπολωθεί σε μεγαλύτερο βαθμό. Για να καταστεί δυνατή η ενεργοποίηση της πύλης, οι δίαυλοι θα πρέπει να ανακάμψουν από την απενεργοποιημένη διαμόρφωση. Η ανάκτηση της ικανότητας του διαύλου επιτυγχάνεται κατά την επαναπόλωση της μεμβράνης στο δυναμικό ηρεμίας. Στα περισσότερα κύτταρα, η ανάκτηση από την απενεργοποιημένη διαμόρφωση είναι ηλεκτρικά σιωπηλή (δεν υπάρχει ροή ιόντων Na+) (Σχήμα).](http://slideplayer.gr/slide/5247127/16/images/31/%CE%9F%CE%B9+%CF%84%CE%B1%CF%83%CE%B5%CE%BF-%CE%B5%CE%BE%CE%B1%CF%81%CF%84%CF%8E%CE%BC%CE%B5%CE%BD%CE%BF%CE%B9+%CE%B4%CE%AF%CE%B1%CF%85%CE%BB%CE%BF%CE%B9+%CE%B9%CF%8C%CE%BD%CF%84%CF%89%CE%BD+Na%2B+%5BVoltage-gated+sodium+channels+%28VGSCs%29%5D+%CE%BC%CE%B5%CF%84%CE%B1%CF%86%CE%AD%CF%81%CE%BF%CF%85%CE%BD+%CE%B7%CE%BB%CE%B5%CE%BA%CF%84%CF%81%CE%B9%CE%BA%CF%8C+%CE%B4%CF%85%CE%BD%CE%B1%CE%BC%CE%B9%CE%BA%CF%8C+%CF%80%CF%81%CE%BF%CF%82+%CF%84%CE%B1+%CE%BC%CE%AD%CF%83%CE%B1+%CE%BA%CE%B1%CE%B9+%CE%B5%CE%AF%CE%BD%CE%B1%CE%B9+%CF%85%CF%80%CE%B5%CF%8D%CE%B8%CF%85%CE%BD%CE%BF%CE%B9+%CE%B3%CE%B9%CE%B1+%CF%84%CE%B7+%CE%B4%CE%B7%CE%BC%CE%B9%CE%BF%CF%85%CF%81%CE%B3%CE%AF%CE%B1+%CE%B4%CF%85%CE%BD%CE%B1%CE%BC%CE%B9%CE%BA%CF%8E%CE%BD+%CE%B5%CE%BD%CE%AD%CF%81%CE%B3%CE%B5%CE%B9%CE%B1%CF%82+%CF%83%CF%84%CE%B9%CF%82+%CE%BD%CE%B5%CF%85%CF%81%CE%B9%CE%BA%CE%AD%CF%82+%CE%AF%CE%BD%CE%B5%CF%82%2C+%CF%83%CE%BA%CE%B5%CE%BB%CE%B5%CF%84%CE%B9%CE%BA%CF%8C+%CE%BA%CE%B1%CE%B9+%CE%BA%CE%B1%CF%81%CE%B4%CE%B9%CE%B1%CE%BA%CF%8C+%CE%BC%CF%85.+%CE%9F%CE%B9+VGSCs+%CE%B5%CE%AF%CE%BD%CE%B1%CE%B9+%CF%83%CF%85%CE%BD%CE%AE%CE%B8%CF%89%CF%82+%CE%BA%CE%BB%CE%B5%CE%B9%CF%83%CF%84%CE%BF%CE%AF+%CF%83%CE%B5+%CF%86%CF%85%CF%83%CE%B9%CE%BF%CE%BB%CE%BF%CE%B3%CE%B9%CE%BA%CE%AC+%CE%B4%CF%85%CE%BD%CE%B1%CE%BC%CE%B9%CE%BA%CE%AC+%CE%B7%CF%81%CE%B5%CE%BC%CE%AF%CE%B1%CF%82+%CE%8C%CF%84%CE%B1%CE%BD+%CE%B7+%CE%BC%CE%B5%CE%BC%CE%B2%CF%81%CE%AC%CE%BD%CE%B7+%CE%B5%CE%BA%CF%80%CE%BF%CE%BB%CF%89%CE%B8%CE%B5%CE%AF%2C+%CE%BF%CE%B9+VGSCs+%CE%B1%CE%BD%CE%BF%CE%AF%CE%B3%CE%BF%CF%85%CE%BD+%28%CE%B5%CE%BD%CE%B5%CF%81%CE%B3%CE%BF%CF%80%CE%BF%CE%B9%CE%BF%CF%8D%CE%BD%CF%84%CE%B1%CE%B9%29+%CE%BA.jpg "Κλασσικό μοντέλο διαύλων ιόντων Na+ σε νευρικές ίνες. Οι δίαυλοι είναι κλειστοί στα –90 mV. Όταν η μεμβράνη εκπολώνεται στα +30 mV, οι δίαυλοι ανοίγουν και μεταφέρουν ιόντα προς τα μέσα. Μετά από ελάχιστα ms, οι δίαυλοι απενεργοποιούνται λόγω της παρεμπόδισης της πύλης του διαύλου (α-υπομονάδα) από το αποφρακτικό σωματίδιο. Καθώς η μεμβράνη επαναπολώνεται στα –50 mV, οι δίαυλοι επανέρχονται μερικώς από την απενεργοποιημένη μορφή, πίσω στην κλειστή διαμόρφωση τους που δεν επιτρέπει τη ροή ιόντων.")
