Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Ανάλυση της αλληλουχίας των γονιδιωμάτων

2
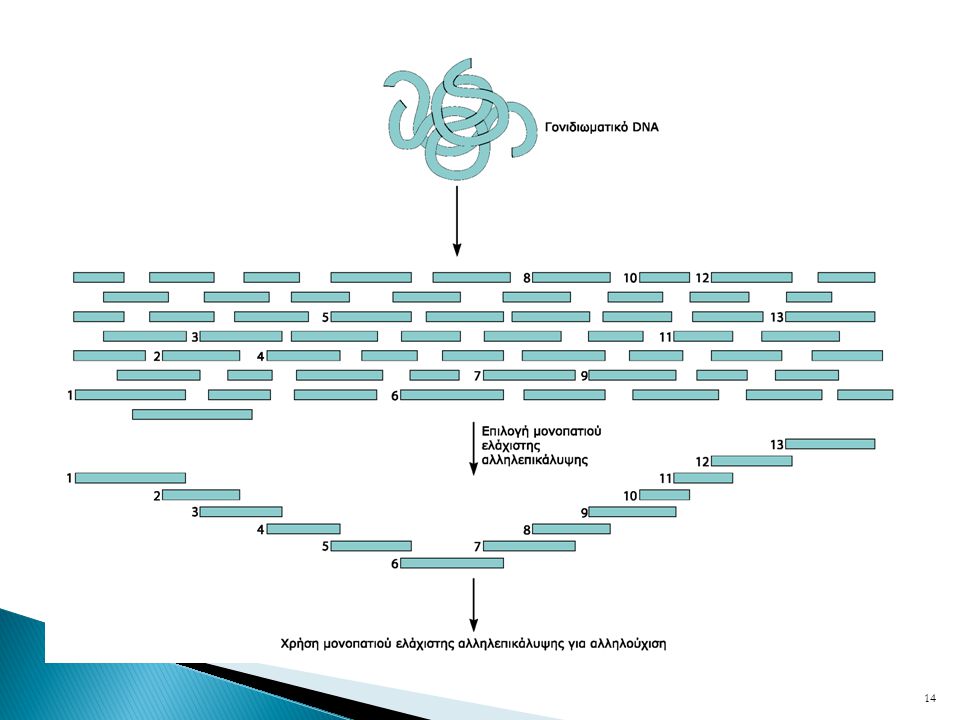
Μέθοδος συνεχών κλώνων / Συναρμολόγηση χαρτογραφημένων τμημάτων
Αλληλούχηση τυχαίων τμημάτων ολικού γονιδιώματος / Μέθοδος τυχαίας προσπέλασης

3
Μέθοδος τμηματικής κλωνοποίησης ολικού γονιδιώματος
Αλληλούχηση τυχαίων τμημάτων ολικού γονιδιώματος Μέθοδος τυχαίας προσπέλασης Whole genome shotgun method

4
Craig Venter, 1990, για βακτηριακά γονιδιώματα:
Πρόοδος τεχνολογίας αλληλούχησης Αυξανόμενη υπολογιστική ισχύς Δημιουργία κλώνων πολλών χιλιάδων μικρών τμημάτων (σε ισάριθμα πλασμίδια) Μαζική αλληλούχηση τυχαίων κλώνων Συναρμολόγηση των αναγνώσεων χωρίς πρότερη γνώση της θέσης τους στο γονιδίωμα
 Μαζική αλληλούχηση τυχαίων κλώνων. Συναρμολόγηση των αναγνώσεων χωρίς πρότερη γνώση της θέσης τους στο γονιδίωμα.")
5
Το παράδειγμα του Hemophilus influenzae
Γονιδίωμα μεγέθους 1,8Mbp Στις αρχές του 1990, το μεγαλύτερο γονιδίωμα που είχε αλληλουχηθεί: κυτταρομεγαλοϊός ~0,23Mbp Venter – πρόταση στρατηγικής τυχαίας προσπέλασης για την αλληλούχηση του H. influenzae: Σημαντικό στην ιατρική Μικρό γονιδίωμα Ανυπαρξία φυσικού χάρτη Περιεχόμενο G+C = ~38% (όσο και του ανθρώπου)
")
6
Τα βήματα Αποτελέσματα Κλασματοποίηση μέσω υπερήχων
Απομόνωση τμημάτων kb Κλωνοποίηση (19,687 κλώνοι) Αλληλούχηση με χρήση 14 πρωτότυπων αυτόματων συσκευών αλληλούχησης Συναρμολόγηση (30 ώρες σε υπολογιστή με RAM 512 Mbytes) Αποτελέσματα 11,631,485 bp, ≈6x το γονιδίωμα 140 contigs 50 χάσματα
 Αλληλούχηση με χρήση 14 πρωτότυπων αυτόματων συσκευών αλληλούχησης. Συναρμολόγηση (30 ώρες σε υπολογιστή με RAM 512 Mbytes) Αποτελέσματα. 11,631,485 bp, ≈6x το γονιδίωμα. 140 contigs. 50 χάσματα.")
7
Στρατηγικές προσπέλασης χασμάτων-1

8
Στρατηγικές προσπέλασης χασμάτων-2

9
Αποτελέσματα Συνεχής αλληλουχία 1.830.137bp
Εντοπισμός 1743 περιοχών που θα πρέπει να κωδικοποιούν πρωτεΐνες

10
ΕΙΚΟΝΑ 10.16: Χάρτης της αλληλουχίας του γονιδιώματος του Hemophilus influenzae.

