Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Θέμα 3ο Aρχή μεθόδων, πειραματικά πρωτοκόλλα, διδακτικά σχήματα
Μέθοδοι ανάλυσης μεταλλάξεων χωρίς προσδιορισμό των νουκλεοτιδικών αλληλουχιών Θέμα 3ο Aρχή μεθόδων, πειραματικά πρωτοκόλλα, διδακτικά σχήματα

2
Εισαγωγή Η μεταλλαγή στα γονίδια είναι υπεύθυνη για διάφορες γενετικές διαταραχές. Ο προσδιορισμός αυτών των μεταλλαγών απαιτεί νέες μεθόδους ανίχνευσης. Εντατική έρευνα κατά τη διάρκεια των ετών έχει οδηγήσει σε διάφορες νέες τεχνικές ανίχνευσης, μερικές από τις όποιες είναι εξειδικευμένες, όπως για παράδειγμα για τον καρκίνο του μαστού. Αυτές οι αναλύσεις γίνονται λαμβάνοντας υπόψη, το ιστορικό του καρκίνου, του ασθενή και το οικογενειακό ιστορικό

3
Μερικές από τις τεχνικές
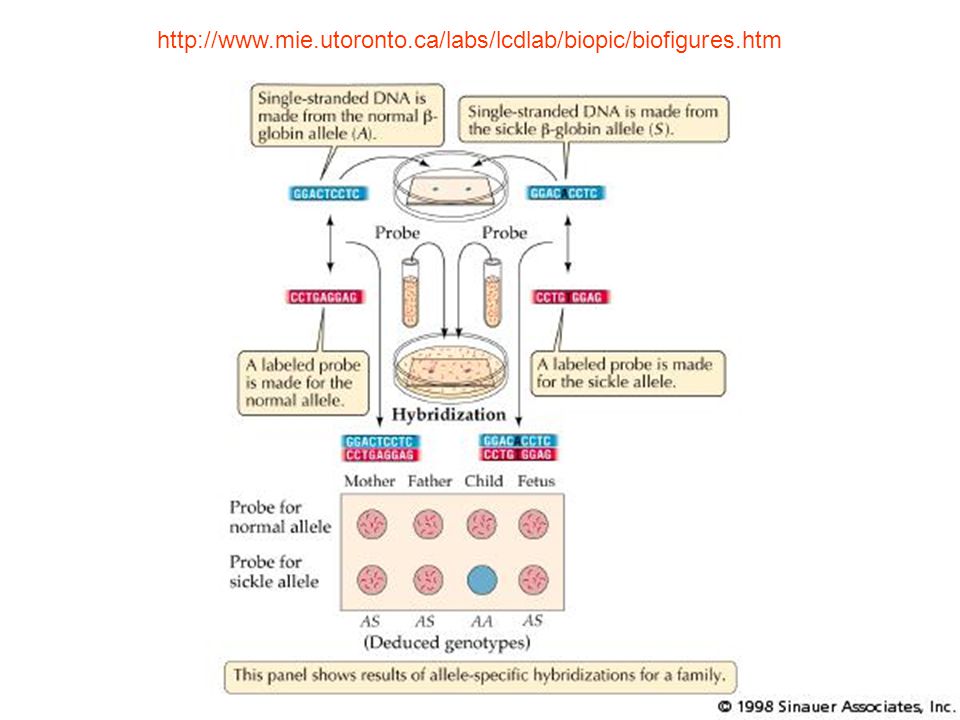
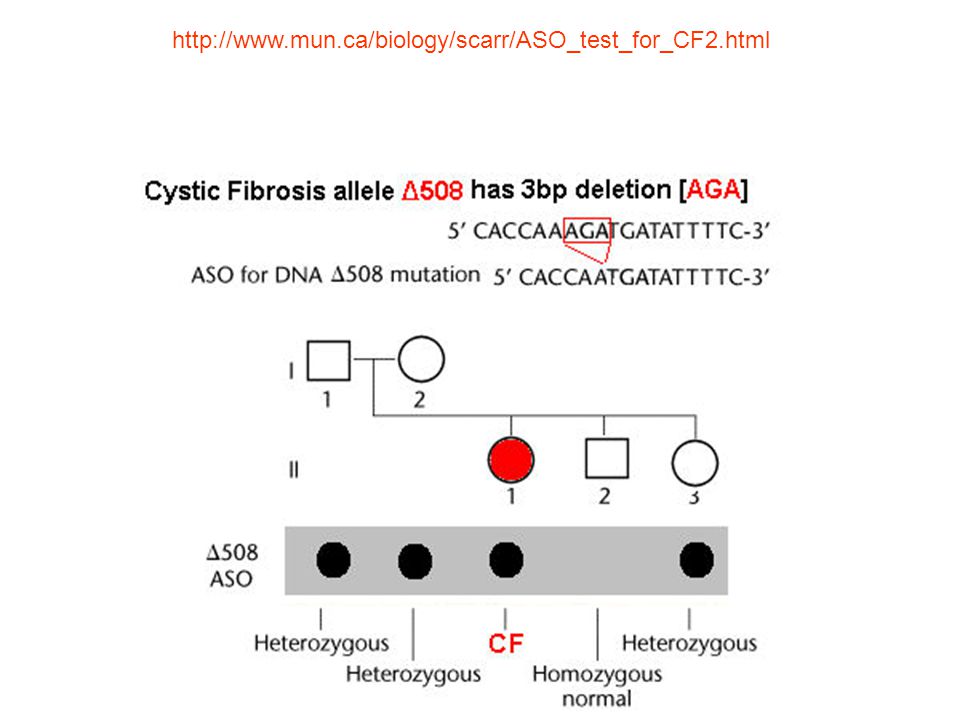
Allele Specific Oligonucleotides (ASO). Υβριδισμός με ειδικά ολιγονουκλεοτίδια. Protein Truncation Test (PTT). Πρωτεϊνική δοκιμή αποκοπής. Single Strand Conformational Polymorphism (SSCP). Πολυμορφισμός διαμόρφωσης μονόκλωνης αλυσίδας. Nucleotide sequencing. Αλληλούχηση νουκλεοτιδίων. Denaturing Gradient Gel Electrophoresis (DGGE). Ηλεκτροφόρηση σε πηκτή με κλίση αποδιατακτικού. Heteroduplex analysis. Ετερόδιπλου μορίου ανάλυση. DNA microarray technology. Τεχνολογία DNA μικρομωσαϊκών. Temperature gradient gel electrophoresis (TGGE). Ηλεκτροφόρηση σε πηκτή βαθμιδωτής θερμοκρασίας (παραλλαγή της DGGE). DENATURING HIGH-PERFORMANCE LIQUID CHROMATOGRAPHY (DHPLC) Υγρή χρωματογραφία με υψηλής απόδοσης αποδιάταξη Conformation Sensitive Gel Electrophoresis (CSGE). Ηλεκτροφόρηση σε πηκτή με ευαισθησία διαμόρφωσης Universal Heteroduplex Generator (UHG) Καθολική Γεννήτρια Ετερόδιπλου μορίου In situ υβριδισμός
. Υβριδισμός με ειδικά ολιγονουκλεοτίδια. Protein Truncation Test (PTT). Πρωτεϊνική δοκιμή αποκοπής. Single Strand Conformational Polymorphism (SSCP). Πολυμορφισμός διαμόρφωσης μονόκλωνης αλυσίδας. Nucleotide sequencing. Αλληλούχηση νουκλεοτιδίων. Denaturing Gradient Gel Electrophoresis (DGGE). Ηλεκτροφόρηση σε πηκτή με κλίση αποδιατακτικού. Heteroduplex analysis. Ετερόδιπλου μορίου ανάλυση. DNA microarray technology. Τεχνολογία DNA μικρομωσαϊκών. Temperature gradient gel electrophoresis (TGGE). Ηλεκτροφόρηση σε πηκτή βαθμιδωτής θερμοκρασίας (παραλλαγή της DGGE). DENATURING HIGH-PERFORMANCE LIQUID CHROMATOGRAPHY (DHPLC) Υγρή χρωματογραφία με υψηλής απόδοσης αποδιάταξη. Conformation Sensitive Gel Electrophoresis (CSGE). Ηλεκτροφόρηση σε πηκτή με ευαισθησία διαμόρφωσης. Universal Heteroduplex Generator (UHG) Καθολική Γεννήτρια Ετερόδιπλου μορίου. In situ υβριδισμός.")
4
Υβριδισμός με ειδικά ολιγονουκλεοτίδια
Υβριδισμός με ειδικά ολιγονουκλεοτίδια. (Allele Specific Oligonucleotides)(ASO) Αυτή η μέθοδος χρησιμοποιείται για να ανιχνεύσει μια ή περισσότερες γνωστές σημειακές μεταλλαγές στον πληθυσμό. Χρησιμοποιεί τους ελέγχους των νουκλεοτιδίων που είναι κοντά σε συγκεκριμένες και ιδιαίτερες ακολουθίες DNA. Αυτή είναι η απλούστερη και η λιγότερο ακριβής προσέγγιση από το να ψάχνουμε για οποιαδήποτε νέα μεταλλαγή στα μέλη μιας οικογένειας. Το σημαντικότερο μειονέκτημα αυτής της τεχνικής είναι ότι χρησιμοποιεί ενεργό ράδιο και μπορεί να προκαλέσει άλλες μεταλλαγές εάν αυτό είναι παρόν. το πρωτόκολλο της ASO, στηρίζεται στη διάκριση μεταξύ της διαφοράς δύο μορίων DNA σε μια βάση κατά την υβριδοποίηση. PCR φθορισμού εφαρμόζεται στα τμήματα που λέγονται SNP ακολουθίες. Μετά από αυστηρή υβριδοποίηση και πλύση, μετριέται η ένταση του φθορισμού για κάθε ένα SNP ολιγονουκλεοτίδιο
(ASO) Αυτή η μέθοδος χρησιμοποιείται για να ανιχνεύσει μια ή περισσότερες γνωστές σημειακές μεταλλαγές στον πληθυσμό. Χρησιμοποιεί τους ελέγχους των νουκλεοτιδίων που είναι κοντά σε συγκεκριμένες και ιδιαίτερες ακολουθίες DNA. Αυτή είναι η απλούστερη και η λιγότερο ακριβής προσέγγιση από το να ψάχνουμε για οποιαδήποτε νέα μεταλλαγή στα μέλη μιας οικογένειας. Το σημαντικότερο μειονέκτημα αυτής της τεχνικής είναι ότι χρησιμοποιεί ενεργό ράδιο και μπορεί να προκαλέσει άλλες μεταλλαγές εάν αυτό είναι παρόν. το πρωτόκολλο της ASO, στηρίζεται στη διάκριση μεταξύ της διαφοράς δύο μορίων DNA σε μια βάση κατά την υβριδοποίηση. PCR φθορισμού εφαρμόζεται στα τμήματα που λέγονται SNP ακολουθίες. Μετά από αυστηρή υβριδοποίηση και πλύση, μετριέται η ένταση του φθορισμού για κάθε ένα SNP ολιγονουκλεοτίδιο.")
7
Πρωτεϊνική δοκιμή αποκοπής (Protein truncation test) (PTT)
Η PTT είναι μια μέθοδος που ανιχνεύει τις μεταλλαγές που οφείλονται στη πρόωρη λήξη της μετάφρασης του mRNA. Ως αποτέλεσμα αυτού, το πρωτεϊνικό προϊόν είναι περικεκομμένο. Η περικεκομμένη πρωτεϊνη θα μπορούσε να προκύψει λόγω μιας μεταλλαγής που μετατοπίζει το πλαίσιο ανάγνωσης, μιας μη κωδικής μεταλλαγής και μιας μεταλλαγής θέσης του ματίσματος. Το πλεονέκτημα της μεθόδου PTT είναι το ότι μπορεί να ανιχνεύσει τις μεταλλαγές πολλών κιλοβάσεων. Εντούτοις, αυτή η μέθοδος δεν μπορεί να ανιχνεύσει τους πολυμορφισμούς, τις σιωπηλές μεταλλαγές και τις παρερμηνεύσιμες μεταλλαγές.

