Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
第七章 蛋白质的生物合成

2
一、概述 基因的遗传信息在转录过程中从DNA转移到mRNA,再由mRNA将这种遗传信息表达为蛋白质中氨基酸顺序的过程叫做翻译。
合成体系:20种氨基酸,mRNA、tRNA、核蛋白体、酶和因子,以及无机离子、ATP 、GTP 合成方向:N→C端。

3
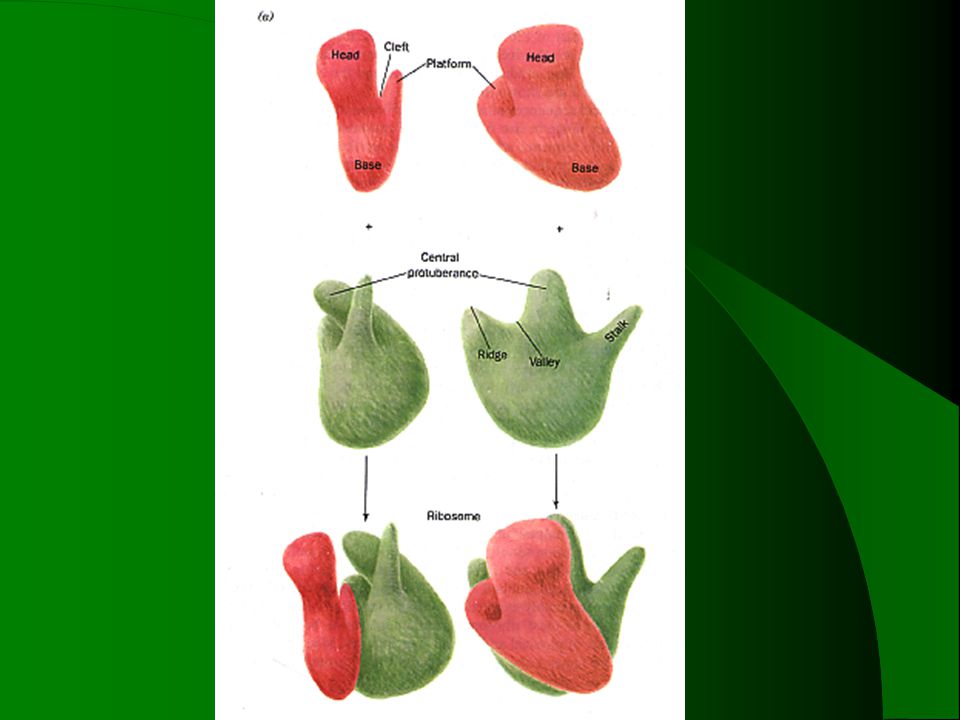
二、参与蛋白质合成的三类RNA及核糖体 1.rRNA 与蛋白质一起构成核糖体——蛋白质合成“工厂” 存在 核糖体结构组成 核糖体的基本功能
结合mRNA,在mRNA上选择适当的区域开始翻译 密码子(mRNA)和反密码子(tRNA)的正确配对 肽键的形成 存在 核糖体可游离存在,真核中,也可同内质网结合,形成粗糙的内质网。原核中,与mRNA形成串状——多核糖体
和反密码子(tRNA)的正确配对. 肽键的形成. 存在. 核糖体可游离存在,真核中,也可同内质网结合,形成粗糙的内质网。原核中,与mRNA形成串状——多核糖体.")
5
原核生物核糖体组成 真核生物核糖体组成

6
2. tRNA 结合氨基酸:一种氨基酸有几种tRNA携带,结合需要ATP供能,氨基酸结合在tRNA3‘-CCA的位置。
反密码子:每种tRNA的反密码子,决定了所带氨基酸能准确的在mRNA上对号入座 。 反密码子与mRNA的第三个核苷酸配对时,不严格遵从碱基配对原则

7
3. mRNA 携带着DNA的遗传信息,是多肽链的合成模板 在原核细胞内,存在时间短,在转录的同时翻译 在真核细胞内,较稳定
蛋白质合成时,mRNA结合于核糖体小亚基上,大亚 基结合带氨基酸的tRNA,tRNA的反密码子与mRNA密码子配对,ATP供能,合成蛋白质。

8
三、遗传密码子 1.密码子 为一个氨基酸编码进入蛋白质多肽链特定线性位置的三个核苷酸单位称为密码子(Coden)或三联体密码。 密码子的发现
统计学方法 人工合成仅由一种核苷酸组成的多聚核苷酸,推测由哪一种氨基酸合成的多肽 核糖体结合试验 年,Nirenberg用poly u加入C14标记的20种aa,仅有苯丙氨酸的寡肽,UUU=苯丙氨酸,用此法破译了全部密码,编出遗传密码表。