32
Οι πρωτεΐνες των τασεο-εξαρτώμενων διαύλων πιθανά εξελίχθηκαν από κοινά προγονικά γονίδια
Οι δίαυλοι είναι μακρομοριακά πρωτεϊνικά συμπλέγματα ενσωματωμένα στη διπλοστιβάδα λιπιδίων των κυτταρικών μεμβρανών. Αποτελούνται από υπομονάδες καθεμιά από τις οποίες να έχει συγκεκριμένη λειτουργία και κωδικοποιείται από ένα διαφορετικό γονίδιο. Οι δομικές και λειτουργικές ομοιότητες μεταξύ των τασεο-εξαρτώμενων διαύλων ιόντων Na+, Ca2+, and K+ καταδεικνύουν ότι και οι τρεις πρωτεΐνες προήλθαν από ένα κοινό προγονικό γονίδιο (common ancestral gene). Η κατανομή αυτών των διαύλων στους διαφορετικούς οργανισμούς (θηλαστικά, φυτά, βακτήρια, κλπ) παρέχει ενδείξεις ως προς την πορεία εξέλιξης τους. Οι τασεο-εξαρτώμενοι δίαυλοι K+ απαντώνται επίσης στους σακχαρομύκητες και πρωτόζωα, ενώ οι τασεο-εξαρτώμενοι δίαυλοι Na+ απαντώνται μόνο στους πολυκύτταρους οργανισμούς. Ενδιάμεσοι στην κατανομή είναι οι τασεο-εξαρτώμενοι δίαυλοι ιόντων Ca2+ οι οποίοι δρουν κατά τη συναπτική διαβίβαση σημάτων: Απαντώνται στους πολυκύτταρους οργανισμούς και μόνο σε πολύπλοκα πρωτόζωα όπως το Paramecium. Επομένως οι πρωτεΐνες των τασεο-εξαρτώμενων διαύλων ιόντων K+ προήλθαν πρώτες κατά την εξέλιξη. Οι πρωτεΐνες των τασεο-εξαρτώμενων διαύλων ιόντων Ca2+ και Na+ φαίνεται ότι προήλθαν με επαναλαμβανόμενους γονιδιακούς διπλασιασμούς (gene duplication) ενός προγονικού γονιδίου μιας υπομονάδας του διαύλου των ιόντων K+. Αφού όλοι οι δίαυλοι ιόντων K+ φέρουν μια ίδια υποπεριοχή-P που επενδύει την πύλη του διαύλου, είναι πιθανόν ότι τασεο-εξαρτώμενοι και ανεξάρτητοι από το ηλεκτρικό δυναμικό δίαυλοι ιόντων K+ να έχουν προέλθει από ένα κοινό προγονικό γονίδιο.
. Η κατανομή αυτών των διαύλων στους διαφορετικούς οργανισμούς (θηλαστικά, φυτά, βακτήρια, κλπ) παρέχει ενδείξεις ως προς την πορεία εξέλιξης τους. Οι τασεο-εξαρτώμενοι δίαυλοι K+ απαντώνται επίσης στους σακχαρομύκητες και πρωτόζωα, ενώ οι τασεο-εξαρτώμενοι δίαυλοι Na+ απαντώνται μόνο στους πολυκύτταρους οργανισμούς. Ενδιάμεσοι στην κατανομή είναι οι τασεο-εξαρτώμενοι δίαυλοι ιόντων Ca2+ οι οποίοι δρουν κατά τη συναπτική διαβίβαση σημάτων: Απαντώνται στους πολυκύτταρους οργανισμούς και μόνο σε πολύπλοκα πρωτόζωα όπως το Paramecium. Επομένως οι πρωτεΐνες των τασεο-εξαρτώμενων διαύλων ιόντων K+ προήλθαν πρώτες κατά την εξέλιξη. Οι πρωτεΐνες των τασεο-εξαρτώμενων διαύλων ιόντων Ca2+ και Na+ φαίνεται ότι προήλθαν με επαναλαμβανόμενους γονιδιακούς διπλασιασμούς (gene duplication) ενός προγονικού γονιδίου μιας υπομονάδας του διαύλου των ιόντων K+. Αφού όλοι οι δίαυλοι ιόντων K+ φέρουν μια ίδια υποπεριοχή-P που επενδύει την πύλη του διαύλου, είναι πιθανόν ότι τασεο-εξαρτώμενοι και ανεξάρτητοι από το ηλεκτρικό δυναμικό δίαυλοι ιόντων K+ να έχουν προέλθει από ένα κοινό προγονικό γονίδιο.")
33
II. Δίαυλοι που εξαρτώνται από προσδέτη (Ligand-gated ion channels)
Υπεύθυνοι (διαμεσολαβητές) για τη γρήγορη δράση των νευροδιαβιβαστών σε συνάψεις μεταβάλλοντας το δυναμικό της μεμβράνης ως απόκριση στο νευροδιαβιβαστή [δέσμευση προσδέτη (ligand binding)]. Ενεργοποιούνται επιλεκτικά από έναν συγκεκριμένο προσδέτη. Διακρίνουν μεταξύ αρνητικά και θετικά φορτισμένων ιόντων, αλλά κατά τα άλλα δεν είναι ισχυρά επιλεκτικοί. ● Δίαυλοι που άγουν κατιόντα (Cation-conducting channels) – Υποδοχείς ακετυλοχολίνης (AcCh), σεροτονίνης και γλουταμινικού οξέος (Glu). ● Δίαυλοι που άγουν ανιόντα (Anion-conducting channels) – Υποδοχείς γλυκίνης (Gly) και γ-αμινοβουτυρικού οξέος (GABA; g-aminobutiric acid)
 για τη γρήγορη δράση των νευροδιαβιβαστών σε συνάψεις μεταβάλλοντας το δυναμικό της μεμβράνης ως απόκριση στο νευροδιαβιβαστή [δέσμευση προσδέτη (ligand binding)]. Ενεργοποιούνται επιλεκτικά από έναν συγκεκριμένο προσδέτη. Διακρίνουν μεταξύ αρνητικά και θετικά φορτισμένων ιόντων, αλλά κατά τα άλλα δεν είναι ισχυρά επιλεκτικοί. ● Δίαυλοι που άγουν κατιόντα (Cation-conducting channels) – Υποδοχείς ακετυλοχολίνης (AcCh), σεροτονίνης και γλουταμινικού οξέος (Glu). ● Δίαυλοι που άγουν ανιόντα (Anion-conducting channels) – Υποδοχείς γλυκίνης (Gly) και γ-αμινοβουτυρικού οξέος (GABA; g-aminobutiric acid)")