8
Σχηματική αναπαράσταση high-throughput στερεάς φάσης
protein truncation test (HTS-PTT)
")
9
Protein Truncation Test (PTT) voor BRCA1
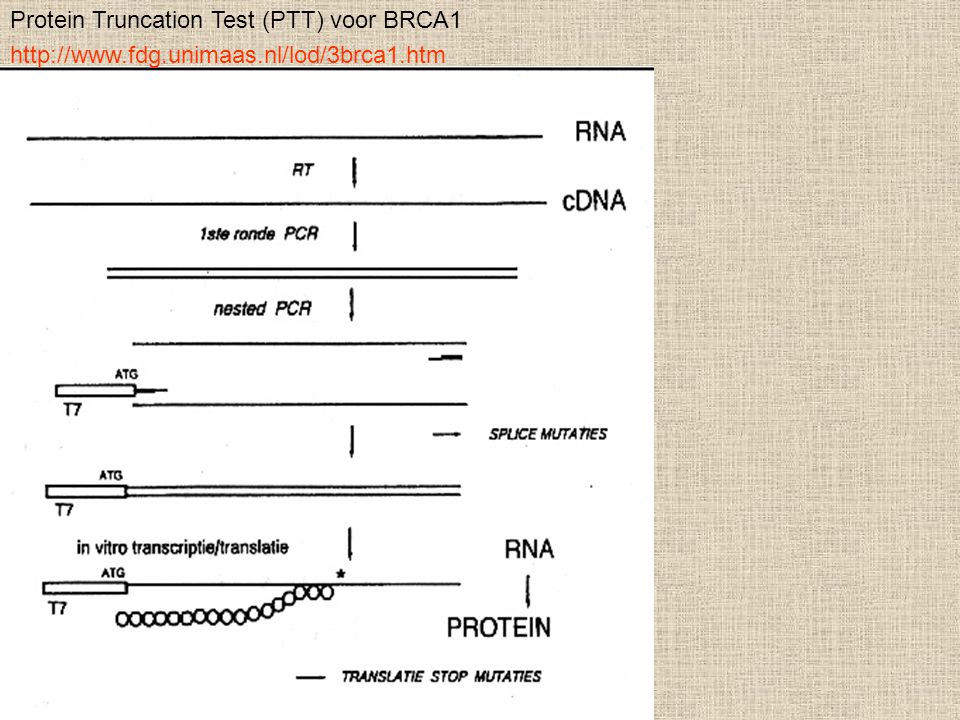
10
Πολυμορφισμός διαμόρφωσης μονόκλωνης αλυσίδας (Single Strand Conformational Polymorphism)(SSCP)
Αυτή η μέθοδος μπορεί να ανιχνεύσει μεταλλαγές σε βάσεις μονόκλωνης αλυσίδας όπως τις μεταλλαγές μετατόπισης του πλαισίου ανάγνωσης, μη κωδικές μεταλλαγές και κωδικές μεταλλαγές στον καρκίνο του μαστού. Η βασική αρχή αυτής της δοκιμής είναι το ότι το δίκλωνο DNA όταν αποδιατάσεται, παίρνει μια ειδική διαμόρφωση. Αυτή η διαμόρφωση είναι μοναδική και εξαρτάται από την αρχική ακολουθία νουκλεοτιδίων. Αυτή η μέθοδος είναι αρκετά ευαίσθητη για να ανιχνεύσει ακόμη και διαφορά στις μονόκλωνες νουκλεοτιδικές αλυσίδες που καταλαμβάνουν μια διαφορετική διαμόρφωση και όταν υποβάλλονται σε ηλεκτροφόρηση, η παραλλαγή των νουκλεοτιδίων καταλαμβάνει μια διαφορετική θέση