11
Η προσέγγιση της τυχαίας προσπέλασης έχει καθιερωθεί για γονιδιώματα από 106 έως 3x109 bp.
Ένα μεγάλο εργαστήριο αλληλούχησης «διαβάζει» το γονιδίωμα ενός μικροβίου σε μία ημέρα Από τα πρώτα κίνητρα για την αλληλούχηση μικροβιακών γονιδιωμάτων ήταν οι εφαρμογές στην Ιατρική Όμως, η πρόκληση ήταν η αλληλούχηση του ανθρώπινου γονιδιώματος, μεγέθους 3x109 bp.

12
Μερικές ερωτήσεις-1: Γιατί κλασματοποίηση μέσω υπερήχων (και όχι ενζύμων περιορισμού); Για τη δημιουργία «τυχαίων» και «αλληλεπικαλυπτόμενων» τμημάτων DNA Ώστε να υπάρχουν όλες οι αλληλουχίες σε τμήματα των kb

13
Πέψη με ένζυμο περιορισμού
2 kb 0.5 kb 3.5 kb 1.8 kb 2 kb 1.8 kb Τμήματα εύρους kb

15
Μερικές ερωτήσεις-2: ≈ 6-10 x
Πόσους κλώνους χρειάζεται να αλληλουχήσουμε για να είμαστε σίγουροι ότι καλύψαμε το γονιδίωμα με τους αλληλεπικαλυπτόμενους κλώνους; ≈ 6-10 x

16
P0=e-m, όπου m είναι ο βαθμός κάλυψης
Σημασία του βαθμού κάλυψης (fold coverage) Ποια η πιθανότητα μια τυχαία βάση να μην αλληλουχηθεί; P0=e-m, όπου m είναι ο βαθμός κάλυψης Άρα, με κάλυψη 1x, P0= 2,73-1 = 0,37. δηλαδή, το 37% του γονιδιώματος δεν θα αλληλουχηθεί 6x, 1-P0=99,75% 10x, 1-P0=99,995% Ερώτημα: Πόσους πλασμιδιακούς κλώνους με μέσο μήκος ένθεσης 3kb χρειάζεστε για να είστε σίγουροι κατά 99,75% ότι έχετε κλωνοποιήσει μια συγκεκριμένη ανθρώπινη αλληλουχία;
 Ποια η πιθανότητα μια τυχαία βάση να μην αλληλουχηθεί; P0=e-m, όπου m είναι ο βαθμός κάλυψης. Άρα, με κάλυψη 1x, P0= 2,73-1 = 0,37. δηλαδή, το 37% του γονιδιώματος δεν θα αλληλουχηθεί. 6x, 1-P0=99,75% 10x, 1-P0=99,995% Ερώτημα: Πόσους πλασμιδιακούς κλώνους με μέσο μήκος ένθεσης 3kb χρειάζεστε για να είστε σίγουροι κατά 99,75% ότι έχετε κλωνοποιήσει μια συγκεκριμένη ανθρώπινη αλληλουχία;")
17
Μέθοδος των συνεχών κλώνων
Συναρμολόγηση χαρτογραφημένων τμημάτων Clone contig method