35
Δίαυλος που άγει κατιόντα (Cation-conducting channel)
Υποδοχέας ακετυλοχολίνης Όταν μια νευρική ώση (δυναμικό ενέργειας) φθάσει στο τελικό κομβίο (νευρική απόληξη) μιας προ-συναπτικής νευρικής ίνας, τότε η μεμβράνη γίνεται πιο διαπερατή στα ιόντα Ca2+. Τα ιόντα Ca2+ διαχέονται μέσα στη νευρική απόληξη και ενεργοποιούν ένζυμα τα οποία προκαλούν τη μετακίνηση των συναπτικών κυστιδίων προς τη συναπτική σχισμή. Μερικά κυστίδια συντήσσονται με τη μεμβράνη των νευρικών απολήξεων της προ-συναπτικής νευρικής ίνας και απελευθερώνουν το νευροδιαβιβαστή, όπως ακετυλοχολίνη (AcCh) (εξωκύττωση) στη συναπτική σχισμή. Ο νευροδιαβιβαστής διαχέεται κατά πλάτος της συναπτικής σχισμής και προσδένει στον υποδοχέα του (υποδοχέας της AcCh; AcChR) είτε στη μεμβράνη των νευρικών απολήξεων μιας μετα-συναπτικής ίνας είτε στο μυ. Όταν όλοι οι υποδοχείς καταληφθούν από το νευροδιαβιβαστή, τότε ενεργοποιούνται (ανοίγουν) οι δίαυλοι ιόντων Na+ και επιτρέπουν τη διάχυση των ιόντων Na+ προς τα μέσα. Μ’ αυτόν τον τρόπο το μεμβρανικό δυναμικό γίνεται λιγότερο αρνητικό και προσεγγίζει το δυναμικό (ουδός) πυροδότησης. Η ουδός πυροδότησης προσεγγίζεται στη περίπτωση που εκκριθεί επαρκής ποσότητα του νευροδιαβιβαστή και ενεργοποιηθούν αρκετοί δίαυλοι ιόντων Na+. Καθώς η μεμβράνη εκπολώνεται οδηγεί στην έναρξη και κορύφωση ενός δυναμικού ενέργειας. Με άλλα λόγια το σήμα μεταδίδεται είτε στη μετα-συναπτική ίνα ή στο μυ στην περίπτωση μιας νευρο-μυϊκής σύναψης. Στη συνέχεια, τα μόρια του νευροδιαβιβαστή μεταφέρονται με συγκεκριμένους μεταφορείς πίσω στις απολήξεις της προ-συναπτικής νευρικής ίνας κοντά στους υποδοχείς του όπου αποδομούνται από ένζυμα, Για παράδειγμα η Ach αποδομείται από τη χολινεστεράση..
 φθάσει στο τελικό κομβίο (νευρική απόληξη) μιας προ-συναπτικής νευρικής ίνας, τότε η μεμβράνη γίνεται πιο διαπερατή στα ιόντα Ca2+. Τα ιόντα Ca2+ διαχέονται μέσα στη νευρική απόληξη και ενεργοποιούν ένζυμα τα οποία προκαλούν τη μετακίνηση των συναπτικών κυστιδίων προς τη συναπτική σχισμή. Μερικά κυστίδια συντήσσονται με τη μεμβράνη των νευρικών απολήξεων της προ-συναπτικής νευρικής ίνας και απελευθερώνουν το νευροδιαβιβαστή, όπως ακετυλοχολίνη (AcCh) (εξωκύττωση) στη συναπτική σχισμή. Ο νευροδιαβιβαστής διαχέεται κατά πλάτος της συναπτικής σχισμής και προσδένει στον υποδοχέα του (υποδοχέας της AcCh; AcChR) είτε στη μεμβράνη των νευρικών απολήξεων μιας μετα-συναπτικής ίνας είτε στο μυ. Όταν όλοι οι υποδοχείς καταληφθούν από το νευροδιαβιβαστή, τότε ενεργοποιούνται (ανοίγουν) οι δίαυλοι ιόντων Na+ και επιτρέπουν τη διάχυση των ιόντων Na+ προς τα μέσα. Μ’ αυτόν τον τρόπο το μεμβρανικό δυναμικό γίνεται λιγότερο αρνητικό και προσεγγίζει το δυναμικό (ουδός) πυροδότησης. Η ουδός πυροδότησης προσεγγίζεται στη περίπτωση που εκκριθεί επαρκής ποσότητα του νευροδιαβιβαστή και ενεργοποιηθούν αρκετοί δίαυλοι ιόντων Na+. Καθώς η μεμβράνη εκπολώνεται οδηγεί στην έναρξη και κορύφωση ενός δυναμικού ενέργειας. Με άλλα λόγια το σήμα μεταδίδεται είτε στη μετα-συναπτική ίνα ή στο μυ στην περίπτωση μιας νευρο-μυϊκής σύναψης. Στη συνέχεια, τα μόρια του νευροδιαβιβαστή μεταφέρονται με συγκεκριμένους μεταφορείς πίσω στις απολήξεις της προ-συναπτικής νευρικής ίνας κοντά στους υποδοχείς του όπου αποδομούνται από ένζυμα, Για παράδειγμα η Ach αποδομείται από τη χολινεστεράση..")
36
Υποδοχέας της Ακετυλοχολίνης
● 5 υπομονάδες (α2βγδ) ● Κάθε υπομονάδα έχει 4 ομόλογα πολυπεπτίδια (διαμεμβρανικούς Μ1-4 α- έλικες) ● -υπομονάδες περιέχουν στην εξωκυτταρική τους πλευρά τις θέσεις δέσμευσης της AcCh ● Η δέσμευση της AcCh ανοίγει την πύλη και ενεργοποιεί το δίαυλο
 ● Κάθε υπομονάδα έχει 4 ομόλογα. πολυπεπτίδια (διαμεμβρανικούς Μ1-4 α- έλικες) ● -υπομονάδες περιέχουν στην εξωκυτταρική. τους πλευρά τις θέσεις δέσμευσης της AcCh. ● Η δέσμευση της AcCh ανοίγει την πύλη και. ενεργοποιεί το δίαυλο.")