11
Η A ---> T μετάλλαξη σημειώνεται με μπλέ.
PCR-SSCP ανάλυση από φυσιολογικό και δρεπανοειδές κύτταρο β-γλοβίνης γονίδια. Η A ---> T μετάλλαξη σημειώνεται με μπλέ. Η περιοχή γύρω από την μετάλλαξη ενισχύεται με PCR και τα προϊόντα χωρίζονται σε ένα μη αποδιάταξης πήκτωμα πολυακρυλαμιδίου. Τα PCR προϊόντα από τη θέση του άγριου-τύπου και από τη θέση του δρεπανοκυττάρου θα μετακινηθούν διαφορετικά εξαιτίας των ειδικών προσαρμογών στις αλληλουχίες. Ομόζυγα άτομα φυσιολογικά θα εμφανίζουν 2 ζώνες όπως και τα ομόζυγα στην ασθένεια άτομα (με διαφορετικό όμως μήκος από τα φυσιολογικά). Άτομα ετερόζυγα για την ασθένεια θα εμφανίζουν 4 ζώνες.
. Άτομα ετερόζυγα για την ασθένεια θα εμφανίζουν 4 ζώνες.")
12
SSCP πηκτή από εδαφική μικροβιακή κοινότητα
Schmalenberger & Tebbe Mol. Ecol : Οι ζώνες κλωνοποιήθηκαν και τοποθετήθηκαν διαδοχικά και βρέθηκαν να είναι σύνθετες ομάδες ακολουθιών

13
SSCP Διαχειριστής για High Throughput απεικόνιση

14
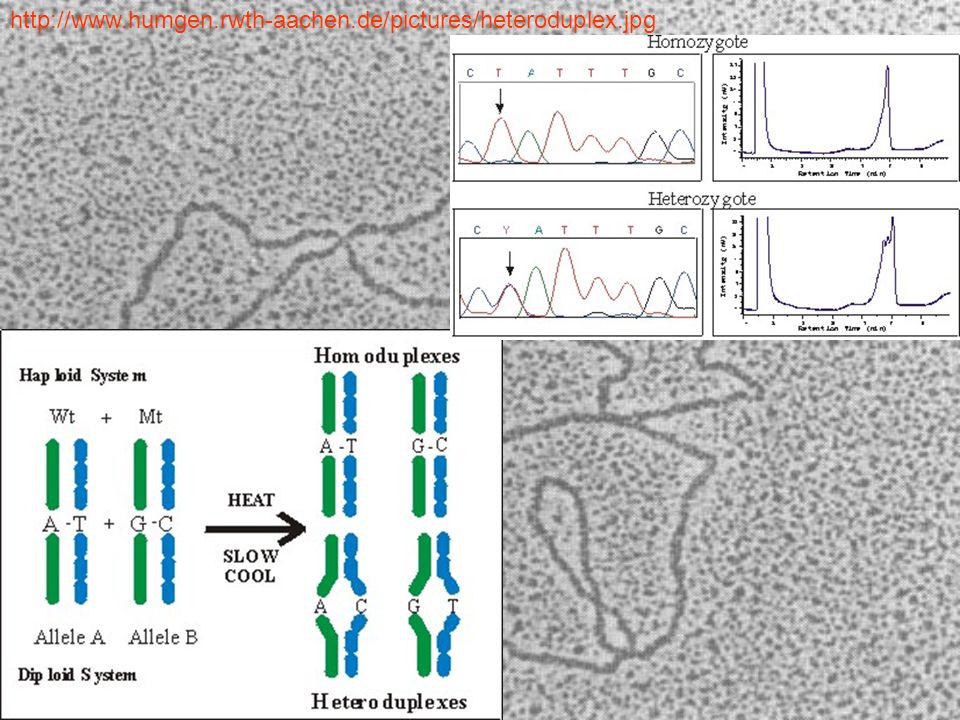
Heteroduplex ανάλυση (HDA)
Η αρχή πίσω από αυτήν την τεχνική είναι: οι κλώνοι του DNA μπορούν να χωριστούν μεταξύ τους με αποδιάταξη. Ομοίως, μπορούν επίσης να επαναδιαταχθούν, όταν οι κλώνοι βρεθούν πάλι μαζί, με άλλα λόγια, να επανενωθούν για να διαμορφώσουν ομόδιπλη δομή. Αλλά εάν υπάρχει μια μεταλλαγή στον έναν από τους κλώνους, το συνιστάμενο DNA θα είναι σε ετερόδιπλη κατάσταση. Μια τέτοια ιδιότητα του DNA μπορεί να χρησιμοποιηθεί καλύτερα για την ανίχνευση των σημειακών μεταλλαγών. Επιπλέον, οι ετερόδιπλοί DNA κλώνοι κινούνται πιο αργά επάνω σε πήκτωμα πολυακρυλαμιδίου όταν συγκρίνεται με το ομόδιπλο DNA

