Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Χαρτογράφηση του γονιδιώματος

2
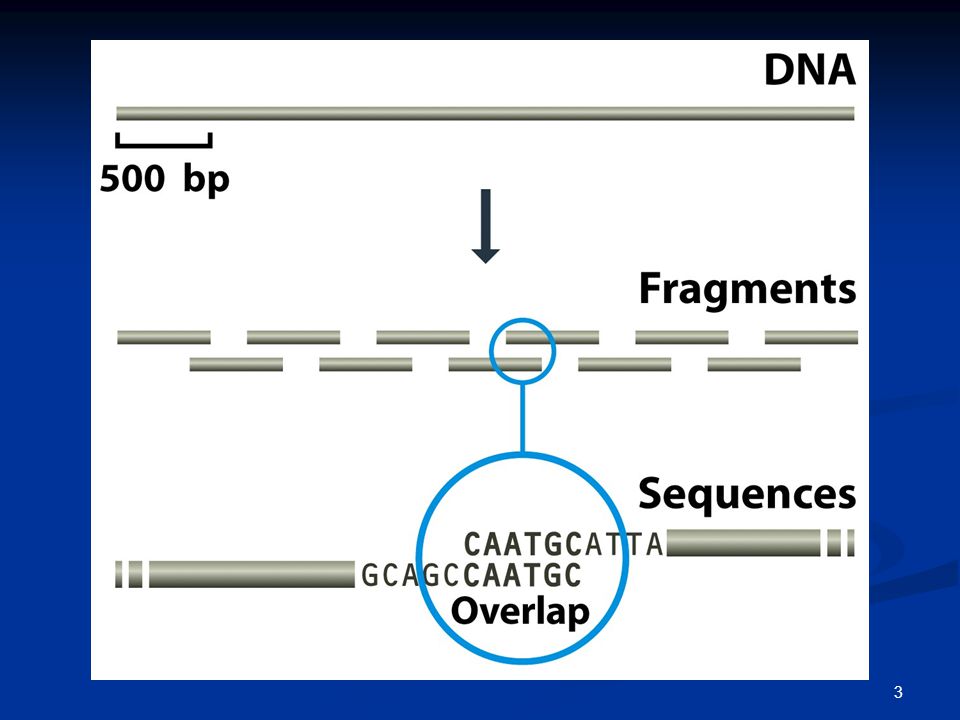
Το μέγιστο μήκος της αλληλουχίας του DNA που μπορεί να καθοριστεί με τις υπάρχουσες μεθόδους δεν ξεπερνάει ~1000 bp. Η αλληλουχία ενός μεγάλου μορίου DNA πρέπει να «συναρμολογηθεί».

4
Η ολοκληρωμένη αλληλουχία ενός τμήματος DNA προσδιορίζεται μέσω της συναρμολόγησης επικαλυπτόμενων αναγνώσεων αλληλουχίας.