37
AcCh – Διεγερτικός νευροδιαβιβαστής αφού καθιστά τη μεμβράνη λιγότερο αρνητική καθώς αυξάνει τη διαπερατότητα της σε ιόντα Na+. Υπάρχουν 2 τύποι υποδοχέων AcCh: Μουσκαρινικοί και νικοτινικοί υποδοχείς. Μουσκαρινικοί: Βρίσκονται σε κύτταρα που διεγείρονται από μεταγαγγλιακές νευράδες του ΠΝΣ, και μεταγαγγλιακεές χολινεργικές νευράδες το ΣΝΣ. Νικοτινικοί: Βρίσκονται σε συνάψεις προ- και μετα-γαγγλιακών νευράδων του ΠΝΣ & ΣΝΣ, και μεμβράνες μυϊκών ινών σκελετικού μυ στο σημείο της νευρομυϊκής σύναψης. Διατομή διαμέσου του κέντρου των Μ2 ελικών, ενός από καθεμιά υπομονάδα (εγκόλπωση) που απεικονίζει την παρεμπόδιση της πύλης του διαύλου από τις ογκώδεις πλευρικές αλυσίδες των καταλοίπων λευκίνης (Leu). Όταν και οι δύο θέσεις δέσμευσης της AcCh (μια σε κάθε α-υπομονάδα) είναι κατειλημμένες, τότε επέρχεται μεταβολή στη διαμόρφωση του υποδοχέα και οι Μ2 έλικες περιστρέφονται ελαφρώς με αποτέλεσμα να απομακρύνονται οι πλευρικές αλυσίδες της Leu από την πύλη του διαύλου και να αντικαθιστώνται με μικρότερα πολικά κατάλοιπα. Αυτός ο μηχανισμός ενεργοποίησης της πύλης του διαύλου επιτρέπει τη διάχυση ιόντων Na+, K+, ή Ca2+. Ο μοριακός μηχανισμός κλεισίματος της πύλης του διαύλου του υποδοχέα της AcCh δεν είναι επαρκώς κατανοητός.
 που απεικονίζει την παρεμπόδιση της πύλης του διαύλου από τις ογκώδεις πλευρικές αλυσίδες των καταλοίπων λευκίνης (Leu). Όταν και οι δύο θέσεις δέσμευσης της AcCh (μια σε κάθε α-υπομονάδα) είναι κατειλημμένες, τότε επέρχεται μεταβολή στη διαμόρφωση του υποδοχέα και οι Μ2 έλικες περιστρέφονται ελαφρώς με αποτέλεσμα να απομακρύνονται οι πλευρικές αλυσίδες της Leu από την πύλη του διαύλου και να αντικαθιστώνται με μικρότερα πολικά κατάλοιπα. Αυτός ο μηχανισμός ενεργοποίησης της πύλης του διαύλου επιτρέπει τη διάχυση ιόντων Na+, K+, ή Ca2+. Ο μοριακός μηχανισμός κλεισίματος της πύλης του διαύλου του υποδοχέα της AcCh δεν είναι επαρκώς κατανοητός.")
38
Δομική οργάνωση του Υποδοχέα του GABA
Δίαυλος που άγει ανιόντα (Anion-conducting channel) Υποδοχείς GABA Δομική οργάνωση του Υποδοχέα του GABA Ολιγομερή 5 διαφορετικών υπομονάδων οι οποίες είναι ομόλογες κατά 30-50% Ο πόρος σχηματίζεται κυρίως από την διαμεμβρανική υποπεριοχή M2 καθενός μονομερούς GABA – Ανασταλτικός νευροδιαβιβαστής αφού καθιστά τη μεμβράνη πιο αρνητική καθώς αυξάνει τη διαπερατότητα της σε ιόντα K+ GABA έχει χρησιμοποιηθεί για τη θεραπεία επιληψίας & υπέρτασης
 Υποδοχείς GABA. Δομική οργάνωση του Υποδοχέα του GABA. Ολιγομερή 5 διαφορετικών υπομονάδων οι οποίες είναι ομόλογες κατά 30-50% Ο πόρος σχηματίζεται κυρίως από την διαμεμβρανική υποπεριοχή M2 καθενός μονομερούς. GABA – Ανασταλτικός νευροδιαβιβαστής αφού καθιστά τη μεμβράνη πιο αρνητική καθώς αυξάνει τη διαπερατότητα της σε ιόντα K+ GABA έχει χρησιμοποιηθεί για τη θεραπεία επιληψίας & υπέρτασης.")
39
Μοριακή βάση της ATP-εξαρτώμενης Μεταφοράς

40
ATPάσες τύπου-P (P-type ATPases)
Μεταφέρουν διάφορα κατιόντα ενάντια μιας διαβάθμισης του ηλεκτροχημικού δυναμικού (Electrochemical potential gradient) Χρησιμοποιούν την ενέργεια που προέρχεται από την υδρόλυση του ATP και έχουν όμοιο καταλυτικό κύκλο. Σχηματίζουν σταθερά ακυλοφωσφορικά ενδιάμεσα μεταφέροντας τη γ-φωσφορική ομάδα του ATP σε συγκεκριμένα κατάλοιπα Asp στη θέση δέσμευσης του ATP. Σε αντίθεση η ATPάσες τύπου V και F δεν σχηματίζουν ακυλοφωσφορικά ενδιάμεσα και μεταφέρουν μόνον πρωτόνια. Χαρακτηρίζονται από 3 διατηρημένα πρότυπα (μοτίβα): DKTG, TGES/A και GDGxxG Απαντώνται στη πλασματική μεμβράνη των ανωτέρων ευκαρυωτικών (Αντλία Na+/K+), & όλων των ευκαρυωτικών κυττάρων (Αντλία Ca2+), στην apical πλασματική μεμβράνη των κυττάρων του στομάχου των θηλαστικών (Αντλία H+/K+) και στη μεμβράνη στου σαρκοπλασματικού δικτύου των μυϊκών κυττάρων (Αντλία Ca2+).
 Χρησιμοποιούν την ενέργεια που προέρχεται από την υδρόλυση του ATP και έχουν όμοιο καταλυτικό κύκλο. Σχηματίζουν σταθερά ακυλοφωσφορικά ενδιάμεσα μεταφέροντας τη γ-φωσφορική ομάδα του ATP σε συγκεκριμένα κατάλοιπα Asp στη θέση δέσμευσης του ATP. Σε αντίθεση η ATPάσες τύπου V και F δεν σχηματίζουν ακυλοφωσφορικά ενδιάμεσα και μεταφέρουν μόνον πρωτόνια. Χαρακτηρίζονται από 3 διατηρημένα πρότυπα (μοτίβα): DKTG, TGES/A και GDGxxG. Απαντώνται στη πλασματική μεμβράνη των ανωτέρων ευκαρυωτικών (Αντλία Na+/K+), & όλων των ευκαρυωτικών κυττάρων (Αντλία Ca2+), στην apical πλασματική μεμβράνη των κυττάρων του στομάχου των θηλαστικών (Αντλία H+/K+) και στη μεμβράνη στου σαρκοπλασματικού δικτύου των μυϊκών κυττάρων (Αντλία Ca2+).")