18
Κατακερματισμός του DNA σε κλάσματα μήκους έως 1.5 Mb (μερική πέψη)
Κλωνοποίηση κλασμάτων σε φορείς μεγάλης χωρητικότητας Χαρτογράφηση κλασμάτων στα χρωμοσώματα Καθορισμός της αλληλουχίας των μεγάλων κλασμάτων μέσω τυχαίας προσπέλασης (shotgun sequencing) και συναρμολόγησης Δημιουργία μικρότερων κλασμάτων Κλωνοποίηση σε πλασμιδιακούς φορείς Αλληλούχηση συζευγμένων άκρων (paired-end sequencing) Δημιουργία συναρμολογημάτων (contigs)
 και συναρμολόγησης. Δημιουργία μικρότερων κλασμάτων. Κλωνοποίηση σε πλασμιδιακούς φορείς. Αλληλούχηση συζευγμένων άκρων (paired-end sequencing) Δημιουργία συναρμολογημάτων (contigs)")
19
Κατακερματισμός του DNA σε κλάσματα μήκους έως 1.5 Mb (μερική πέψη)
Κλωνοποίηση κλασμάτων σε φορείς μεγάλης χωρητικότητας Χαρτογράφηση κλασμάτων στα χρωμοσώματα Καθορισμός της αλληλουχίας των μεγάλων κλασμάτων μέσω τυχαίας προσπέλασης (shotgun sequencing) και συναρμολόγησης Δημιουργία μικρότερων κλασμάτων Κλωνοποίηση σε πλασμιδιακούς φορείς Αλληλούχηση συζευγμένων άκρων (paired-end sequencing) Δημιουργία συναρμολογημάτων (contigs)
 και συναρμολόγησης. Δημιουργία μικρότερων κλασμάτων. Κλωνοποίηση σε πλασμιδιακούς φορείς. Αλληλούχηση συζευγμένων άκρων (paired-end sequencing) Δημιουργία συναρμολογημάτων (contigs)")
20
Φορείς μεγάλης χωρητικότητας
Το πλήθος των κλώνων που απαιτούνται προς αλληλούχηση είναι αντίστροφα ανάλογο με το μέγεθος των ενθέσεων γονιδιωματικού DNA που περιέχουν οι κλώνοι

21
Πιθανές επιπλοκές μεγάλων κλώνων YACs

22
Κατακερματισμός του DNA σε κλάσματα μήκους έως 1.5 Mb (μερική πέψη)
Κλωνοποίηση κλασμάτων σε φορείς μεγάλης χωρητικότητας Χαρτογράφηση κλασμάτων στα χρωμοσώματα Καθορισμός της αλληλουχίας των μεγάλων κλασμάτων μέσω τυχαίας προσπέλασης (shotgun sequencing) και συναρμολόγησης Δημιουργία μικρότερων κλασμάτων Κλωνοποίηση σε πλασμιδιακούς φορείς Αλληλούχηση συζευγμένων άκρων (paired-end sequencing) Δημιουργία συναρμολογημάτων (contigs)
 και συναρμολόγησης. Δημιουργία μικρότερων κλασμάτων. Κλωνοποίηση σε πλασμιδιακούς φορείς. Αλληλούχηση συζευγμένων άκρων (paired-end sequencing) Δημιουργία συναρμολογημάτων (contigs)")
23
Και πάλι χαρτογράφηση…
Φυσικοί χάρτες μέσω RE αποτυπωμάτων κλώνων με μεγάλες ενθέσεις Μερική πέψη με HindIII ή MboI για δημιουργία τμημάτων DNA ~170kb Βιβλιοθήκη BAC Πέψη απομονωμένων κλώνων με ένζυμο περιορισμού και ηλεκτροφόρηση Σύγκριση «αποτυπωμάτων» για την αποκάλυψη αλληλεπικαλυπτόμενων κλώνων

24
ΕΙΚΟΝΑ 11.9: Η ανάλυση αποτυπωμάτων χρησιμοποιείται για τη χαρτογράφηση και τη συναρμολόγηση κλώνων τεχνητών χρωμοσωμάτων βακτηρίων (BAC).
.")
25
Sequence-Tagged Sites
Ετικέτες μικρού μήκους χρησιμεύουν για τον προσδιορισμό του προσανατολισμού και της διαδοχικής σειράς γονιδιωματικών κλώνων Ενώ ένα STS αντιπροσωπεύει ένα φυσικό τμήμα DNA μοναδικό στο γονιδίωμα, δεν χρειάζεται ένας κλώνος αλλά μπορεί να παραχθεί μέσω μιας PCR Η PCR είναι ταχύτερη και πιο εύχρηστη από το στύπωμα κατά Southern

26
ΕΙΚΟΝΑ 11.10: Χαρτογράφηση με βάση την ανίχνευση STS σε κλώνους BAC.