5
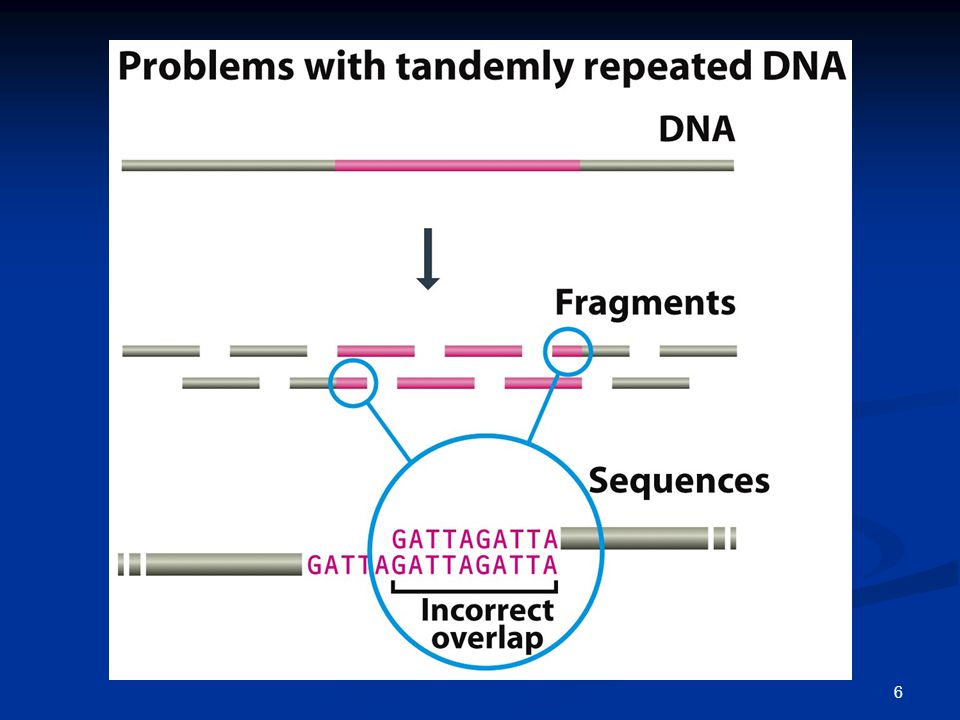
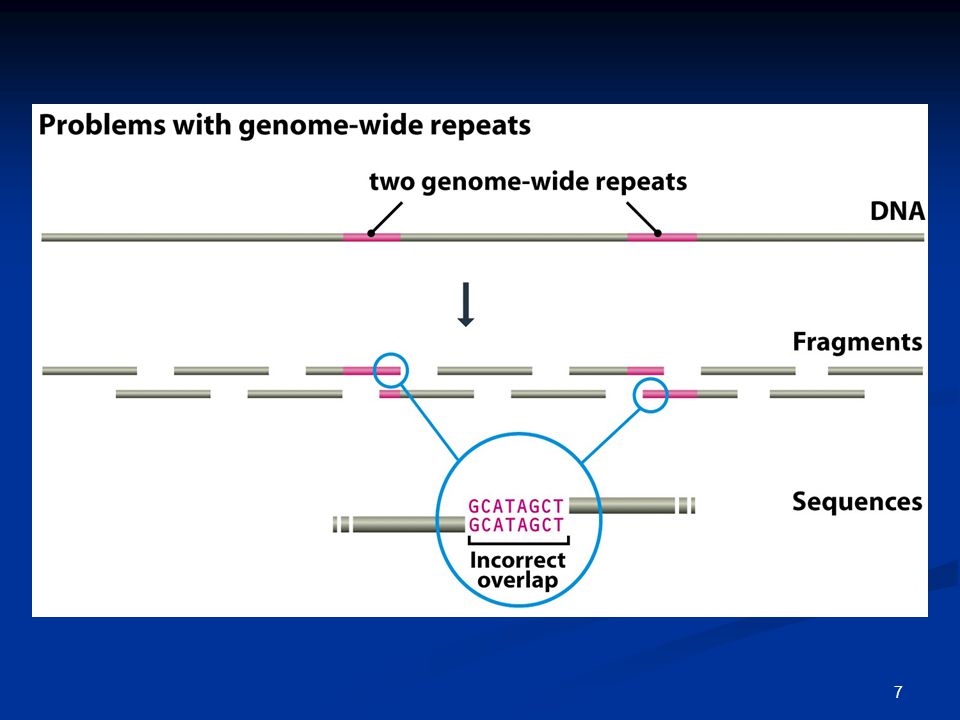
Προβλήματα κατά τη στοίχιση / συναρμολόγηση των αλληλουχιών
Επαναλαμβανόμενες αλληλουχίες; Διαχείριση τεράστιου αριθμού αναγνώσεων;

8
Στρατηγικές αλληλούχησης ολόκληρων γονιδιωμάτων
Μέθοδος των συνεχών κλώνων (clone contig method) Τυχαία προσπέλαση (whole-genome shotgun method)
 Τυχαία προσπέλαση (whole-genome shotgun method)")
9
Figure 3.3 Genomes 3 (© Garland Science 2007)
")
10
Γενετικοί και φυσικοί χάρτες
Γενετική χαρτογράφηση: χάρτες σύνδεσης γονιδίων σχετική θέση γονιδίων στα χρωμοσώματα Φυσική χαρτογράφηση: με τεχνικές μοριακής βιολογίας (in situ hybridization) καθορίζεται επακριβώς η φυσική θέση γονιδίων στα χρωμοσώματα
 καθορίζεται επακριβώς η φυσική θέση γονιδίων στα χρωμοσώματα.")
11
Γενετική χαρτογράφηση

12
«Γενετικοί δείκτες» : αλληλόμορφα με διακριτούς φαινότυπους
Ψηλοί και κοντοί μίσχοι στα μοσχομπίζελα του Μέντελ Χρώμα οφθαλμών στις φρουτόμυγες. Μέχρι το 1922, από τα 50 χαρτογραφημένα γονίδια, τα 9 έλεγχαν χρώμα ματιών Red, light red, vermilion, garnet, carnation, cinnabar, ruby, sepia, scarlet, pink, cardinal, claret, purple, brown

13
Τέτοιοι ορατοί χαρακτήρες ‘σκοράρονται’ με αναπόφευκτο υποκειμενισμό
Ανάγκη να βρεθούν πιο ‘αντικειμενικοί’ δείκτες: Βιοχημικοί δείκτες