41
ATPάση Na+/ K (Na+/K-ATPase)
Ανήκει στην οικογένεια ATPασών τύπου-P. Αποτελείται από δύο υπομονάδες: την καταλυτική α-υπομονάδα και την β-υπομονάδα που απαιτείται για την κατάλληλη αναδίπλωση της α-υπομονάδας και για τον έλεγχο της δέσμευσης των ιόντων K+ και του κλεισίματος του διαύλου Η α-υπομονάδα φέρει 10 διαμεμβρανικούς α-έλικες (Μ1-10) και περιέχει δύο κύριες περιοχές: την περιοχή δέσμευσης του ATP (ATP-binding domain) και το τμήμα της μεμβράνης το οποίο εμπλέκεται στην πορεία διακίνησης του κατιόντος (cation- translocation pathway) Η περιοχή δέσμευσης του ATP συνδέεται απευθείας με τα διαμεμβρανικά τμήματα δρώντας κατ’ αυτόν τον τρόπο ως δομικός σύνδεσμος μεταξύ δύο λειτουργικών περιοχών και ταυτόχρονα παρέχει ‘σύζευξη’ μεταξύ της μεταφοράς των κατιόντων και της υδρόλυσης του ATP.
 και περιέχει δύο κύριες περιοχές: την περιοχή δέσμευσης του ATP (ATP-binding domain) και το τμήμα της μεμβράνης το οποίο εμπλέκεται στην πορεία διακίνησης του κατιόντος (cation- translocation pathway) Η περιοχή δέσμευσης του ATP συνδέεται απευθείας με τα διαμεμβρανικά τμήματα δρώντας κατ’ αυτόν τον τρόπο ως δομικός σύνδεσμος μεταξύ δύο λειτουργικών περιοχών και ταυτόχρονα παρέχει ‘σύζευξη’ μεταξύ της μεταφοράς των κατιόντων και της υδρόλυσης του ATP.")
42
ATPάση Na+/ K (Na+/K-ATPase)
● Καταναλώνει κυτταρική ενέργεια ● Διαμορφώνει διαβαθμίσεις συγκέντρωσης και ηλεκτρικού δυναμικού:Σε ένα κύτταρο η ATPάση Na+/K+ (Na+/K+-ATPase) δημιουργεί μια άνιση (μη-ισοδύναμη) κατανομή των ιόντων Na+ και K+ κατά πλάτος της μεμβράνης Υδρόλυση ενός μορίου 1 ATP μεταφέρει 2K+ μέσα και 3Na+ έξω ενάντια στις διαβαθμίσεις των συγκεντρώσεων τους
 δημιουργεί μια άνιση (μη-ισοδύναμη) κατανομή των ιόντων Na+ και K+ κατά πλάτος της μεμβράνης. Υδρόλυση ενός μορίου 1 ATP μεταφέρει 2K+ μέσα και 3Na+ έξω. ενάντια στις διαβαθμίσεις των συγκεντρώσεων τους.")
43
ATPάση Na+/K+ Κύκλος αντλίας ATPάσης Na+/K+
Διατηρεί την ανόμοια κατανομή των ιόντων Na+ και K+ κατά μήκος της κυτταρικής μεμβράνης διακινώντας 3Na+ και 2K+ ανά κάθε ένα ATP που υδρολύεται. Κατά τον κύκλο μεταφοράς των ιόντων, τα ιόντα ‘περικλείονται’ από τον μεταφορέα. Υφίσταται μετατροπή μεταξύ δύο δομικών διαμορφώσεων: E1 η οποία έχει υψηλή συγγένεια (high affinity) για κατιόντα Na+ και ATP και η E2, η οποία έχει υψηλή συγγένεια για κατιόντα K+. Απενεργοποιείται ειδικά από την ουαβαίνη (ouabain) ATPάση Na/K: Διαμόρφωση E2-P Κύκλος αντλίας ATPάσης Na+/K+
 για κατιόντα Na+ και ATP και η E2, η οποία έχει υψηλή συγγένεια για κατιόντα K+. Απενεργοποιείται ειδικά από την ουαβαίνη (ouabain) ATPάση Na/K: Διαμόρφωση E2-P. Κύκλος αντλίας ATPάσης Na+/K+")
44
Θεραπευτική δράση των καρδιοτονωτικών στεροειδών όπως η digitalis (παράγωγο της ουαβαίνης)
Η ATPάση Na/K είναι ένας υποδοχέας της digitalis και άλλων συναφών καρδιακών γλυκοσιδίων (cardiac glycosides) που χρησιμοποιούνται για την ενίσχυση του κτύπου της καρδιάς (heartbeat) Η αναστολή της ATPάσης Na/K από τα καρδιοτονωτικά στεροειδή όμοια με την ουαβαίνη οδηγεί στη μείωση της διαβάθμισης κατιόντων Na+ και της ενεργότητας του ανταλλάκτη κατιόντων Na+/Ca2+. Ως επακόλουθο, μειώνεται η ενδοκυτταρική συγκέντρωση των κατιόντων Ca2+ με αποτέλεσμα την καλύτερη συστολή του καρδιακού μυ. Ουαβαίνη (Ouabain)
 που χρησιμοποιούνται για την ενίσχυση του κτύπου της καρδιάς (heartbeat) Η αναστολή της ATPάσης Na/K από τα καρδιοτονωτικά στεροειδή όμοια με την ουαβαίνη οδηγεί στη μείωση της διαβάθμισης κατιόντων Na+ και της ενεργότητας του ανταλλάκτη κατιόντων Na+/Ca2+. Ως επακόλουθο, μειώνεται η ενδοκυτταρική συγκέντρωση των κατιόντων Ca2+ με αποτέλεσμα την καλύτερη συστολή του καρδιακού μυ. Ουαβαίνη (Ouabain)")
45
ATPάση Ca2+ του Σαρκοπλασματικού δικτύου (Ca2+-ATPase of Sarcoplasmic Reticulum)
Παίζει σημαντικό ρόλο στη χαλάρωση των μυών μεταφέροντας ιόντα Ca2+ που απελευθερώνονται πίσω στο SR. Η ATPάση Ca2+ έχει μια πρωτεϊνική υπομονάδα που αποτελείται από 10 διαμεμβρανικούς α-έλικες. Η ATPάση Ca2+ έχει υψηλή ομολογία με την ATPάση Na/K.