9
遗传密码

10
2. 遗传密码子的特点 无标点、不重叠 密码子是不重叠的,每个三联体中的三个核苷酸只编码一个氨基酸,核苷酸不重叠使用噬菌体x174中某些基因之间有重叠现象 简并(degeneracy) 几种密码子对应于相同一种氨基酸。这些密码子为同义密码子 通用性 绝大多数密码子对各种生物都适用,某些线粒体中遗传密码有例外 终止信号 UAG、UAA、UGA 起始信号 AUG(真核中起始为Met、原核中起始为fMet,翻译中间为Met)和氨酸的密码子(GUG)(极少出现)
和氨酸的密码子(GUG)(极少出现)")
11
四、蛋白质生物合成过程 1.氨基酸的活化 以mRNA为模板,氨基酸经活化获得的氨酰tRNA为原料,GTP、ATP供能,在核糖体中完成。
tRNA在氨基酰-tRNA 合成酶的帮助下,能够识别相应的氨基酸,并通过tRNA氨基酸臂的 3'-OH 与氨基酸的羧基形成活化酯-氨基酰-tRNA。 氨基酰-tRNA的形成是一个两步反应过程:第一步是氨基酸与 ATP 作用, 形成氨基酰腺嘌呤核苷酸; 第二步是氨基酰基转移到 tRNA 的 3'-OH 端上, 形成氨基酰-tRNA。

12
氨基酸活化图示

13
氨基酸活化的总反应式是: 氨基酰-tRNA 合成酶
氨基酸 + ATP + tRNA + H2O 氨基酰-tRNA + AMP + PPi 每一种氨基酸至少有一种对应的氨基酰-tRNA 合成酶。它既催化氨基酸与 ATP 的作用, 也催化氨基酰基转移到 tRNA。 氨基酰-tRNA 合成酶具有高度的专一性。 每一种氨基酰-tRNA 合成酶只能识别一种相应的 tRNA。 tRNA 分子能接受相应的氨基酸, 决定于它特有的碱基顺序, 而这种碱基顺序能够被氨基酰-tRNA 合成酶所识别。

14
氨基酸的活化

15
2.在核糖体上合成肽链 肽链的合成过程(以原核为例) 起始 延伸 终止与释放
氨基酰-tRNA通过反密码臂上的三联体反密码子识别mRNA上相应的遗传密码,并将所携带的氨基酸按mRNA遗传密码的顺序安置在特定的位置,最后在核糖体中合成肽链。 肽链的合成过程(以原核为例) 起始 延伸 终止与释放
 起始. 延伸. 终止与释放.")
16
肽链合成的起始 起始密码的识别 首先辨认出mRNA链上的起始点(AUG),核糖体小亚基上的16S rRNA和mRNA的SD序列(位于起始位点上游4-13个核苷酸)结合 N-甲酰甲硫氨酸-tRNA的活化形成 起始复合物的形成(图示)
")
17
肽链的延长 进位 (氨酰tRNA进入A位点) 参与因子:延长因子EFTu(Tu)、EFTs(Ts)、GTP、氨酰tRNA 肽链的形成
肽酰基从P位点转移到A位点,形成新的肽链 移位(translocase) 在移位因子(移位酶)EF-G的作用下,核糖体沿mRNA(5’-3’)作相对移动,使原来在A位点的肽酰-tRNA回到P位点
 在移位因子(移位酶)EF-G的作用下,核糖体沿mRNA(5’-3’)作相对移动,使原来在A位点的肽酰-tRNA回到P位点.")
18
核糖体移动方向 P位点 A位点

19
进位 核糖体移位 肽链的形成

20
延长过程中肽链的生成 肽基转移酶

21
肽链的延伸过程

22
肽链合成的终止与释放 识别mRNA的终止密码子,水解所合成肽链与tRNA间的酯键,释放肽链 R1识别UAA、UAG R2识别UAA、UGA
RR帮助P位点的tRNA残基脱落,而后核糖体脱落

23
多核糖体 在细胞内一条mRNA链上结合着多个核糖体,甚至可多到几百个。蛋白质开始合成时,第一个核糖体在mRNA的起始部位结合,引入第一个蛋氨酸,然后核糖体向mRNA的3’端移动一定距离后,第二个核糖体又在mRNA的起始部位结合,现向前移动一定的距离后,在起始部位又结合第三个核糖体,依次下去,直至终止。每个核糖体都独立完成一条多肽链的合成,所以这种多核糖体可以在一条mRNA链上同时合成多条相同的多肽链,这就大大提高了翻译的效率