14
DNA δείκτες για γενετική χαρτογράφηση
Τα γονίδια είναι χρήσιμοι δείκτες αλλά: Δεν καλύπτουν όλο το εύρος των χρωμοσωμάτων Δεν έχουν πολλά (διακριτά) αλληλόμορφα Υπόκεινται σε επιλογικές δράσεις DNA δείκτες RFLPs SSLPs SNPs
 αλληλόμορφα. Υπόκεινται σε επιλογικές δράσεις. DNA δείκτες. RFLPs. SSLPs. SNPs.")
15
Πολυμορφισμοί Μήκους κλασμάτων περιορισμού: Restriction Fragment Length Polymorphism (RFLP)
")
18
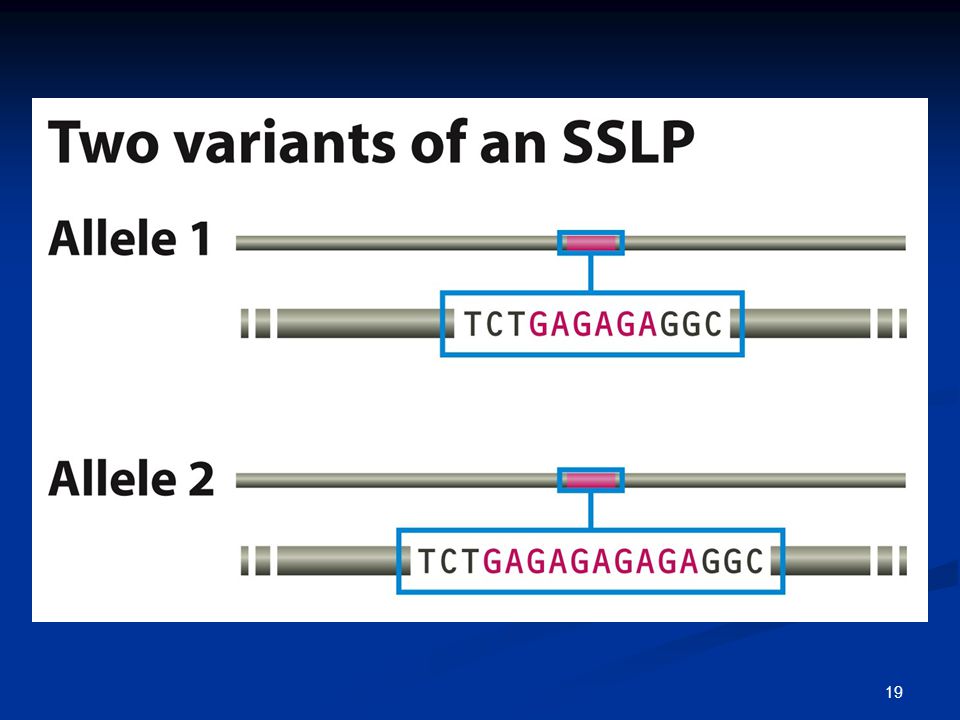
Πολυμορφισμοί μήκους απλής αλληλουχίας: Simple Sequence Length Polymorphisms (SSLP)
Minisatellites Microsatellites / Simple Tandem Repeats (STRs)
")
21
Πολυμορφισμοί μεμονωμένων νουκλεοτιδίων: Single Nucleotide Polymorphisms (SNPs)
Πάνω από 4 εκατομμύρια στο ανθρώπινο γονιδίωμα Δεν είναι όλα τα SNPs RFLPs, όμως πολλά είναι

22
Μέθοδοι τυποποίησης SNPs
Ειδικός υβριδισμός ολιγονουκλεοτιδίου

23
Ανίχνευση υβριδισμού μέσω αποχρωματισμού

24
Υβριδισμός με ολιγονουκλεοτίδιο με αταίριαστο νουκλεοτίδιο στο ένα άκρο

25
Δοκιμασία σύνδεσης ολιγονουκλεοτιδίων

26
Δοκιμασία ARMS

27
Στρατηγικές διαλογής SNPs που βασίζονται σε υβριδισμό ολιγονουκλεοτιδίων
Τεχνολογία DNA chips Τεχνική υβριδισμού διαλυμάτων

28
Ανάδειξη υβριδισμού φθορίζοντος ανιχνευτή σε μικροσυστοιχία
DNA chip

29
Ανάλυση γενετικής σύνδεσης

30
Ο πρώτος νόμος του Μέντελ: τα αλληλόμορφα διαχωρίζονται τυχαία
Στη μονοϋβριδική διασταύρωση μιας ψηλής μπιζελιάς (Τ) με μια κοντή μπιζελιά (t), ο Μέντελ πήρε 787 ψηλούς και 277 κοντούς απογόνους (αναλογία 2,84:1). Δηλαδή, ο κάθε απόγονος έχει την ίδια πιθανότητα να κληρονομήσει είτε το Τ είτε το t αλληλόμορφο (όπως φαίνεται στο αβάκιο).
 με μια κοντή μπιζελιά (t), ο Μέντελ πήρε 787 ψηλούς και 277 κοντούς απογόνους (αναλογία 2,84:1). Δηλαδή, ο κάθε απόγονος έχει την ίδια πιθανότητα να κληρονομήσει είτε το Τ είτε το t αλληλόμορφο (όπως φαίνεται στο αβάκιο).")
31
Ο δεύτερος νόμος του Μέντελ: τα ζεύγη των αλληλομόρφων διαχωρίζονται ανεξάρτητα
Στη διϋβριδική διασταύρωση μελετώνται δυο γονίδια. Το δεύτερο καθορίζει το σχήμα των μπιζελιών: στρογγυλά R ή ρυτιδωμένο r. Οι γονότυποι και οι φαινότυποι είναι εκείνοι που προβλέπονται από τον πρώτο και δεύτερο νόμο του Μέντελ, ο οποίο ορίζει ότι τα ζεύγη των αλληλομόρφων διαχωρίζονται ανεξάρτητα.