46
Η ATPάση Ca2+ του SR έχει 4 περιοχές:
● Η Περιοχή M ή περιοχή δέσμευσης στη μεμβράνη (Membrane-bound domain), που αποτελείται από 10 διαμεμβρανικά τμήματα ● Η περιοχή N δέσμευσης του νουκλεοτιδίου (N-Nucleotide-binding domain), όπου προσδένει η αδενίνη του ATP και ADP ● Η Περιοχή P (Phosphatase domain), η οποία περιέχει κατάλοιπα Asp, τα οποία φωσφορυλιώνονται κατά την υδρόλυση του ATP ● Η Περιοχή A (A-domain) η οποία είναι απαραίτητη για τις μεταβολές στη δομική διαμόρφωση της πρωτεΐνης μεταξύ των καταστάσεων E1 και E2
, που αποτελείται από 10 διαμεμβρανικά τμήματα. ● Η περιοχή N δέσμευσης του νουκλεοτιδίου. (N-Nucleotide-binding domain), όπου προσδένει η αδενίνη του ATP και ADP. ● Η Περιοχή P (Phosphatase domain), η οποία περιέχει κατάλοιπα Asp, τα οποία φωσφορυλιώνονται κατά την υδρόλυση του ATP. ● Η Περιοχή A (A-domain) η οποία είναι απαραίτητη για τις μεταβολές στη δομική διαμόρφωση της πρωτεΐνης μεταξύ των καταστάσεων E1 και E2.")
48
ATPάση H+/K+ (H+/K+-ATPase)
● Ανήκει στην οικογένεια των ATPασών τύπου-P (δομικά όμοια με την ATPάση Na+/K): Είναι ένα ετεροδιμερές που αποτελείται από 2 υπομονάδες, α και β. Η α-υπομονάδα αποτελείται από 10 διαμεμβρανικούς α-έλικες (Μ1-10) και φέρει την καταλυτική περιοχή ενώ η β-υπομονάδα εντοπίζεται μεταξύ των Μ9-Μ10. ● Μεταφέρει ένα ιόν H+ από τα οξυντικά κύτταρα σε ανταλλαγή με 1 ιόν K+ από τον αυλό του στομάχου: Εμπλέκεται στην έκκριση οξέος από το γαστρικό βλεννογόνο μεταφέροντας πρωτόνια (H+) έξω από το κύτταρο σε ανταλλαγή με κατιόντα K+ από τον εξωκυττάριο χώρο με αποτέλεσμα ο αυλός του στομάχου να γίνεται όξινος. ● Καθώς το έλκος του στομάχου και δωδεκαδακτύλου οφείλεται στην όξινη έκκριση, σημαίνει ότι η ATPάση H+/K+ αποτελεί φαρμακολογικό στόχο [Σιμετιδίνη, ανταγωνιστής των υποδοχέων ισταμίνης H2 ή omeprazole (Prilosec), αναστέλλει την αντλία] E1-P E2-P Ε1 δεσμεύει H+ και ATP και μετατρέπεται σε Ε1-P. Μεταβολή διαμόρφωσης σε Ε2-P που απελευθερώνει H+ και δεσμεύει K+. Αποφωσφορυλίωση Ε2-P, απελευθέρωση K+ και επιστροφή στη διαμόρφωση Ε1.
: Είναι ένα ετεροδιμερές που αποτελείται από 2 υπομονάδες, α και β. Η α-υπομονάδα αποτελείται από 10 διαμεμβρανικούς α-έλικες (Μ1-10) και φέρει την καταλυτική περιοχή ενώ η β-υπομονάδα εντοπίζεται μεταξύ των Μ9-Μ10. ● Μεταφέρει ένα ιόν H+ από τα οξυντικά κύτταρα σε ανταλλαγή με 1 ιόν K+ από τον αυλό του στομάχου: Εμπλέκεται στην έκκριση οξέος από το γαστρικό βλεννογόνο μεταφέροντας πρωτόνια (H+) έξω από το κύτταρο σε ανταλλαγή με κατιόντα K+ από τον εξωκυττάριο χώρο με αποτέλεσμα ο αυλός του στομάχου να γίνεται όξινος. ● Καθώς το έλκος του στομάχου και δωδεκαδακτύλου οφείλεται στην όξινη έκκριση, σημαίνει ότι η ATPάση H+/K+ αποτελεί φαρμακολογικό στόχο [Σιμετιδίνη, ανταγωνιστής των υποδοχέων ισταμίνης H2 ή omeprazole (Prilosec), αναστέλλει την αντλία] E1-P. E2-P. Ε1 δεσμεύει H+ και ATP και μετατρέπεται σε Ε1-P. Μεταβολή διαμόρφωσης σε Ε2-P που απελευθερώνει H+ και δεσμεύει K+. Αποφωσφορυλίωση Ε2-P, απελευθέρωση K+ και επιστροφή στη διαμόρφωση Ε1.")
49
Όξυνση του αυλού του στομάχου από τα οξυντικά κύτταρα του βλεννογόνου του
Ο στόμαχος περιέχει διάλυμα 0.1Μ HCl που εκκρίνεται από τα οξυντικά κύτταρα του βλεννογόνου του . Αυτά τα κύτταρα φέρουν μια ATPάση H+/K+ στην κορυφαία (apical) πλασματική μεμβράνη τους η οποία προεξέχει προς τον αυλό του στομάχου και δημιουργεί μια διαβάθμιση της συγκέντρωσης των [H+]: pH=1.0 στον αυλό του στομάχου σε σύγκριση με pH=7.0 στο κυτταρόπλασμα των οξυντικών κυττάρων. Αν τα οξυντικά κύτταρα αντλούσαν προς τα έξω ιόντα H+, τότε η απώλεια των πρωτονίων θα οδηγούσε στην αύξηση της συγκέντρωσης των [OH-] στο κυτοσόλιο και επομένως στην αύξηση του pH. Τα οξυντικά κύτταρα αποφεύγουν αυτήν την αύξηση του pΗ σε συνδυασμό με την όξυνση του αυλού του στομάχου χρησιμοποιώντας τους αντι-μεταφορείς Cl-/HCO3- στην βασοπλευρική μεμβράνη για να αντλήσουν την περίσσεια [OH-] από το κυτοσόλιο στο αίμα. Οι αντι-μεταφορείς ανιόντων Cl-/HCO3- ενεργοποιούνται με το υψηλό pH στο κυτοσόλιο. Η περίσσεια [OH-] αντιδράει με το CO2 που διαχέεται από το αίμα και σχηματίζουν HCO3-, με τη δράση της καρβονικής ανυδράσης. Το δικαρβονικό ιόν εξάγεται διαμέσου της βασοπλευρικής μεμβράνης με την ανταλλαγή ιόντων Cl- που στη συνέχεια εξέρχεται προς τον αυλό του στομάχου διαμέσου των διαύλων χλωρίου στην κορυφαία μεμβράνη. Για τη διατήρηση της ηλεκτρο ουδετερότητας, για κάθε ιόν Cl- που μεταφέρεται έξω από το κύτταρο, ένα ιόν K+ μεταφέρεται μέσα διαμέσου των διαύλων K+. Έτσι η περίσσεια ιόντων K που αντλείται προς τα μέσα από την ATPάση H+/K+ επιστρέφει στον αυλό του στομάχου, διατηρώντας έτσι τη φυσιολογική ενδοκυττάρια συγκέντρωση των ιόντων [K+]. Το καθαρό αποτέλεσμα είναι η ισοδύναμη έκκριση ιόντων H+ και Cl (HCl) στον αυλό του στομάχου, ενώ το pH στο κυτοσόλιο παραμένει ουδέτερο, και η περίσσεια HO- ως HCO3-, μεταφέρονται στο αίμα.