16
Τεχνολογία DNA μικροσυστοιχιών (chips) (DNA microarray technology)
Η σύνδεση των βάσεων ή η συμπληρωματικότητα των βάσεων είναι η αρχή πίσω από αυτή τη νέα τεχνολογία Οι πιθανές εφαρμογές της τεχνολογίας αυτής είναι η σκιαγράφηση της έκφρασης των γονιδίων και ο προσδιορισμός των ακολουθιών των γονιδίων (συμπεριλαμβανομένων των ακολουθιών που αφορούν τις μεταλλαγές) Σε αυτήν την μέθοδο, ένας μεγάλος αριθμός των DNA τμημάτων τοποθετείται σε μια φωτογραφική διαφάνεια γυαλιού. Τα τμήματα αφήνονται να υβριδώσουν ή να συνδεθούν με ιχνηθετημένο DNA (έλεγχοι), που υβριδιοποιούν το DNA στη φωτογραφική διαφάνεια γυαλιού Το ποσό της υβριδοποίησης αναλύεται έπειτα σε κάθε σημείο στη φωτογραφική διαφάνεια Στα γονίδια δίνεται ένα χρώμα όπου τα καλά υβριδοποιημένα χρωματίζονται κόκκινα και τα γονίδια που υβριδοποιούνται λιγότερο είναι χρωματισμένα πράσινα Θα μπορούσε να χρησιμοποιηθεί στον έλεγχο και την ταξινόμηση της ενίσχυσης των χρωμοσωμάτων και της έλλειψης αυτών, στους όγκους, στην ανακάλυψη φαρμάκων και την ανάπτυξή τους Επίσης η τεχνική εφαρμόζεται πρακτικά στην ανίχνευση των μεταλλαγών και την περαιτέρω έρευνα του μηχανισμού της ανάπτυξης των όγκων Δυνατότητα προσδιορισμού δεκάδων, εκατοντάδων ή χιλιάδων αλλαγών στα DNA, RNA, και, εσχάτως,στις πρωτεΐνες, σε 1 δοκιμασία (φυσικά, στο ίδιο δείγμα)
 Σε αυτήν την μέθοδο, ένας μεγάλος αριθμός των DNA τμημάτων τοποθετείται σε μια φωτογραφική διαφάνεια γυαλιού. Τα τμήματα αφήνονται να υβριδώσουν ή να συνδεθούν με ιχνηθετημένο DNA (έλεγχοι), που υβριδιοποιούν το DNA στη φωτογραφική διαφάνεια γυαλιού. Το ποσό της υβριδοποίησης αναλύεται έπειτα σε κάθε σημείο στη φωτογραφική διαφάνεια. Στα γονίδια δίνεται ένα χρώμα όπου τα καλά υβριδοποιημένα χρωματίζονται κόκκινα και τα γονίδια που υβριδοποιούνται λιγότερο είναι χρωματισμένα πράσινα. Θα μπορούσε να χρησιμοποιηθεί στον έλεγχο και την ταξινόμηση της ενίσχυσης των χρωμοσωμάτων και της έλλειψης αυτών, στους όγκους, στην ανακάλυψη φαρμάκων και την ανάπτυξή τους. Επίσης η τεχνική εφαρμόζεται πρακτικά στην ανίχνευση των μεταλλαγών και την περαιτέρω έρευνα του μηχανισμού της ανάπτυξης των όγκων. Δυνατότητα προσδιορισμού δεκάδων, εκατοντάδων ή χιλιάδων αλλαγών στα DNA, RNA, και, εσχάτως,στις πρωτεΐνες, σε 1 δοκιμασία (φυσικά, στο ίδιο δείγμα)")
17
Τι είναι οι Μικροδορυφόροι;
D2S123 TAGGCCACACACACACACACA κοινός εκκινητής • Mόνo, di, tri, τέτρα νουκλεοτιδικές επαναλήψεις • HNPCC – διαστολής/συστολής nl επαναλήψεις

18
Σύγκριση μεκλασσικές μοριακές μεθόδους
Βασική αρχή: συμπληρωματικότητα βάσεων νουκλεϊνικών οξέων Αντί για 1 ελέγχονται 1000άδες στόχοι ταυτόχρονα στο ίδιο δείγμα Απαραίτητη η ρομποτική (για την κατασκευή) και η βιοπληροφορική (για την ανάλυση).
 και η βιοπληροφορική (για την ανάλυση).")
19
Ηλεκτροφόρηση σε πηκτή βαθμιδωτής θερμοκρασίας (Temperature gradient gel electrophoresis) (TGGE)Ι
Είναι μια μορφή ηλεκτροφόρησης που μελετά τη συμπεριφορά των ουσιών κάτω από διαφορετικές θερμοκρασίες Η TGGE είναι μια από την οικογένεια των ηλεκτροφορητικών μεθόδων για το χωρισμό των νουκλεϊνικών οξέων όπως το DNA ή το RNA που στηρίζεται σις εξαρτώμενες από την θερμοκρασία αλλαγές στη δομή Η αρχική μέθοδος ήταν η DGGE, που είναι σχεδόν ίδια Η DGGE εφευρέθηκε από τον Leonard Lerman, ενώ ήταν καθηγητής στο SUNY Albany Ενώ ο ίδιος εξοπλισμός μπορεί να χρησιμοποιηθεί για την ανάλυση των πρωτεϊνών, που παραγάγουν παρόμοια στην όψη σχέδια, οι θεμελιώδεις αρχές είναι αρκετά διαφορετικές για τις πρωτεϊνες και τα νουκλεϊνικά οξέα

20
(TGGE)ΙΙ Είναι σαφές ότι η μέθοδος TGGE χρησιμοποιεί τη θερμότητα ως αρχικό μηχανισμό για το ξετίληγμα και την αποδιάταξη του DNA Αυτό που δεν είναι προφανές εντούτοις, είναι το πως αυτή η διαδικασία εμφανίζεται και το πώς το DNA μπορεί να αναλυθεί χρησιμοποιόντας αυτή την τεχνική Αυτό που είναι ίσως το πιό ενδιαφέρον κατά την εξέταση της διαδικασίας της αποδιάταξης με την TGGE μέθοδο είναι ότι εμφανίζεται σε τέτοιες συστηματικές διαδικασίες, που είναι δυνατόν να αναδημιουργήσει τα τμήματα μόλις αυτά χωριστούν Η εργασία με PCR ηλεκτροφόρηση των τμημάτων ξεκινά με μόρια διπλής έλικας Σε μια ορισμένη θερμοκρασία, το DNA αρχίζει να λειώνει, με κατάληξη σε μια δομή ομοιάζουσα με φούρκα Σε αυτήν την διαμόρφωση η απομάκρυνση επιβραδύνεται συγκρινόμενη με το εντελώς δίκλωνο τμήμα του DNA Έκτοτε η θερμοκρασία τήξης εξαρτάται σημαντικά από την ακολουθία των βάσεων, των τμημάτων του DNA του ίδιου μεγέθους αλλά οι διαφορετικές ακολουθίες μπορούν να χωριστούν Κατά συνέπεια η TGGE όχι μόνο χωρίζει τα μόρια, αλλά δίνει τις πρόσθετες πληροφορίες σχετικά με την τήξη, τη συμπεριφορά και την σταθερότητα