27
Χαρτογράφηση με υβρίδια ακτινοβολημένων κυττάρων
1990: Οι Cox και Myers βελτίωσαν μια παλιά (Goss and Harris, 1975) μέθοδο δημιουργίας υβριδίων κυττάρων τρωκτικών και ανθρώπου
 μέθοδο δημιουργίας υβριδίων κυττάρων τρωκτικών και ανθρώπου.")
28
ΕΙΚΟΝΑ 11.11: Παραγωγή σειρών υβριδίων ακτινοβολημένων κυττάρων με σκοπό τη χαρτογράφηση του γονιδιώματος. Οι Goss & Harris δημιούργησαν υβριδικά κύτταρα με πολύ μικρά τμήματα ανθρώπινων χρωμοσωμάτων (ισχυρές δόσεις ακτίνων Χ). Στη συνέχεια προκάλεσαν σύντηξη με κύτταρα τρωκτικών και απομόνωσαν κυτταρικές σειρές κάθε μία των οποίων είχε ένα μόνο τμήμα ανθρώπινου γονιδιώματος. Τέλος, έκαναν μελέτες σύνδεσης στα υβριδικά κύτταρα, προσδιορίζοντας τη συχνότητα με την οποία δύο γονίδια (ή δείκτες) συγκληρονομούνταν (βρίσκονταν μαζί σε ένα σετ υβριδίων).
. Στη συνέχεια προκάλεσαν σύντηξη με κύτταρα τρωκτικών και απομόνωσαν κυτταρικές σειρές κάθε μία των οποίων είχε ένα μόνο τμήμα ανθρώπινου γονιδιώματος. Τέλος, έκαναν μελέτες σύνδεσης στα υβριδικά κύτταρα, προσδιορίζοντας τη συχνότητα με την οποία δύο γονίδια (ή δείκτες) συγκληρονομούνταν (βρίσκονταν μαζί σε ένα σετ υβριδίων).")
29
Οι Cox και Meyers δημιούργησαν ένα σετ 90 ακτινοβολημένων υβριδίων
Οι σειρές αυτές γονοτυπήθηκαν για την παρουσία ή απουσία χιλιάδων STSs Τα πρότυπα παρουσίας/απουσίας των STSs συγκρίθηκαν ώστε να προβλεφθεί η πιθανή σχετική διάταξη των STSs καθώς και η απόσταση μεταξύ τους

30
ΕΙΚΟΝΑ 11.12: Εφαρμογή της χαρτογράφησης μέσω ανάλυσης υβριδίων ακτινοβολημένων κυττάρων (RH, Radiation Hybrid) για τον προσδιορισμό της διάταξης ετικετών θέσεων αλληλουχίας (STS, Sequence-Tagged Sites). Αν η εμφάνιση δυο STSs (πχ 1 και 19) σε πολλές κυτταρικές σειρές είναι πιο συχνή από ό,τι θα αναμενόταν τυχαία, δείχνει ότι αυτοί οι δύο δείκτες έχουν μικρή απόσταση μεταξύ τους.
 σε πολλές κυτταρικές σειρές είναι πιο συχνή από ό,τι θα αναμενόταν τυχαία, δείχνει ότι αυτοί οι δύο δείκτες έχουν μικρή απόσταση μεταξύ τους.")
31
Χρησιμοποιήθηκαν δύο τέτοιες προσεγγίσεις:
Παρότι η διακριτική ικανότητα ενός τέτοιου χάρτη είναι χαμηλή, αποτελεί έναν ανεξάρτητο τρόπο προσδιορισμού της διάταξης τμημάτων DNA Χρησιμοποιήθηκαν δύο τέτοιες προσεγγίσεις: Ο ένας χάρτης είχε διακριτική ικανότητα ~1,5 εκατ bp Ο άλλος ~400,000 bp

32
A complete coverage, sequence-ready BAC contig in the chromosome 16p(13.1–11.2) region
55 BAC κλώνοι (πράσινο) συναρμολογήθηκαν στο contig βάσει restriction fingerprint analysis. Τα μεγέθη όλων των κλώνων έχουν καθοριστεί Συνολικά 76 BACs (λευκοί ρόμβοι) στο εσωτερικό των παραλληλογράμμων των κλώνων χαρτογραφήθηκαν με FISH και παρείχαν σημεία αγκυροβόλησης στα χρωμοσώματα
 συναρμολογήθηκαν στο contig βάσει restriction fingerprint analysis. Τα μεγέθη όλων των κλώνων έχουν καθοριστεί. Συνολικά 76 BACs (λευκοί ρόμβοι) στο εσωτερικό των παραλληλογράμμων των κλώνων χαρτογραφήθηκαν με FISH και παρείχαν σημεία αγκυροβόλησης στα χρωμοσώματα.")
33
Κατακερματισμός του DNA σε κλάσματα μήκους έως 1.5 Mb (μερική πέψη)
Κλωνοποίηση κλασμάτων σε φορείς μεγάλης χωρητικότητας Χαρτογράφηση κλασμάτων στα χρωμοσώματα Καθορισμός της αλληλουχίας των μεγάλων κλασμάτων μέσω τυχαίας προσπέλασης (shotgun sequencing) και συναρμολόγησης Δημιουργία μικρότερων κλασμάτων Κλωνοποίηση σε πλασμιδιακούς φορείς Αλληλούχηση συζευγμένων άκρων (paired-end sequencing) Δημιουργία συναρμολογημάτων (contigs)
 και συναρμολόγησης. Δημιουργία μικρότερων κλασμάτων. Κλωνοποίηση σε πλασμιδιακούς φορείς. Αλληλούχηση συζευγμένων άκρων (paired-end sequencing) Δημιουργία συναρμολογημάτων (contigs)")
34
ΕΙΚΟΝΑ 10.5: Δημιουργία πλασμιδιακής βιβλιοθήκης υποκλωνοποίησης για αλληλούχηση συζευγμένων άκρων.