24
五、真核细胞蛋白质合成的特点 核糖体为80S,由60S的大亚基和40S的小亚基组成 起始密码AUG 起始tRNA为Met-tRNA
起始复合物结合在mRNA 5’端AUG上游的帽子结构,真核mRNA无富含嘌呤的SD序列(除某些病毒mRNA外) 已发现的真核起始因子有近9种(eukaryote Initiation factor,eIF) eIF4A.eIF4E.P220复合物称为帽子结构结合蛋白复合物(CBPC) 肽链终止因子(EF1α EF1βγ )及释放因子(RF) 线粒体、叶绿体内蛋白质的合成同于原核细胞
 已发现的真核起始因子有近9种(eukaryote Initiation factor,eIF) eIF4A.eIF4E.P220复合物称为帽子结构结合蛋白复合物(CBPC) 肽链终止因子(EF1α EF1βγ )及释放因子(RF) 线粒体、叶绿体内蛋白质的合成同于原核细胞.")
26
蛋白质合成过程小结 肽链合成方向N C(同位素证明) 以mRNA的5’-3’方向阅读遗传密码 该合成过程是一个耗能过程
肽链的起始需要5ATP,延长时只需4ATP,合成一个n肽所需能量4×n+1 ATP,原核生物中,肽链的终止不需GTP,则合成n肽所需能量3×n+1

27
六、肽链合成后的“加工处理” N端改造 fMet的切除 信号肽(能透膜,进行蛋白质的锚定)的切除 氨基酸的修饰/改造
肽链内或肽链间的二硫键的形成、乙酰化、甲基化 氨基酸残基的修饰(Pro-OH/Cys-OH) 4.糖基化 (Asp、Ser、Thr、Asn) 5. 某些多肽要经特殊的酶切一段肽链后才有生物活性(如:胰岛素) 6. 高级结构的形成 在分子伴侣的协助下形成正确的结构 7.锚定(定位)
 4.糖基化 (Asp、Ser、Thr、Asn) 5. 某些多肽要经特殊的酶切一段肽链后才有生物活性(如:胰岛素) 6. 高级结构的形成 在分子伴侣的协助下形成正确的结构. 7.锚定(定位)")
28
七、蛋白质生物合成的调节 转录水平调节 转录后水平调节 翻译水平调节 蛋白质合成抑制剂: 抗生素类阻断剂
a. 链霉素、卡那霉素、新霉素等,主要抑制革兰氏阴性细菌蛋白质合成的三个阶段:①50S起始复合物的形成,使氨基酰tRNA从复合物中脱落;②在肽链延伸阶段,使氨基酰tRNA与mRNA错配;③在终止阶段,阻碍终止因了与核蛋白体结合,使已合成的多肽链无法释放,而且还抑制70S核糖体的介离。

29
干扰素对病毒蛋白合成的抑制 b.四环素和土霉素 c.氯霉素 d.白喉霉素(diphtheria toxin)
由白喉杆菌所产生的白喉霉素是真核细胞蛋白质合成抑制剂。它对真核生物的延长因子-2(EF-2)起共价修饰作用,生成EF-2腺苷二磷酸核糖衍生物,从而使EF-2失活,它的催化效率很高,只需微量就能有效地抑制细胞整个蛋白质合成,而导致细胞死亡 e.亚胺环己酮(放线菌酮) 只抑制真核60S亚基的肽酰转移酶活性 干扰素对病毒蛋白合成的抑制
起共价修饰作用,生成EF-2腺苷二磷酸核糖衍生物,从而使EF-2失活,它的催化效率很高,只需微量就能有效地抑制细胞整个蛋白质合成,而导致细胞死亡. e.亚胺环己酮(放线菌酮) 只抑制真核60S亚基的肽酰转移酶活性. 干扰素对病毒蛋白合成的抑制.")
30
本章重点: 1. 遗传密码的特性 2. 蛋白质生物合成的一般过程

Παρόμοιες παρουσιάσεις





 03088007 陈夏琴.>")
 班 周岑. Question 许多弱酸或弱碱,当 CaKa 或 CbKb 小于 10 -8 时,就不能用酸碱滴定法直接滴定 解决 : 增强酸碱强度 方法 : 可采用非水溶剂(包括有机溶剂或不 含水的无机溶剂)>")


 2 、研究串联谐振现象及电路参数对谐振特性 的影响。>")






 §6-2 集成模数转换器(ADC) §6-3 应用举例>")