32
Αν όμως τα γονίδια βρίσκονται στο ίδιο χρωμόσωμα;
Τα Α και Β θα έπρεπε να κληρονομούνται μαζί. Κατά συνέπεια, ο δεύτερος νόμος του Μέντελ δεν θα έπρεπε να ισχύει για την κληρονομική μεταβίβαση των Α και Β. Το C βρίσκεται σε διαφορετικό χρωμόσωμα, οπότε ο νόμος του Μέντελ θα πρέπει να ισχύει για τη μεταβίβαση των Α και C ή Β και C. O Μέντελ δεν ανακάλυψε τη σύνδεση γιατί καθένα από τα 7 γονίδια που μελέτησε ήταν σε διαφορετικά χρωμοσώματα.

33
Μερική σύνδεση

34
Τα συμβάντα σε ένα ζεύγος ομόλογων χρωμοσωμάτων κατά τη μείωση στα αναπαραγωγικά κύτταρα

35
Οι συνέπειες του επιχιασμού στα συνδεδεμένα γονίδια:
Οι συνέπειες του επιχιασμού στα συνδεδεμένα γονίδια: Αριστερά, χωρίς ανασυνδυασμό (γονικοί γαμέτες) Δεξιά με ανασυνδυασμό (δύο γονικοί και δύο ανασυνδυασμένοι γαμέτες) Αν εξετάζαμε το αποτέλεσμα της μείωσης σε 100 πανομοιότυπα κύτταρα και παίρναμε: 200AB 200 ab Πλήρης σύνδεση Αν όμως συνέβαινε επιχιασμός σε 40 από τις 100 μειώσεις τότε θα παίρναμε: 160 AB 160 ab 40 Ab 40 aB Μερική σύνδεση
 Δεξιά με ανασυνδυασμό (δύο γονικοί και δύο ανασυνδυασμένοι γαμέτες) Αν εξετάζαμε το αποτέλεσμα της μείωσης σε 100 πανομοιότυπα κύτταρα και παίρναμε: 200AB. 200 ab. Πλήρης σύνδεση. Αν όμως συνέβαινε επιχιασμός σε 40 από τις 100 μειώσεις τότε θα παίρναμε: 160 AB. 160 ab. 40 Ab. 40 aB. Μερική σύνδεση.")
36
Από τη μερική σύνδεση στη γενετική χαρτογράφηση: Από τη συχνότητα ανασυνδυασμού συνάγουμε τις σχετικές θέσεις των γονιδίων πάνω σε ένα χρωμόσωμα

37
Όταν προγραμματισμένα πειράματα διασταυρώσεων είναι «ανήθικα»