35
Κατακερματισμός του DNA σε κλάσματα μήκους έως 1.5 Mb (μερική πέψη)
Κλωνοποίηση κλασμάτων σε φορείς μεγάλης χωρητικότητας Χαρτογράφηση κλασμάτων στα χρωμοσώματα Καθορισμός της αλληλουχίας των μεγάλων κλασμάτων μέσω τυχαίας προσπέλασης (shotgun sequencing) και συναρμολόγησης Δημιουργία μικρότερων κλασμάτων Κλωνοποίηση σε πλασμιδιακούς φορείς Αλληλούχηση συζευγμένων άκρων (paired-end sequencing) Δημιουργία συναρμολογημάτων (contigs)
 και συναρμολόγησης. Δημιουργία μικρότερων κλασμάτων. Κλωνοποίηση σε πλασμιδιακούς φορείς. Αλληλούχηση συζευγμένων άκρων (paired-end sequencing) Δημιουργία συναρμολογημάτων (contigs)")
36
ΕΙΚΟΝΑ 10.4: Η αλληλούχηση συζευγμένων άκρων (paired-end sequencing) συμβάλλει στην αποφυγή λαθών κατά τη συναρμολόγηση των αναγνώσεων αλληλουχίας.
 συμβάλλει στην αποφυγή λαθών κατά τη συναρμολόγηση των αναγνώσεων αλληλουχίας.")
37
ΕΙΚΟΝΑ 10.6: Η αλληλεπικάλυψη των κλώνων διευκολύνει την ταυτοποίηση της αλληλουχίας περιοχών επαναλαμβανόμενου DNA.

38
Βάδιση στο χρωμόσωμα-1

39
Βάδιση στο χρωμόσωμα-2

40
… τα χάσματα όμως παραμένουν…

41
EIKONA 10.14: Συναρμολόγηση της αλληλουχίας μιας περιοχής μεγάλου μήκους από πολλές αναγνώσεις αλληλουχίας μικρού μήκους.

42
ΕΙΚΟΝΑ 10.15: Κατά τη συναρμολόγηση των αλληλουχιών μπορεί να παρουσιαστούν αρκετά προβλήματα.

43
Πώς αλληλουχήθηκε το ανθρώπινο γονιδίωμα
Human Genome Projects ή Πώς αλληλουχήθηκε το ανθρώπινο γονιδίωμα Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

44
Λίγη ιστορία Αρχές 1980: Robert Sinsheimer (πρύτανης του UC Santa Cruz) απέτυχε να συγκεντρώσει αρκετά χρήματα για την κατασκευή του μεγαλύτερου τηλεσκοπίου στον κόσμο. Ίσως όμως να ήταν αρκετά για τη δημιουργία Κέντρου Γονιδιωματικής. Όμως, οι χρηματοδότες δεν πείσθηκαν. Μάιος 1985: συνάντηση σε Santa Cruz, με αυθεντίες όπως Bolstein, Gilbert, Lee Hood, Sulston, περί ενδιαφέροντος και δυνατότητας αλληλούχησης του ανθρώπινου γονιδιώματος. 1988: Ο Charles DeLisi, Υπουργός Ενέργειας των ΗΠΑ παραχωρεί κονδύλια του προϋπολογισμού για Πρόγραμμα Ανθρώπινου Γονιδιώματος Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007
 απέτυχε να συγκεντρώσει αρκετά χρήματα για την κατασκευή του μεγαλύτερου τηλεσκοπίου στον κόσμο. Ίσως όμως να ήταν αρκετά για τη δημιουργία Κέντρου Γονιδιωματικής. Όμως, οι χρηματοδότες δεν πείσθηκαν. Μάιος 1985: συνάντηση σε Santa Cruz, με αυθεντίες όπως Bolstein, Gilbert, Lee Hood, Sulston, περί ενδιαφέροντος και δυνατότητας αλληλούχησης του ανθρώπινου γονιδιώματος. 1988: Ο Charles DeLisi, Υπουργός Ενέργειας των ΗΠΑ παραχωρεί κονδύλια του προϋπολογισμού για Πρόγραμμα Ανθρώπινου Γονιδιώματος. Ανασυνδυασμένο DNA. Ακαδημαϊκές Εκδόσεις")
50
Οι συμμετέχοντες

51
Πού πραγματικά θα γίνει;

52
Δεδομένων των προβλημάτων…
Γιατί να μην επικεντρωθούν οι προσπάθειες στα μοναδικά τμήματα του γονιδώματος; Μέσω υβριδοποίησης θα μπορούσε να γίνει αρνητική επιλογή των υποκλώνων που περιέχουν επαναλαμβανόμενες αλληλουχίες και αφαίρεσή τους από τη «γραμμή» αλληλούχησης.