21
Σχήμα heteroduplex ανάλυσης στην TGGE
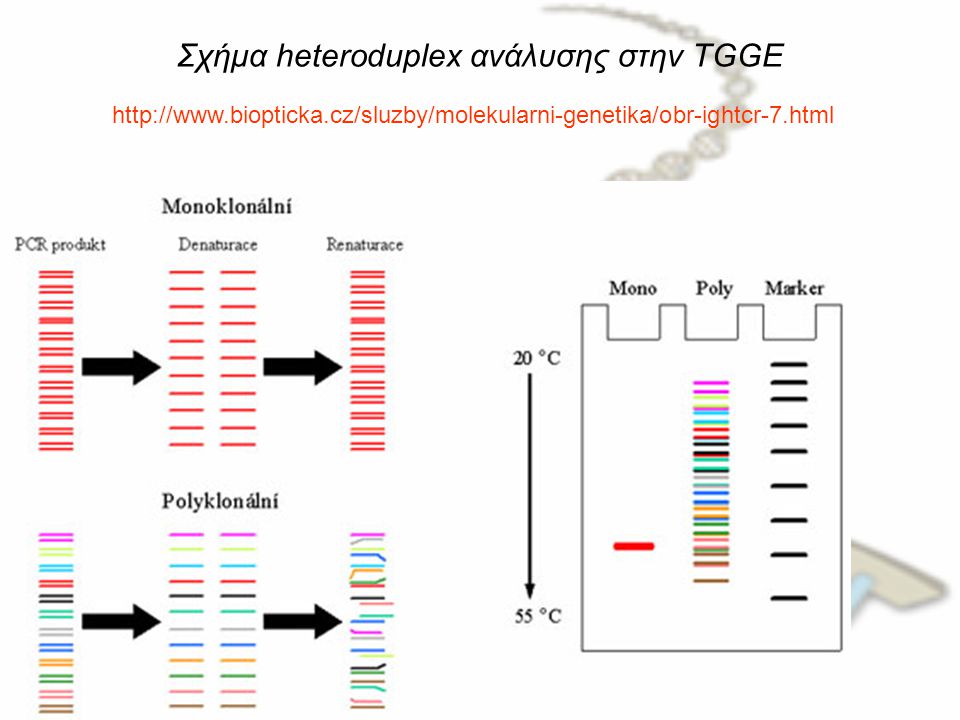
22
Ηλεκτροφόρηση σε πήκτωμα με κλίση αποδιατακτικού (DGGE)Ι
Για να καταλάβει κανείς την T/DGGE, υπάρχουν δύο θεμελιώδη σημεία: η δομή του DNA αλλάζει με τη θερμοκρασία, και το δεύτερο είναι πώς αυτές οι αλλαγές στη δομή έχουν επιπτώσεις στη μετακίνηση του DNA μέσω ενός πηκτώματος Αρχίζουμε με ένα μόριο δίκλωνου DNA με μερικές εκατοντάδες ζεύγη βάσεων σε μήκος Στη θερμοκρασία δωματίου, παρουσία τουλάχιστον ενός mM από άλας, η δίκλωνη μορφή είναι αρκετά σταθερή, και μπορούμε να εξετάσουμε το μόριο που είναι δύο σειρές στενά τυλιγμένες μεταξύ τους, ενεργά οι δύο άκρες του DNA είναι αρνητικά φορτισμένες (ανιόν) και στην παρουσία ενός ηλεκτρικού πεδίου, θα κινηθεί προς το θετικό ηλεκτρόδιο Ένα πήκτωμα είναι ένα μοριακό πλέγμα, με τις τρύπες κατά προσέγγιση με το ίδιο μέγεθος με τη διάμετρο του DNA Παρουσία του ηλεκτρικού πεδίου, το DNA θα προσπαθήσει να κινηθεί μέσω του πλέγματος, και για ένα δεδομένο σύνολο συνθηκών, η ταχύτητα της μετακίνησης είναι κατά προσέγγιση ανάλογη προς το μήκος του DNA μορίου - αυτή είναι η βάση για τον εξαρτώμενο από το μέγεθος χωρισμό στην τυποποιημένη ηλεκτροφόρηση.
 και στην παρουσία ενός ηλεκτρικού πεδίου, θα κινηθεί προς το θετικό ηλεκτρόδιο. Ένα πήκτωμα είναι ένα μοριακό πλέγμα, με τις τρύπες κατά προσέγγιση με το ίδιο μέγεθος με τη διάμετρο του DNA. Παρουσία του ηλεκτρικού πεδίου, το DNA θα προσπαθήσει να κινηθεί μέσω του πλέγματος, και για ένα δεδομένο σύνολο συνθηκών, η ταχύτητα της μετακίνησης είναι κατά προσέγγιση ανάλογη προς το μήκος του DNA μορίου - αυτή είναι η βάση για τον εξαρτώμενο από το μέγεθος χωρισμό στην τυποποιημένη ηλεκτροφόρηση.")
23
http://bccm.belspo.be/newsletter/17-05/bccm02.htm Αρχές της DGGE.
PCR ενισχυμένα προϊόντα κομμένα σε ίσα μήκη διαχωρίζονται ηλεκτροφορητικά με μια ακολουθιο-εξαρτώμενη μέθοδο βασιζόμενη στην περιεκτικότητά τους σε GC. Η αυξανόμενη κλίση των συστατικών αποδιάταξης κατά μήκος της πηκτής παραχωρεί τα δίκλωνα ενισχυμένα προϊόντα στο μονόκλωνο DNA μέσω των περιοχών που λειώνουν και έτσι θα μειωθεί η κινητικότητά τους (και η θέση τους στο πήκτωμα) Ένας GC- σφικτήρας συνδέεται στο 5’ άκρο σε έναν από τους PCR εκκινητές αποτρέποντας τα ενισχυμένα προϊόντα από την πλήρη αποδιάταξη. Διαφορετικές αλληλουχίες θα οδηγήσουν σε διαφορετική προέλευση των λειωμένων περιοχών και συνεπώς σε διαφορετικές θέσεις στην πηκτή που τα τμήματα του DNA σταματούν.
 Ένας GC- σφικτήρας συνδέεται στο 5’ άκρο σε έναν από τους PCR εκκινητές. αποτρέποντας τα ενισχυμένα προϊόντα από την πλήρη αποδιάταξη. Διαφορετικές αλληλουχίες θα οδηγήσουν σε διαφορετική προέλευση των λειωμένων περιοχών. και συνεπώς σε διαφορετικές θέσεις στην πηκτή που τα τμήματα του DNA σταματούν.")
24
Ηλεκτροφόρηση σε πήκτωμα με κλίση αποδιατακτικού (DGGE)ΙΙ
Καθώς κάποιος αυξάνει τη θερμοκρασία, οι δύο κλώνοι του DNA αρχίζουν να χωρίζουν και αυτό λειώνει Σε κάποια υψηλή θερμοκρασία, οι δύο κλώνοι θα χωρίσουν εντελώς Εντούτοις, σε κάποια ενδιάμεση θερμοκρασία, οι δύο κλώνοι θα χωριστούν εν μέρει, με μέρος του μορίου να είναι ακόμη δίκλωνο και ένα άλλο μέρος μονόκλωνο Κάποιος θα μπορούσε να το κάνει αυτό από το ένα τέλος, για να καταστήσει μία Υ διαμορφωμένη δομή με 3 άκρα, και από τις δύο άκρες για να κάνουν μια δομή με 4 άκρες, ή στη μέση για να κάνει μια φυσαλίδα Αυτό που κάνει την D/TGGE επωφελή είναι ότι η κινητικότητα του DNA μορίου μέσω του πηκτώματος μειώνεται δραστικά όταν διαμορφώνονται σε αυτό μερικώς λειωμένες δομές, και το σημαντικότερο, η ακριβής θερμοκρασία στο οποίο αυτό εμφανίζεται εξαρτάται από την αλληλουχία Κατά συνέπεια η D/TGGE προσφέρει μια μέθοδο εξαρτώμενη από την ακολουθία, ανεξαρτήτως μεγέθους για το χωρισμό των μορίων DNA.