38
Τρεις τρόποι μεταφοράς DNA μεταξύ βακτηρίων

39
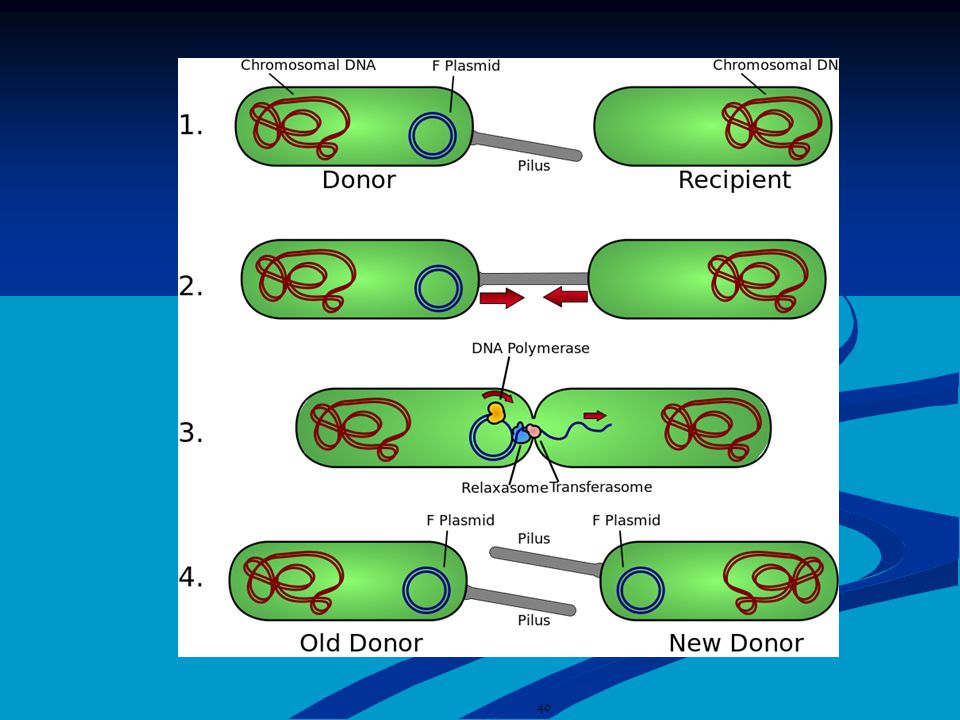
Βακτηριακή σύζευξη

42
Βακτηριοφάγοι

43
Ο κύκλος ζωής του φάγου

45
Γενικευμένη μεταγωγή

46
Ειδικευμένη μεταγωγή

47
Οι αρχές με τις οποίες επιτελείται μια γενετική διασταύρωση με βακτηριοφάγους
(α) Βακτήρια του στελέχους B της E. coli μολύνονται ταυτόχρονα με τους δύο γονικούς βακτηριοφάγους h+r και hr+. (β) Αντιγραφή και των δύο γονικών χρωμοσωμάτων. (γ) Πραγματοποιείται ζευγάρωμα κάποιων χρωμοσωμάτων από κάθε γονικό τύπο και συμβαίνει διασκελισμός μεταξύ των δύο γενετικών τόπων, με αποτέλεσμα τη δημιουργία ανασυνδυασμένων φάγων h+r+ και hr. (δ) Όταν τα βακτήρια λύονται, οι απόγονοι φάγοι συναρμολογούνται και απελευθερώνονται στο θρεπτικό μέσο. Μεταξύ των απογόνων εντοπίζονται τόσο οι γονικοί όσο και οι ανασυνδυασμένοι φάγοι.
 Βακτήρια του στελέχους B της E. coli μολύνονται ταυτόχρονα με τους δύο γονικούς βακτηριοφάγους h+r και hr+. (β) Αντιγραφή και των δύο γονικών χρωμοσωμάτων. (γ) Πραγματοποιείται ζευγάρωμα κάποιων χρωμοσωμάτων από κάθε γονικό τύπο και συμβαίνει διασκελισμός μεταξύ των δύο γενετικών τόπων, με αποτέλεσμα τη δημιουργία ανασυνδυασμένων φάγων h+r+ και hr. (δ) Όταν τα βακτήρια λύονται, οι απόγονοι φάγοι συναρμολογούνται και απελευθερώνονται στο θρεπτικό μέσο. Μεταξύ των απογόνων εντοπίζονται τόσο οι γονικοί όσο και οι ανασυνδυασμένοι φάγοι.")
48
Προκαταρκτικός γενετικός χάρτης λεπτής δομής της περιοχής rII του φάγου T4. Ο χάρτης προέκυψε από τον Benzer μετά από διασταυρώσεις μιας αρχικής σειράς 60 διαφορετικών μεταλλαγμένων στελεχών στην περιοχή rII. 48

49
Τμηματική υποδιαίρεση της περιοχής rII του φάγου T4 με τη χρησιμο- ποίηση των ελλειμμάτων.
Το Επίπεδο I δείχνει το συνολικό γενε- τικό χάρτη του T4. Στο Επίπεδο II, επτά ελλείμματα ορίζουν επτά τμήματα της περιοχής rII. Στο Επίπεδο III, τρία ελ- λείμματα ορίζουν τέσσερα υποτμήματα των τμημάτων A5. Στο Επίπεδο IV, τρία ελλείμματα ορίζουν τέσσερα υποτμή- ματα του τμήματος A5γ. Στο Επίπεδο V παρουσιάζεται η διάταξη και η σχετική απόσταση των θέσεων των μεταλλαγών rII στο υποτμήμα A5γ2α2, όπως προέκυψε από διασταυρώσεις κατά ζεύγη των επτά σημειακών μεταλ- λαγμάτων. Το Επίπεδο VI είναι ένα μοντέλο της διπλής έλικας του DNA στο οποίο φαίνεται η κλίμακα, κατά προσέγγιση, του χάρτη στο Επίπεδο V. 49

50
Τη δεκαετία του 1950, o Benzer χαρτογράφησε ανεξάρτητες μεταλλάξεις με διαλληλικές διασταυρώσεις ή και με τη χρησιμοποίηση ελλειμμάτων και κατέληξε: Το γονίδιο αποτελείται από περιοχές οι οποίες είναι αδιαίρετες από διασκελισμό (recons) Το γονίδιο αποτελείται από περιοχές οι οποίες μπορούν να μεταλλαχθούν (mutons) Οι μεταλλάξεις δεν συμβαίνουν με την ίδια συχνότητα κατά μήκος ενός γονιδίου Ο γονιδιακός τόπος rII του φάγου Τ4 αποτελείται από δύο λειτουργικές περιοχές Α και Β (cistrons)
 Το γονίδιο αποτελείται από περιοχές οι οποίες μπορούν να μεταλλαχθούν (mutons) Οι μεταλλάξεις δεν συμβαίνουν με την ίδια συχνότητα κατά μήκος ενός γονιδίου. Ο γονιδιακός τόπος rII του φάγου Τ4 αποτελείται από δύο λειτουργικές περιοχές Α και Β (cistrons)")
51
Χάρτης λεπτής δομής της περιοχής rII που προέκυψε από τα πειράματα του Benzer.
Ο αριθμός των ανεξάρτητα απομονωμένων μεταλλαγών που χαρτογραφήθηκαν σε μια δεδομένη θέση υποδεικνύεται από τον αριθμό των μικρών τετραγώνων που υπάρχουν στη θέση αυτή. Τα θερμά σημεία αντιπροσωπεύονται από ένα μεγάλο αριθμό τέτοιων τετραγώνων. 51