53
Αλληλούχηση cDNA 1990. Sydney Brenner: Η αλληλούχηση μόνο των cDNA θα μείωνε έως και 100 φορές τον όγκο της αλληλούχησης. Αλλά δεν έβρισκε χρηματοδότηση

54
1991. O Craig Venter (τότε στο ΝΙΗ) ανέφερε ότι είχε αλληλουχήσει ~600 ESTs από εγκεφαλικό ιστό
Ο Venter εγκατέλειψε το ΝΙΗ και ίδρυσε το The Institute of Genome Research (TIGR) όπου κατοχύρωνε τα ESTs Η απάντηση από μια ομάδα φαρμακευτικών εταιρειών (Merck and Co) που αλληλουχούσαν ESTs στο Washington U και τα διέθεταν σε δημόσιες βάσεις δεδομένων
 όπου κατοχύρωνε τα ESTs. Η απάντηση από μια ομάδα φαρμακευτικών εταιρειών (Merck and Co) που αλληλουχούσαν ESTs στο Washington U και τα διέθεταν σε δημόσιες βάσεις δεδομένων.")
55
Γενετικοί χάρτες Η δημιουργία γενετικών και φυσικών χαρτών κρίθηκε απαραίτητη για τη διευκόλυνση της συναρμολόγησης των αλληλουχιών Ως το 1979 είχαν εντοπιστεί ~200 γονίδια σε διάφορα ανθρώπινα χρωμοσώματα, συνδυάζοντας τη μελέτη οικογενών ανωμαλιών με την ανάλυση υβριδικών σωματικών κυττάρων Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

56
Εν αρχή ην… RFLPs Οι πρώτοι χάρτες (White et al, U of Utah and Helen Donis-Keller et al, Collaborative Research Inc) έφεραν ~400 RFLP δείκτες Όχι τυχαία κατανομή, άρα όχι ομοιόμορφη κάλυψη του γονιδιώματος 1990: χαρτογράφηση 2000 γονιδίων και 5000 τμημάτων DNA Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

57
Single Sequence Repeats
1993: Χάρτης 2000 χαρτογραφημένων CA επαναλήψεων από τη Genethon (Παρίσι) που κάλυπτε 90% του γονιδιώματος 1993: Χάρτης τρι- και τετρα-νουκλεοτιδικών επαναλήψεων από Cooperative Human Linkage Center (ΗΠΑ) 1994: Συνολικός χάρτης 5870 πολυμορφικών δεικτών με μέση απόσταση μεταξύ τους 0,7 cM (~700 kb) και από τις δύο εταιρείες μαζί Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007
 που κάλυπτε 90% του γονιδιώματος. 1993: Χάρτης τρι- και τετρα-νουκλεοτιδικών επαναλήψεων από Cooperative Human Linkage Center (ΗΠΑ) 1994: Συνολικός χάρτης 5870 πολυμορφικών δεικτών με μέση απόσταση μεταξύ τους 0,7 cM (~700 kb) και από τις δύο εταιρείες μαζί. Ανασυνδυασμένο DNA. Ακαδημαϊκές Εκδόσεις")
58
1993: Χάρτης 2000 χαρτογραφημένων CA επαναλήψεων από τη Genethon (Παρίσι) που κάλυπτε 90% του γονιδιώματος 1993: Χάρτης τρι- και τετρα-νουκλεοτιδικών επαναλήψεων από Cooperative Human Linkage Center (ΗΠΑ) 1994: Συνολικός χάρτης 5870 πολυμορφικών δεικτών με μέση απόσταση μεταξύ τους 0,7 cM (~700 kb) και από τις δύο εταιρείες μαζί Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007
 1994: Συνολικός χάρτης 5870 πολυμορφικών δεικτών με μέση απόσταση μεταξύ τους 0,7 cM (~700 kb) και από τις δύο εταιρείες μαζί. Ανασυνδυασμένο DNA. Ακαδημαϊκές Εκδόσεις")
59
ΕΙΚΟΝΑ 11.7: Γενετικός χάρτης του χρωμοσώματος 4 που σχεδιάστηκε με βάση μικροδορυφορικούς δείκτες.
Οι θέσεις των δεικτών αυτών και οι γενετικές τους αποστάσεις προσδιορί-στηκαν ακολουθώντας την κληρονόμηση των αλληλομόρφων κάθε δείκτη σε ένα καθορισμένο σύνολο μεγάλων οικογενειών. Ανασυνδυασμένο DNA

60
Αργοί ρυθμοί αλληλούχησης τα πρώτα 8 χρόνια (1990-98)
Δημιουργία χαρτών ήταν η κύρια προτεραιότητα Ποτέ πριν αλληλούχηση σε τέτοια κλίμακα νέες τεχνολογίες απειρία αντιμετώπισης περιοχών με επαναλήψεις Απαίτηση συντονισμού υψηλού επιπέδου εργαστηρίων από όλο τον κόσμο, εφαρμογή βιομηχανικών αρχών και προγραμματισμού παραγωγής Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