25
Δομή μικροβιακής κοινότητας από Hopane - Degrading καλλιέργειες
LC JI WS ET TC

26
Ανάλυση των ζωνών από DGGE ενισχυμένων προφίλ A και B
Γειτονική RDP αλληλουχία (Sab) A B Γειτονική RDP αλληλουχία (Sab) ND Alcaligenes sp. (.992) P. azotoformans (.980) Burkholderia cepacia (.997) P. aeruginosa (.833) Frateuria aurantia (.839) P. echinoides (.771) Rhizobium/Brucella/ Ochrobactrum (.951) Rhizobium sp. (.948) Commomonas sp. (.955) ND P. rhodasiae (.927) Alcaligenes sp. (.920) Burkholderia cepacia (.639) P. alcaligenes (.696) Strentophomonas sp. (.973) P. echinoides (.808) Sphingomonas sp. (.797) Duganella zoogloides (.960) Acetobacter sp. (.752)
 A. B. Γειτονική RDP αλληλουχία (Sab) ND. Alcaligenes sp. (.992) P. azotoformans (.980) Burkholderia cepacia (.997) P. aeruginosa (.833) Frateuria aurantia (.839) P. echinoides (.771) Rhizobium/Brucella/ Ochrobactrum (.951) Rhizobium sp. (.948) Commomonas sp. (.955) ND. P. rhodasiae (.927) Alcaligenes sp. (.920) Burkholderia cepacia (.639) P. alcaligenes (.696) Strentophomonas sp. (.973) P. echinoides (.808) Sphingomonas sp. (.797) Duganella zoogloides (.960) Acetobacter sp. (.752)")
27
Υγρή χρωματογραφία με υψηλής απόδοσης αποδιάταξη (DENATURING HIGH-PERFORMANCE LIQUID CHROMATOGRAPHY) (DHPLC) Η DHPLC προσδιορίζει τις μεταλλαγές και τους πολυμορφισμούς με βάση την ανίχνευση του σχηματισμού ετερόδιπλων μορίων μεταξύ κακώς συνδυασμένων νουκλεοτιδίων στην διπλή έλικα κατά την PCR ενίσχυση του DNA Παραλλαγή ακολουθίας δημιουργεί μεικτό πληθυσμό ετερόδιπλων μορίων και ομόδιπλων κατά τη διάρκεια ανασυνδιασμού του άγριου τύπου και της DNA μετάλλαξης Όταν αυτός ο μικτός πληθυσμός αναλύεται με HPLC κάτω από θερμοκρασίες μερικής αποδιάταξης, τα ετερόδιπλα μόρια διαχωρίζονται με εκχύλιση από τη στήλη νωρίτερα από τα ομόδιπλα εξαιτίας της μειωμένης θερμοκρασία τήξης τους Η ανάλυση μπορεί να εκτελεσθεί σε δείγματα ενός ατόμου για να προσδιοριστεί η ετεροζυγωτία, ή σε μικτά δείγματα για να προσδιοριστεί η παραλλαγή των νουκλεοτιδίων μεταξύ των ατόμων

28
Heteroduplex διαμόρφωση. Άγριου-τύπου (Wt) και μεταλλαγμένου (Mt) PCR προϊόντα θερμαίνονται για να αποδιαταχθεί κάθε κλώνος και μετά αφήνονται να κρυώσουν αργά. Το αποτέλεσμα είναι ένα μείγμα πληθυσμών από τα αρχικά homoduplexes σύν τα heteroduplexes που περιέχουν αταίριαστες βάσεις.

29
Ηλεκτροφόρηση πηκτωμάτων με ευαισθησία διαμόρφωσης (CSGE)
Για την ανάλυση ετερόδιπλων μορίων με CSGE, τα προϊόντα της PCR ηλεκτροφορούνται Είναι σημαντικό να μην αποστειρωθεί το ρυθμιστικό διάλυμα TTE για να λάβει το βέλτιστο διαχωρισμό σε ετερόδιπλα μόρια και ομόδιπλα Ο βέλτιστος χρόνος πολυμερισμού είναι περίπου 1 ώρα. Τα προϊόντα της PCR που περιείχαν ετερόδιπλα μόρια αναμίγνύονται με ρυθμιστικό διάλυμα Τα δείγματα χωρίζονται με ηλεκτροφόρηση σε πρότυπες DNA sequencing συσκευές πηκτωμάτων Το πήκτωμα προ – ηλεκτροφορείται για 15 min, και τα δείγματα διαχωρίζονται στη θερμοκρασία δωματίου Μετά από την ηλεκτροφόρηση, το πήκτωμα χρωματίζεται στο γυάλινο πιάτο με βρωμίδιο του εθιδίου για 10 min ακολουθούμενο με ξέπλυμα στο νερό Ένας φορητός UV πυρσός χρησιμοποιείται για να απεικονίσει τις ζώνες Το σχετικό τμήμα από το πήκτωμα κόβεται, και μεταφέρεται σε ένα κομμάτι χαρτί αποτύπωσης Έπειτα απελευθερώνεται από το χαρτί επάνω στην επιφάνεια ενός διαεικονογράφου με το ξέπλυμα στο νερό Το πήκτωμα φωτογραφίζεται για την αποτύπωση των πηκτωμάτων

30
Ανάλυση αλληλούχισης με προσαρμογής - ευαίσθητη
Ανάλυση αλληλούχισης με προσαρμογής - ευαίσθητη ηλεκτροφόρηση σε πηκτή από τη μεταλλαγή που προκάλεσε το διαφορικό πρότυπο μετακίνησης. Οι 4 διάδρομοι στα αριστερά ανταποκρίνονται στην αλληλουχία του φυσιολογικού υποκινητή του γονιδίου Οι 4 διάδρομοι στα δεξιά είναι από δείγμα αιμοδοτών που εντοπίστηκε η μετατόπιση. Το βέλος δείχνει την παρουσία της T βάσης στη φυσιολογική αλληλουχία.