52
Φυσική χαρτογράφηση

53
Η γενετική χαρτογράφηση δεν επαρκεί
Η ανάλυση ενός γενετικού χάρτη εξαρτάται από τον αριθμό των επιχιασμών που καταγράφονται Δεν είναι πρόβλημα για μικροοργανισμούς (πχ E. coli, S. cerevisiae) Είναι πρόβλημα για άνθρωπο και άλλους ευκαρυώτες Οι γενετικοί χάρτες έχουν περιορισμένη ακρίβεια Hotspots, μη ομοιόμορφη κατανομή επιχιασμών
 Είναι πρόβλημα για άνθρωπο και άλλους ευκαρυώτες. Οι γενετικοί χάρτες έχουν περιορισμένη ακρίβεια. Hotspots, μη ομοιόμορφη κατανομή επιχιασμών.")
54
Σύγκριση μεταξύ γενετικού και φυσικού χάρτη του χρωμοσώματος ΙΙΙ του Saccharomyces cerevisiae

55
Χαρτογράφηση περιορισμού (restriction mapping)
Φθορίζων υβριδισμός in situ (fluorescent in situ hybridization, FISH) Χαρτογράφηση με σημασμένες αλληλουχίες (sequence tagged site, STS, mapping)
 Χαρτογράφηση με σημασμένες αλληλουχίες (sequence tagged site, STS, mapping)")
56
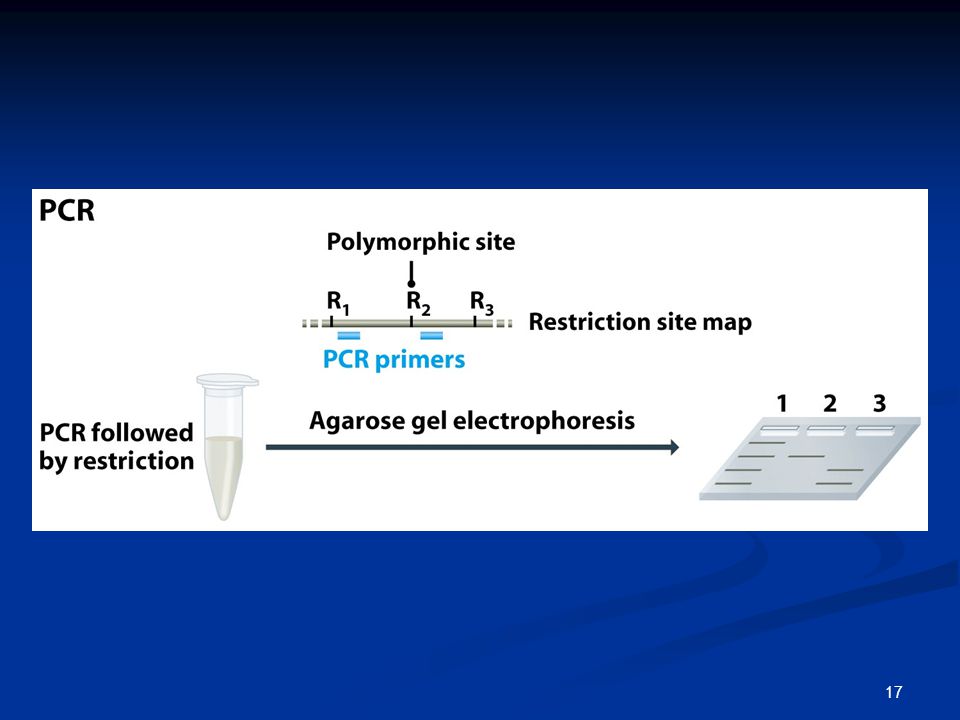
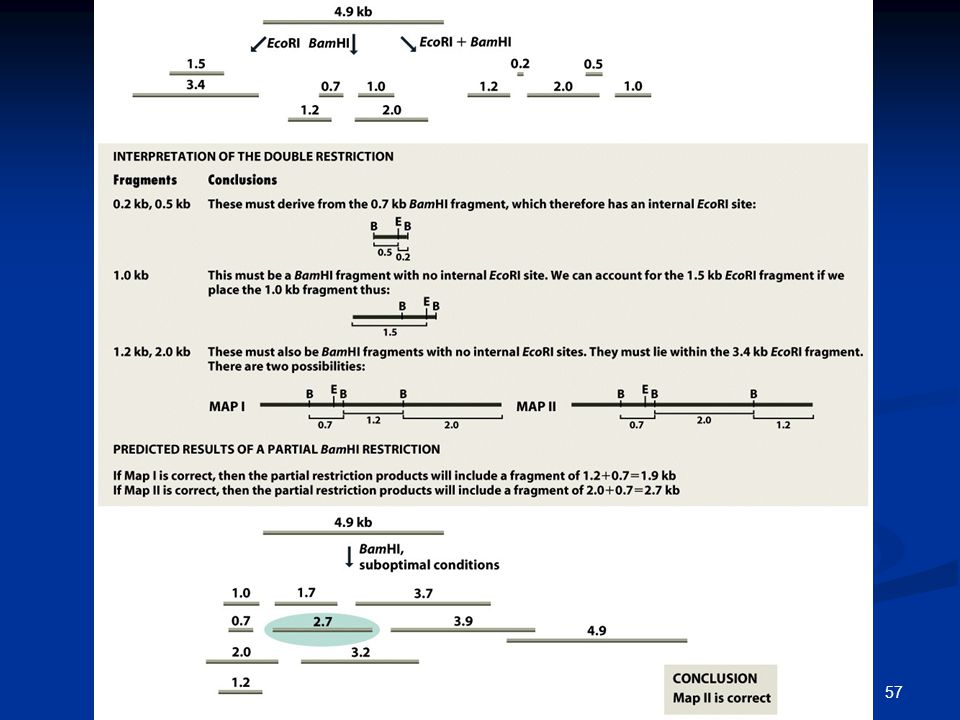
1. Χαρτογράφηση περιορισμού
Πολύ λίγες από τις θέσεις περιορισμού είναι πολυμορφικές δεν χαρτογραφούνται πολλές θέσεις Χαρτογράφηση μη πολυμορφικών θέσεων;