61
Συνεργασία αλλά και ανταγωνισμός
Εμπειρία μέσω αλληλούχησης μικρότερων γονιδιωμάτων (S. cerevisiae, C. elegans) Νέες χρηματοδοτήσεις Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007
 Νέες χρηματοδοτήσεις. Ανασυνδυασμένο DNA. Ακαδημαϊκές Εκδόσεις")
62
Θα υπάρχουν πατέντες; (ένα από τα πολλά ενδιαφέροντα των εταιρειών)
Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

63
Αρχές των Βερμούδων 1996: Διεθνής διάσκεψη στις Βερμούδες χρηματοδοτούμενη από το Wellcome Trust Αυτόματη δημοσίευση αλληλουχιών μέσα σε 24 ώρες σε δημόσιες βάσεις δεδομένων (δηλαδή πριν από τη δημοσίευση των σχετικών άρθρων) Διασφάλιζε τη διάθεση των αποτελεσμάτων σε όλη την επιστημονική κοινότητα Εμπόδιζε τις προσπάθειες κατοχύρωσης με διπλώματα ευρεσιτεχνίας
 Διασφάλιζε τη διάθεση των αποτελεσμάτων σε όλη την επιστημονική κοινότητα. Εμπόδιζε τις προσπάθειες κατοχύρωσης με διπλώματα ευρεσιτεχνίας.")
64
Πρόταση του Venter για διακοπή της επιχορήγησης του IHGSC

65
Το Wellcome Trust διπλασίασε την οικονομική ενίσχυση (και "απείλησε" ότι θα αναλάβει όλο το κόστος του προγράμματος σε περίπτωση που η κυβέρνηση των ΗΠΑ το εγκατέλειπε). Το αμερικανικό Κογκρέσο απέρριψε την πρόταση του Venter για διακοπή της επιχορήγησης του IHGSC Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

66
Κάλυψη των 2/3 του γονιδιώματος Όχι χρωματίνη
H Celera πρώτα εφάρμοσε την προσέγγιση της τυχαίας προσπέλασης στη Δροσόφιλα: Κατάτμησε και κλωνοποίησε το γονιδίωμα της δροσόφιλας σε τρεις διαφορετικές βιβλιοθήκες Αλληλούχησε και από τα δύο άκρα 1,5 εκατ κλώνους Ο Eugene Meyers ανέπτυξε νέους αλγόριθμους για τη συναρμολόγηση Κάλυψη των 2/3 του γονιδιώματος Όχι χρωματίνη Σημαντικός αριθμός χασμάτων Πρώτη πρόχειρη και πολύ χρήσιμη εφαρμογή της τυχαίας προσπέλασης

67
Για το ανθρώπινο γονιδίωμα η Celera:
χρησιμοποίησε ως DNA-μήτρα κλώνους με μικρές ενθέσεις παρήγαγε > 27 εκατ αναγνώσεις σε λιγότερο από 1 χρόνο Μέσα σε 2 χρόνια συναρμολόγησε την αλληλουχία Το IHGSC με τους Francis Collins (NIH), Michael Morgan (Wellcome Trust) και Αριστείδη Πατρινό (Υπουργείο Ενέργειας, ΗΠΑ) συντόνισαν 20 κέντρα γονιδιωματικής σε όλο τον κόσμο για τη συμπλήρωση των χασμάτων
, Michael Morgan (Wellcome Trust) και Αριστείδη Πατρινό (Υπουργείο Ενέργειας, ΗΠΑ) συντόνισαν 20 κέντρα γονιδιωματικής σε όλο τον κόσμο για τη συμπλήρωση των χασμάτων.")
68
Η εκεχειρία Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

69
Φεβρουάριος 2001 IHGSC σε Nature: Celera σε Science:
αλληλούχηση 29,298 χαρτογραφημένων κλώνων, κυρίως BAC 7,5 φορές κάλυψη 46% του γονιδιώματος υπό μορφή συναρμολογημάτων >100 kb Το μεγαλύτερο συναρμολόγημα 28,5 Μb χάσματα Celera σε Science: Κάλυψη: 5,1x. Σε συνδυασμό με IHGSC: 12,6x 49% του γονιδιώματος υπό μορφή συναρμολογημάτων >100 kb Το μεγαλύτερο συναρμολόγημα 2 Mb χάσματα Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