31
Καθολική Γεννήτρια Ετερόδιπλου μορίου (Universal Heteroduplex Generator)(UHG)
Η ανάλυση ετερόδιπλων μορίων σε πλάκες πηκτωμάτων (HDA), είναι μια δημοφιλής μέθοδος ανίχνευσης για τις γενετικές μεταλλαγές, που χρησιμοποιεί DNA τμήματα χαρακτηριστικά παραγμένα με PCR για να δημιουργήσει τα ομο - και ετερόδιπλα μόρια με τις διαμορφωτικές διαφορές και από τις αλληλουχίες-εξαρτώμενα ηλεκτροφορητικά προφίλ Χρήση (UHG) ενισχύει τις παραλλαγές που προκαλούνται από αντικαταστάσεις απλών-βάσεων. Η HDA-UHG σε πλάκες πηκτώματος τροποποιήθηκε σε μιά αποδοτική μέθοδο Η επίδραση της χρώσης των χρωστικών ουσιών TOPRO5 και YOPRO1 στην ανάλυση των ετερόδιπλων μορίων μελετήθηκε, καθώς επίσης και μέθοδοι ανίχνευσης της απορροφητικότητας στο φθορισμό από το υπεριώδες με λέιζερ (LIF) Επιπλέον, τα εμπλεκόμενα πολυμερή υδροξυαιθυλικής κυτταρίνης, μεθυλικής κυτταρίνης, και γραμμικού πολυακριλαμιδίου αξιολογήθηκαν χωριστά.
, είναι μια δημοφιλής μέθοδος ανίχνευσης για τις γενετικές μεταλλαγές, που χρησιμοποιεί DNA τμήματα χαρακτηριστικά παραγμένα με PCR για να δημιουργήσει τα ομο - και ετερόδιπλα μόρια με τις διαμορφωτικές διαφορές και από τις αλληλουχίες-εξαρτώμενα ηλεκτροφορητικά προφίλ. Χρήση (UHG) ενισχύει τις παραλλαγές που προκαλούνται από αντικαταστάσεις απλών-βάσεων. Η HDA-UHG σε πλάκες πηκτώματος τροποποιήθηκε σε μιά αποδοτική μέθοδο. Η επίδραση της χρώσης των χρωστικών ουσιών TOPRO5 και YOPRO1 στην ανάλυση των ετερόδιπλων μορίων μελετήθηκε, καθώς επίσης και μέθοδοι ανίχνευσης της απορροφητικότητας στο φθορισμό από το υπεριώδες με λέιζερ (LIF) Επιπλέον, τα εμπλεκόμενα πολυμερή υδροξυαιθυλικής κυτταρίνης, μεθυλικής κυτταρίνης, και γραμμικού πολυακριλαμιδίου αξιολογήθηκαν χωριστά.")
32
In situ υβριδισμός O in situ υβριδισμός, ο οποίος εφαρμόσθηκε για πρώτη φορά το 1969, αποτελεί πολύτιμη μέθοδο μοριακής βιολογίας για τον παθολoγοανατόμο, επειδή επιτρέπει τη μορφολογική εντόπιση της γενετικής πληροφορίας Eνώ οι κλασσικές τεχνικές της μοριακής βιολογίας, όπως η Southern blot και η PCR, πιστοποιούν απλώς την παρουσία μιας αλληλουχίας DNA ή RNA, ο in situ υβριδισμός προσδιορίζει επιπλέον σε ποιά και πόσα κύτταρα υπάρχει η αλληλουχία αυτή και σε ποιό υποκυτταρικό διαμέρισμα (κυτταρόπλασμα, πυρήνας) εντοπίζεται Eπιπρόσθετα, μας πληροφορεί εάν η παρουσία της αλληλουχίας αυτής σχετίζεται με συγκεκριμένες ανωμαλίες σε κυτταρικό και ιστικό επίπεδο H τεχνική του in situ υβριδισμού βασίζεται στη θεμελιώδη ιδιότητα των νουκλεϊκών οξέων να σχηματίζουν σύμφωνα με το νόμο της συμπληρωματικότητας των βάσεων σταθερά διμερή που ονομάζονται υβρίδια Tα τελευταία μπορεί να αποτελούνται από δύο αλυσίδες DNA ή συνδυασμό αλυσίδων RNA-DNA και RNA-RNA. O in situ υβριδισμός μπορεί να εφαρμοσθεί σε όλα τα είδη υλικού, όπως τα κυτταρικά επιχρίσματα και τις τομές παραφίνης και κρυοστάτη
 εντοπίζεται. Eπιπρόσθετα, μας πληροφορεί εάν η παρουσία της αλληλουχίας αυτής σχετίζεται με συγκεκριμένες ανωμαλίες σε κυτταρικό και ιστικό επίπεδο. H τεχνική του in situ υβριδισμού βασίζεται στη θεμελιώδη ιδιότητα των νουκλεϊκών οξέων να σχηματίζουν σύμφωνα με το νόμο της συμπληρωματικότητας των βάσεων σταθερά διμερή που ονομάζονται υβρίδια. Tα τελευταία μπορεί να αποτελούνται από δύο αλυσίδες DNA ή συνδυασμό αλυσίδων RNA-DNA και RNA-RNA. O in situ υβριδισμός μπορεί να εφαρμοσθεί σε όλα τα είδη υλικού, όπως τα κυτταρικά επιχρίσματα και τις τομές παραφίνης και κρυοστάτη.")
33
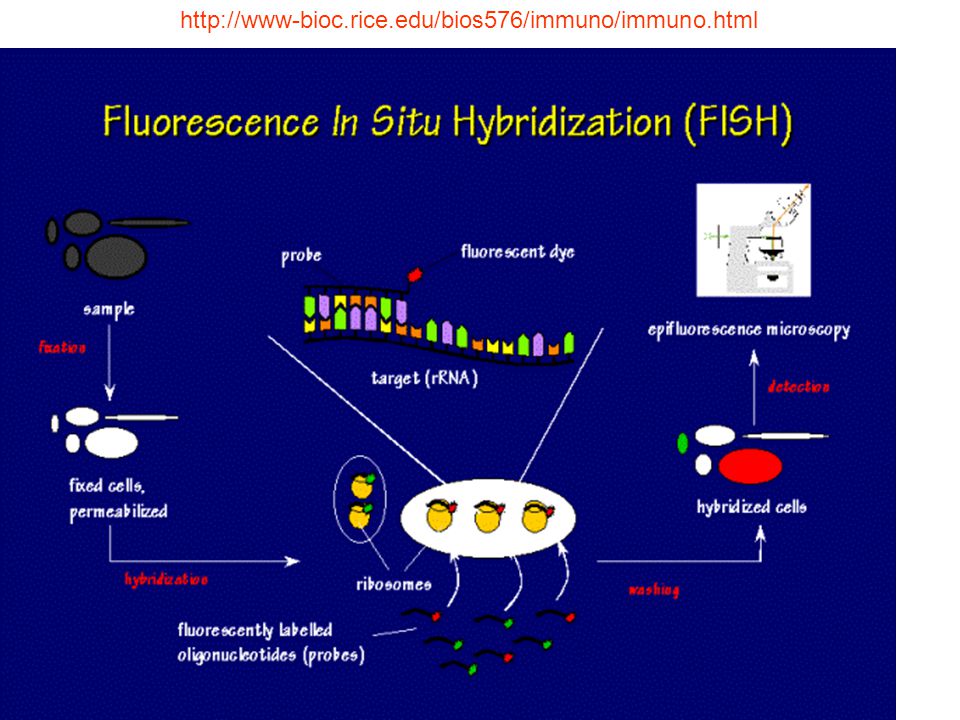
Fluorescence in situ hybridization (FISH)
")
34
Bασικά στάδια in situ υβριδισμού 1. Προετοιμασία του δείγματος
Aυτή περιλαμβάνει την επεξεργασία των κυττάρων του δείγματος με πρωτεολυτικά ένζυμα (πεψίνη ή πρωτεϊνάση K) ή μικροκύματα, ώστε να επιτευχθεί η διαπερατότητα κυτταροπλάσματος και πυρήνα στο δείκτη και η αλληλεπίδραση του με τις αναζητούμενες αλληλουχίες DNA ή RNA H πρωτεόλυση είναι απολύτως απαραίτητη, όταν η μονιμοποίηση του δείγματος έχει γίνει σε ουδέτερη φορμόλη, ενώ δεν απαιτείται σε μονιμοποίηση με αλκοολούχα μονιμοποιητικά Aποδιάταξη και υβριδισμός. Στο στάδιο αυτό επιτελείται απομάκρυνση των δύο αλύσων DNA του δείκτη και κυττάρου, ώστε να επιτευχθεί ακολούθως η σύνδεση τους στο κατάλληλο χημικό και θερμικό περιβάλλον Aνίχνευση αντίδρασης υβριδισμού.Mετά από τη μεθυβριδική έκπλυση των δειγμάτων ακολουθεί η ανίχνευση των σχηματισθέντων υβριδίων που γίνεται, ανάλογα με τη σήμανση του δείκτη, με αυτοραδιογραφία, ιστοχημικό-ανοσοενζυματικό μηχανισμό ή φθορισμό
 ή μικροκύματα, ώστε να επιτευχθεί η διαπερατότητα κυτταροπλάσματος και πυρήνα στο δείκτη και η αλληλεπίδραση του με τις αναζητούμενες αλληλουχίες DNA ή RNA. H πρωτεόλυση είναι απολύτως απαραίτητη, όταν η μονιμοποίηση του δείγματος έχει γίνει σε ουδέτερη φορμόλη, ενώ δεν απαιτείται σε μονιμοποίηση με αλκοολούχα μονιμοποιητικά. Aποδιάταξη και υβριδισμός. Στο στάδιο αυτό επιτελείται απομάκρυνση των δύο αλύσων DNA του δείκτη και κυττάρου, ώστε να επιτευχθεί ακολούθως η σύνδεση τους στο κατάλληλο χημικό και θερμικό περιβάλλον. Aνίχνευση αντίδρασης υβριδισμού.Mετά από τη μεθυβριδική έκπλυση των δειγμάτων ακολουθεί η ανίχνευση των σχηματισθέντων υβριδίων που γίνεται, ανάλογα με τη σήμανση του δείκτη, με αυτοραδιογραφία, ιστοχημικό-ανοσοενζυματικό μηχανισμό ή φθορισμό.")
35
Bασικά στάδια in situ υβριδισμού 2
Bασικά στάδια in situ υβριδισμού 2.Mέθοδοι ενίσχυσης ευαισθησίας in situ υβριδισμού H ενίσχυση της ευαισθησίας του in situ υβριδισμού μπορεί να γίνει σε επίπεδο αναζητούμενης αλληλουχίας προ του υβριδισμού ή σε επίπεδο ανίχνευσης της μετά τον υβριδισμό H πρώτη στρατηγική επιτυγχάνεται με την αύξηση του αριθμού των αντιγράφων DNA ή RNA με τη βοήθεια ενδοκυττάριας αλυσιδωτής αντίδρασης πολυμεράσης (in situ PCR) και ακολούθως, υβριδισμό των πολλαπλασιασθέντων αλληλουχιών με το δείκτη H δεύτερη στρατηγική επιτυγχάνεται με μεθόδους ενίσχυσης του σήματος μετά την αντίδραση υβριδισμού, η σπουδαιότερη από τις οποίες είναι αυτή που βασίζεται στην ενζυματική εναπόθεση τυραμιδίου (μέθοδος CARD) H μέθοδος αυτή επιτρέπει τη βελτίωση της ευαισθησίας της αντίδρασης 5-10 φορές, με αποτέλεσμα να καθιστά δυνατή την ανίχνευση ακόμη και ενός αντιγράφου νουκλεοτιδικής αλληλουχίας.
 και ακολούθως, υβριδισμό των πολλαπλασιασθέντων αλληλουχιών με το δείκτη. H δεύτερη στρατηγική επιτυγχάνεται με μεθόδους ενίσχυσης του σήματος μετά την αντίδραση υβριδισμού, η σπουδαιότερη από τις οποίες είναι αυτή που βασίζεται στην ενζυματική εναπόθεση τυραμιδίου (μέθοδος CARD) H μέθοδος αυτή επιτρέπει τη βελτίωση της ευαισθησίας της αντίδρασης 5-10 φορές, με αποτέλεσμα να καθιστά δυνατή την ανίχνευση ακόμη και ενός αντιγράφου νουκλεοτιδικής αλληλουχίας.")
37
Eφαρμογές του in situ υβριδισμού
Iδιαίτερα σημαντικές είναι οι εφαρμογές του in situ υβριδισμού στο κλινικό εργαστήριο και αφορούν: την ανίχνευση ιών τις μεταβολές γονιδίων σε επίπεδο πυρηνικού DNA (απώλειες, ενισχύσεις) αλλά και έκφρασης αυτών σε επίπεδο κυτταροπλασματικού mRNA τον προσδιορισμό δομικών και αριθμητικών χρωματοσωματικών ανωμαλιών
 αλλά και έκφρασης αυτών σε επίπεδο κυτταροπλασματικού mRNA. τον προσδιορισμό δομικών και αριθμητικών χρωματοσωματικών ανωμαλιών.")
38
Non-isotopic RNase cleavage assay (NIRCA) test of identity Ι
Η NIRCA ανάλυση υιοθετήθηκε για να προσδιορίσει διαφορές στις ακολουθίες μεταξύ RT-PCR προϊόντων και του κλώνου πλασμιδίων pRT14 Το RT-PCR προϊόν παρήχθη από FVB και C57BL/6 RNA από συκώτι με χρήση ολιγο-dT για να εκκινήσει την σύνθεση του πρώτου κλώνου Τα RT-PCR προϊόντα, το πλασμίδιο pRT14 DNA και γενωμικό DNA από FVB ποντίκια ενισχύθηκαν με χρήση εκκινητών που περιείχαν T7 (κωδικές) ή SP6 (αντικωδικές) εκκινητικές αλληλουχίες Τα τμήματα του εκκινητή (όλα 5´ 3´) τρέχουν από την 63 βάση στην 901
 ή SP6 (αντικωδικές) εκκινητικές αλληλουχίες. Τα τμήματα του εκκινητή (όλα 5´ 3´) τρέχουν από την 63 βάση στην 901.")
39
Non-isotopic RNase cleavage assay (NIRCA) test of identity ΙΙ
Κάθε ένα από τα προϊόντα της pcr αντίδρασης χρησιμοποιήθηκε ως πρότυπο για RNA παραγωγή με χρήση T7 και SP6 πολυμεράσες στις χωριστές αντιδράσεις Όλοι οι συνδυασμοί κωδικών και αντικωδικών προϊόντων υβριδοποιήθηκαν, και έγινε πέψη τους με δύο διαφορετικά μίγματα RNAασών Ο πρώτος ήταν ένα μίγμα από 1/300 RNAση 1 και 1/100 RNAse 2, και ο δεύτερος ήταν 1/100 RNAση 3 Μετά από την πέψη, οι αντιδράσεις διαχωρίστηκαν σε ένα 2% πήκτωμα αγαρόζης και φωτογραφήθηκαν