58
Ανάλυση μερικού περιορισμού με σήμανση στο άκρο του DNA

59
H χαρτογράφηση περιορισμού είναι εφικτή όταν:
Υπάρχουν σχετικά λίγες θέσεις κοπής Το προς ανάλυση DNA είναι σχετικά μικρού μεγέθους Λύσεις: «σπάνιοι κόφτες» Ένζυμα που αναγνωρίζουν 7 ή 8 νουκλεοτίδια Ένζυμα που αναγνωρίζουν σπάνια μοτίβα (GC rich)
")
60
Πώς διαχωρίζονται μεγάλα μόρια DNA;
Το πρόβλημα: Όλα τα δίκλωνα DNA μόρια που είναι μεγαλύτερα από ~40kb μετακινούνται σε ένα πήκτωμα αγαρόζης όχι ανάλογα του μήκους τους αλλά ανάλογα του ηλεκτρικού πεδίου Άρα, τα μεγάλα DNA μόρια είναι αδύνατον να διαχωριστούν σε πηκτώματα αγαρόζης

61
Η λύση: Ηλεκτροφόρηση παλμικού πεδίου - Pulsed Field Gel Electrophoresis Διαχωρισμός DNA μορίων ~5Mb Pulse time: από μερικά sec σε ώρες: όσο μεγαλύτερα τα DNA μόρια τόσο μεγαλύτερος "παλμός" χρειάζεται για το διαχωρισμό τους

62
Οπτική χαρτογράφηση

63
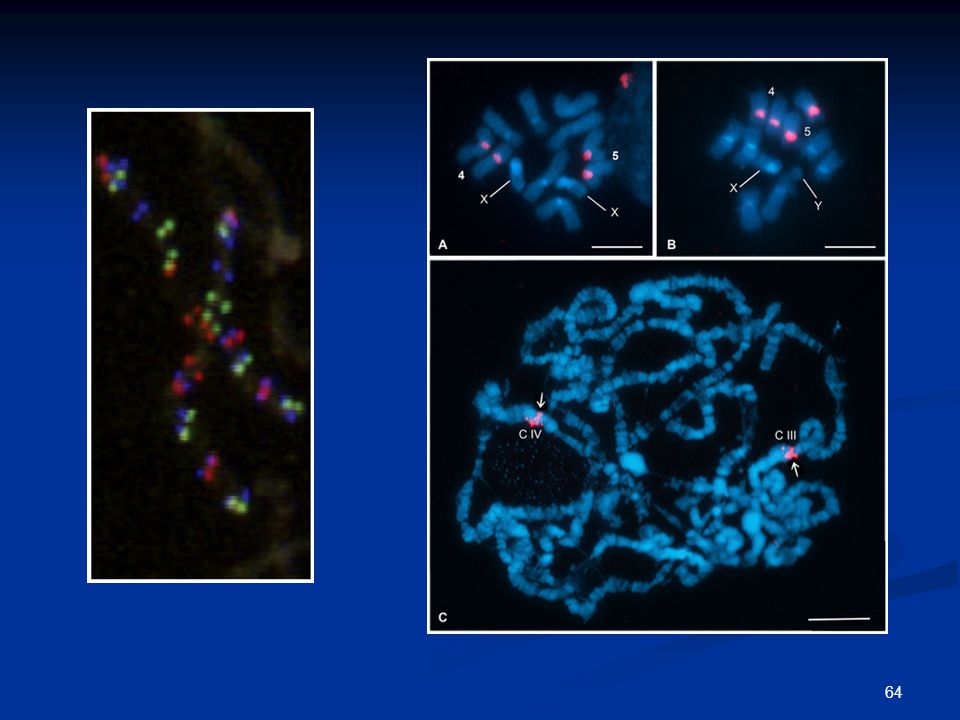
Φθορίζων in situ υβριδισμός (FISH)
")
65
Sequence-Tagged Sites
Ετικέτες μικρού μήκους χρησιμεύουν για τον προσδιορισμό του προσανατολισμού και της διαδοχικής σειράς γονιδιωματικών κλώνων Ενώ ένα STS αντιπροσωπεύει ένα φυσικό τμήμα DNA μοναδικό στο γονιδίωμα, δεν χρειάζεται ένας κλώνος αλλά μπορεί να παραχθεί μέσω μιας PCR Η PCR είναι ταχύτερη και πιο εύχρηστη από το στύπωμα κατά Southern