 πλασματική μεμβράνη τους η οποία προεξέχει προς τον αυλό του στομάχου και δημιουργεί μια διαβάθμιση της συγκέντρωσης των [H+]: pH=1.0 στον αυλό του στομάχου σε σύγκριση με pH=7.0 στο κυτταρόπλασμα των οξυντικών κυττάρων. Αν τα οξυντικά κύτταρα αντλούσαν προς τα έξω ιόντα H+, τότε η απώλεια των πρωτονίων θα οδηγούσε στην αύξηση της συγκέντρωσης των [OH-] στο κυτοσόλιο και επομένως στην αύξηση του pH. Τα οξυντικά κύτταρα αποφεύγουν αυτήν την αύξηση του pΗ σε συνδυασμό με την όξυνση του αυλού του στομάχου χρησιμοποιώντας τους αντι-μεταφορείς Cl-/HCO3- στην βασοπλευρική μεμβράνη για να αντλήσουν την περίσσεια [OH-] από το κυτοσόλιο στο αίμα. Οι αντι-μεταφορείς ανιόντων Cl-/HCO3- ενεργοποιούνται με το υψηλό pH στο κυτοσόλιο. Η περίσσεια [OH-] αντιδράει με το CO2 που διαχέεται από το αίμα και σχηματίζουν HCO3-, με τη δράση της καρβονικής ανυδράσης. Το δικαρβονικό ιόν εξάγεται διαμέσου της βασοπλευρικής μεμβράνης με την ανταλλαγή ιόντων Cl- που στη συνέχεια εξέρχεται προς τον αυλό του στομάχου διαμέσου των διαύλων χλωρίου στην κορυφαία μεμβράνη. Για τη διατήρηση της ηλεκτρο- ουδετερότητας, για κάθε ιόν Cl- που μεταφέρεται έξω από το κύτταρο, ένα ιόν K+ μεταφέρεται μέσα διαμέσου των διαύλων K+. Έτσι η περίσσεια ιόντων K+ που αντλείται προς τα μέσα από την ATPάση H+/K+ επιστρέφει στον αυλό του στομάχου, διατηρώντας έτσι τη φυσιολογική ενδοκυττάρια συγκέντρωση των ιόντων [K+]. Το καθαρό αποτέλεσμα είναι η ισοδύναμη έκκριση ιόντων H+ και Cl- (HCl) στον αυλό του στομάχου, ενώ το pH στο κυτοσόλιο παραμένει ουδέτερο, και η περίσσεια HO- ως HCO3-, μεταφέρονται στο αίμα.")
50
Δευτεροταγής Ενεργή Μεταφορά (Secondary active transport)
ΔG = RTln([C2]/[C1]) Αν C2<C1, τότε η ΔG είναι αρνητική και επομένως η μεταφορά διεξάγεται αυθόρμητα προς τη διαβάθμιση της συγκέντρωσης – Μεταφορείς, ιονοφόρα, πόροι, δίαυλοι Αν C2>C1, τότε η ΔG είναι θετική και επομένως απαιτείται ενέργεια, όπως ATP, για τη μεταφορά των μορίων ενάντια σε μια διαβαθμισμένη συγκέντρωση – ATPάσες τύπου-P, ATPάσες F1FO, Πολυμεταφορείς φαρμάκων, ATPάσες τύπου-V Αν δύο μόρια A και B έχουν μη-ισοδύναμη κατανομή κατά πλάτος της κυτταρικής μεμβράνης και η ΔG για το A είναι αρνητική και << από τη ΔG του B (ΔGΑ << ΔGB), τότε η μεταφορά του A ‘προς τη διαβάθμιση της συγκέντρωσης’ μπορεί να χρησιμοποιηθεί για τη μεταφορά του B ενάντια στη διαβάθμιση της συγκέντρωσης – Ανταλλάκτης Na+/Ca2+ (Na+/Ca2+-exchanger), Μεταφορέας Na+/Γλυκόζης (Na+,glucose transporter), Περμεάση λακτόζης (Lactose-permease)
 Αν C2<C1, τότε η ΔG είναι αρνητική και επομένως η μεταφορά διεξάγεται αυθόρμητα προς τη διαβάθμιση της συγκέντρωσης – Μεταφορείς, ιονοφόρα, πόροι, δίαυλοι. Αν C2>C1, τότε η ΔG είναι θετική και επομένως απαιτείται ενέργεια, όπως ATP, για τη μεταφορά των μορίων ενάντια σε μια διαβαθμισμένη συγκέντρωση – ATPάσες τύπου-P, ATPάσες F1FO, Πολυμεταφορείς φαρμάκων, ATPάσες τύπου-V. Αν δύο μόρια A και B έχουν μη-ισοδύναμη κατανομή κατά πλάτος της κυτταρικής μεμβράνης και η ΔG για το A είναι αρνητική και << από τη ΔG του B (ΔGΑ << ΔGB), τότε η μεταφορά του A ‘προς τη διαβάθμιση της συγκέντρωσης’ μπορεί να χρησιμοποιηθεί για τη μεταφορά του B ενάντια στη διαβάθμιση της συγκέντρωσης – Ανταλλάκτης Na+/Ca2+ (Na+/Ca2+-exchanger), Μεταφορέας Na+/Γλυκόζης (Na+,glucose transporter), Περμεάση λακτόζης (Lactose-permease)")
51
Γενικά χαρακτηριστικά της δευτεροταγούς ενεργής μεταφοράς