70
ΠΙΝΑΚΑΣ 11.1: Στη Διεθνή Κοινοπραξία Αλληλούχησης του Ανθρώπινου Γονιδιώματος (International Human Genome Sequencing Consortium) συμμετείχαν 20 διαφορετικά κέντρα από έξι χώρες. 1. SC: Wellcome Trust Sanger Institute (Hinxton, UK) 2. WUGSC: Washington U Genome Sequencing Center (St Louis, Missouri) 3. WIBR: Whitehead Institute of Biomedical Research (Cambridge, MA) 4. JGI/SHGC: US DOE Joint Genome Institute and Stanford Human Genome Center (Walnut Creek and Stanford, CA) 5. BCM: Baylor College of Medicine Human Genome Sequencing Center (Houston, Texas) Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007
 2. WUGSC: Washington U Genome Sequencing Center (St Louis, Missouri) 3. WIBR: Whitehead Institute of Biomedical Research (Cambridge, MA) 4. JGI/SHGC: US DOE Joint Genome Institute and Stanford Human Genome Center (Walnut Creek and Stanford, CA) 5. BCM: Baylor College of Medicine Human Genome Sequencing Center (Houston, Texas) Ανασυνδυασμένο DNA. Ακαδημαϊκές Εκδόσεις")
71
Τα επιτεύγματα Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

72
Date achieved Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

73
Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

74
ΕΙΚΟΝΑ 11.13: Η διαδικασία ολοκλήρωσης της αλληλούχησης του γονιδιώματος.
Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

75
Ακόμα εντυπωσιακότερη, όμως, είναι η μείωση των χασμάτων
ΠΙΝΑΚΑΣ 11.2: Η ολοκληρωμένη αλληλουχία του ανθρώπινου γονιδιώματος του IHGSC είναι σημαντικά βελτιωμένη σε σχέση με την πρόχειρη αλληλουχία. Μέτρο πιθανότητας (probability score): η πιθανότητα μιας νουκλεοτιδικής θέσης να έχει ταυτοποιηθεί με λάθος τρόπο Ακόμα εντυπωσιακότερη, όμως, είναι η μείωση των χασμάτων Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007
: η πιθανότητα μιας νουκλεοτιδικής θέσης να έχει ταυτοποιηθεί με λάθος τρόπο. Ακόμα εντυπωσιακότερη, όμως, είναι η μείωση των χασμάτων. Ανασυνδυασμένο DNA. Ακαδημαϊκές Εκδόσεις")
76
Η έμφαση στην ακρίβεια άξιζε τα λεφτά της;
Οι ανακρίβειες θα καθιστούσαν δύσκολο τον εντοπισμό γονιδίων Αν η συχνότητα σφάλματος ήταν μεγαλύτερη των SNPs (~1/1000) δεν θα μπορούσαμε να τα διακρίνουμε Οι ανακρίβειες θα καθιστούσαν δύσκολες τις συγκρίσεις με άλλους οργανισμούς Η ακρίβεια είναι απαραίτητη για τη διάκριση διπλασιασμένων γονιδίων και μελών μεγάλων οικογενειών γονιδίων
 δεν θα μπορούσαμε να τα διακρίνουμε. Οι ανακρίβειες θα καθιστούσαν δύσκολες τις συγκρίσεις με άλλους οργανισμούς. Η ακρίβεια είναι απαραίτητη για τη διάκριση διπλασιασμένων γονιδίων και μελών μεγάλων οικογενειών γονιδίων.")
77
Τίνος το DNA; IHGSC Celera Ανώνυμοι δότες, αδυναμία ταυτοποίησής τους
Καταστροφή στοιχείων των δοτών 70% από ένα δείγμα, 30% από άλλα ανώνυμα άτομα Celera 21 δείγματα από 5 διαφορετικές φυλές → χρήσιμες πληροφορίες περί ποικιλομορφίας Ανωνυμία δοτών Όμως… Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

78
Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

79
Ο Venter ομολόγησε ότι είχε παραβιάσει τη διαδικασία επιλογής
Το 2002 το συμβούλιο βιοηθικής της Celera τον απέλυσε Στη βιογραφία του αναφέρεται ότι ο Venter ήταν έτοιμος να φύγει από τη Celera και ότι απολύθηκε λόγω αντιπαλότητας με τον βασικό χρηματοδότη, Tony White. Ο Venter ισχυρίζεται ότι η βασική του επιδίωξη ήταν η «επιτάχυνση» της επιστήμης και ότι κατέφευγε σε ιδιωτικές χρηματοδοτήσεις μόνο όταν δεν μπορούσε να βρει χρήματα από δημόσιες πηγές να χρηματοδοτήσει τις ιδέες του. Ανασυνδυασμένο DNA Ακαδημαϊκές Εκδόσεις 2007

80
Τι να διαβάσετε Brown, Κεφάλαιο 4 Watson, Κεφάλαιο 10

Παρόμοιες παρουσιάσεις











. Θα πρέπει να υπάρχουν δύο διαφορετικά είδη.>")







 - Γ' ΦΑΣΗ Αποθετήρια και Επιστημονικά Ηλεκτρονικά Περιοδικά Ανοικτής Πρόσβασης»>")