40
Non-isotopic RNase cleavage assay (NIRCA) πέψης προφίλ σε τρία χωρισμένα τμήματα
 πέψης προφίλ σε τρία χωρισμένα τμήματα.")
41
Tεχνολογία microchip H σπουδαιότερη επιστημονική επανάσταση των πρόσφατων χρόνων είναι η ανάπτυξη μικροτσίπ εφοδιασμένων με ολιγονουκλεοτίδια που επιτρέπουν ταχύτατες και ευρείας κλίμακας μελέτες γονιδίων Tα συστήματα αυτά με βάση την αντίδραση υβριδισμού επιτρέπουν την ανάλυση αλληλουχίας μεγάλου αριθμού επιλεγμένων γονιδίων και τον προσδιορισμό της έκφρασης αυτών σε επίπεδο mRNA H έκφραση επομένως χιλιάδων γονιδίων ταυτόχρονα μπορεί να μελετηθεί ποσοτικά και με ακρίβεια με τη βοήθεια ηλεκτρονικού υπολογιστή, εξασφαλίζοντας έτσι υψηλή ποιότητα και επαναλληψιμότητα

42
http://www. rsc. org/errorpage. asp. 404;http://www. rsc
ChemicalBiology/Volume/2006/8/Quick_PCR_microchip.asp

Παρόμοιες παρουσιάσεις




 AΠΟ ΦΥΤΙΚΑ ΚΥΤΤΑΡΑ>")









 - Γ' ΦΑΣΗ Αποθετήρια και Επιστημονικά Ηλεκτρονικά Περιοδικά Ανοικτής Πρόσβασης»>")