Χρησιμοποιούνται ευρέως για τη μεταφορά γλυκόζης, αμινοξέων, νευροδιαβιβαστών και άλλων συστατικών Ο ρυθμός μεταφοράς είναι σχετικά βραδύς. Σύγκριση : Δίαυλος ιόντων (Ion channel): ιόντα/sec ATP-εξαρτώμενη ενεργός μεταφορά (ATP-driven transport): ιόντα/sec Μεταφορέας (Transporter): ιόντα/sec Οι συν-μεταφορείς (Co-transporters) επιλεκτικά δεσμεύουν 2 ή περισσότερα μόρια για να μεταφέρουν; Η ταυτόχρονη δέσμευση είναι απαρραίτητη για την έναρξη της μεταφοράς. Αυτοί οι μεταφορείς δεν έχουν πόρο ή δομή διαύλου, και ο μηχανισμός μεταφοράς βασίζεται σε μια σειρά μεταβολών στη διαμόρφωση τους – γι’ αυτό και ο ρυθμός μεταφοράς είναι βραδύς
: ιόντα/sec. ATP-εξαρτώμενη ενεργός μεταφορά (ATP-driven transport): ιόντα/sec. Μεταφορέας (Transporter): ιόντα/sec. Οι συν-μεταφορείς (Co-transporters) επιλεκτικά δεσμεύουν 2 ή περισσότερα μόρια για να μεταφέρουν; Η ταυτόχρονη δέσμευση είναι απαρραίτητη για την έναρξη της μεταφοράς. Αυτοί οι μεταφορείς δεν έχουν πόρο ή δομή διαύλου, και ο μηχανισμός μεταφοράς βασίζεται σε μια σειρά μεταβολών στη διαμόρφωση τους – γι’ αυτό και ο ρυθμός μεταφοράς είναι βραδύς.")
52
Μεταφορείς συζευγμένοι με το σύστημα μεταφοράς ιόντων Na+
ενάντια σε διαβάθμιση συγκέντρωσης Αντι-μεταφορείς Na+/Ca2+ ενάντια σε διαβάθμιση συγκέντρωσης

53
Μεταφορά εντερικής γλυκόζης στο αίμα Ομόρροπος Συν-μεταφορέας
Βασοπλευρική ATPάση Na+/K+ δημιουργεί μια διαβάθμιση Na+ η οποία ωθεί τον ομόρροπο συν-μεταφορέα (Symporter) Ομόρροπος Συν-μεταφορέας Na+-Γλυκόζης Σύζευξη ομόρροπης συν-μεταφοράς 2 Na+ and 1 γλυκόζης Για τη μη-ιοντισμένη (μη-φορτισμένη) γλυκόζη ΔG=RTln([C2]/[C1]), η συν-μεταφορά με 2 Na+ δημιουργεί μια συγκέντρωση γλυκόζης μέσα στο κύτταρο κατά υψηλότερη 1η φάση: Ο συν-μεταφορέας 2Na+/1 Γλυκόζης (Glc) εισάγει Glc (& αμινοξέα) ενάντια στη συγκέντρωση της. Na+ αντλούνται στη συνέχεια έξω από το κύτταρο διαμέσου της βασοπλευρικής μεμβράνης με τη δράση της ATPάσης Na+/K+, με αποτέλεσμα τη διατήρηση της ενδοκυττάριας [Na+] . Αυτή η μεταφορά Na+/Glc ωθείται από την υδρόλυση του ATP. 2η Φάση: Η Glc (& αα) που συσσωρεύονται στο κυττοσόλιο με τη δράση των συν-μεταφορέων εξέρχονται στη συνέχεια από το κύτταρο προς τη διαβάθμιση της συγκέντρωσης της διαμέσου ενός μονο-μεταφορέα γλυκόζης στη βασοπλευρική μεμβράνη GLUT2 (GLUT1, ερυθροκύτταρα; GLUT2, εντερικά & ηπατικά; GLUT4, μυϊκά & λιπώδους ιστού).Το καθαρό αποτέλεσμα είναι η άντληση Na+, Glc (& αα) στον εξωκυττάριο χώρο και αίμα.
 Ομόρροπος Συν-μεταφορέας. Na+-Γλυκόζης. Σύζευξη ομόρροπης συν-μεταφοράς 2 Na+ and 1 γλυκόζης. Για τη μη-ιοντισμένη (μη-φορτισμένη) γλυκόζη ΔG=RTln([C2]/[C1]), η συν-μεταφορά με 2 Na+ δημιουργεί μια συγκέντρωση γλυκόζης μέσα στο κύτταρο κατά 1000 υψηλότερη. 1η φάση: Ο συν-μεταφορέας 2Na+/1 Γλυκόζης (Glc) εισάγει Glc (& αμινοξέα) ενάντια στη συγκέντρωση. της. Na+ αντλούνται στη συνέχεια έξω από το. κύτταρο διαμέσου της βασοπλευρικής μεμβράνης. με τη δράση της ATPάσης Na+/K+, με αποτέλεσμα. τη διατήρηση της ενδοκυττάριας [Na+] . Αυτή η. μεταφορά Na+/Glc ωθείται από την υδρόλυση του ATP. 2η Φάση: Η Glc (& αα) που συσσωρεύονται στο κυττοσόλιο με τη δράση των συν-μεταφορέων εξέρχονται στη. συνέχεια από το κύτταρο προς τη διαβάθμιση της συγκέντρωσης της διαμέσου ενός μονο-μεταφορέα γλυκόζης. στη βασοπλευρική μεμβράνη GLUT2 (GLUT1, ερυθροκύτταρα; GLUT2, εντερικά & ηπατικά; GLUT4, μυϊκά & λιπώδους ιστού).Το καθαρό αποτέλεσμα είναι η άντληση Na+, Glc (& αα) στον εξωκυττάριο χώρο και αίμα.")
Παρόμοιες παρουσιάσεις