66
Χαρτογράφηση με βάση την ανίχνευση STS σε κλώνους BAC.

67
Κοινές πηγές STSs Expressed Sequence Tags (ESTs) Αλληλουχίες SSLPs
Τυχαίες αλληλουχίες του γονιδιώματος

68
Χαρτογράφηση με υβρίδια ακτινοβολημένων κυττάρων
1990: Οι Cox και Myers βελτίωσαν μια παλιά (Goss and Harris, 1975) μέθοδο δημιουργίας υβριδίων κυττάρων τρωκτικών και ανθρώπου
 μέθοδο δημιουργίας υβριδίων κυττάρων τρωκτικών και ανθρώπου.")
69
Παραγωγή σειρών υβριδίων ακτινοβολημένων κυττάρων με σκοπό τη χαρτογράφηση του γονιδιώματος.
Οι Goss & Harris δημιούργησαν υβριδικά κύτταρα με πολύ μικρά τμήματα ανθρώπινων χρωμοσωμάτων (ισχυρές δόσεις ακτίνων Χ). Στη συνέχεια προκάλεσαν σύντηξη με κύτταρα τρωκτικών και απομόνωσαν κυτταρικές σειρές κάθε μία των οποίων είχε ένα μόνο τμήμα ανθρώπινου γονιδιώματος. Τέλος, έκαναν μελέτες σύνδεσης στα υβριδικά κύτταρα, προσδιορίζοντας τη συχνότητα με την οποία δύο γονίδια (ή δείκτες) συγκληρονομούνταν (βρίσκονταν μαζί σε ένα σετ υβριδίων).
. Στη συνέχεια προκάλεσαν σύντηξη με κύτταρα τρωκτικών και απομόνωσαν κυτταρικές σειρές κάθε μία των οποίων είχε ένα μόνο τμήμα ανθρώπινου γονιδιώματος. Τέλος, έκαναν μελέτες σύνδεσης στα υβριδικά κύτταρα, προσδιορίζοντας τη συχνότητα με την οποία δύο γονίδια (ή δείκτες) συγκληρονομούνταν (βρίσκονταν μαζί σε ένα σετ υβριδίων).")
70
Οι Cox και Meyers δημιούργησαν ένα σετ 90 ακτινοβολημένων υβριδίων
Οι σειρές αυτές γονοτυπήθηκαν για την παρουσία ή απουσία χιλιάδων STSs Τα πρότυπα παρουσίας/απουσίας των STSs συγκρίθηκαν ώστε να προβλεφθεί η πιθανή σχετική διάταξη των STSs καθώς και η απόσταση μεταξύ τους

71
Εφαρμογή της χαρτογράφησης μέσω ανάλυσης υβριδίων ακτινοβολημένων κυττάρων (RH, Radiation Hybrid) για τον προσδιορισμό της διάταξης ετικετών θέσεων αλληλουχίας (STS, Sequence-Tagged Sites). Αν η εμφάνιση δυο STSs (πχ 1 και 19) σε πολλές κυτταρικές σειρές είναι πιο συχνή από ό,τι θα αναμενόταν τυχαία, δείχνει ότι αυτοί οι δύο δείκτες έχουν μικρή απόσταση μεταξύ τους.
 σε πολλές κυτταρικές σειρές είναι πιο συχνή από ό,τι θα αναμενόταν τυχαία, δείχνει ότι αυτοί οι δύο δείκτες έχουν μικρή απόσταση μεταξύ τους.")
72
Χρησιμοποιήθηκαν δύο τέτοιες προσεγγίσεις:
Παρότι η διακριτική ικανότητα ενός τέτοιου χάρτη είναι χαμηλή, αποτελεί έναν ανεξάρτητο τρόπο προσδιορισμού της διάταξης τμημάτων DNA Χρησιμοποιήθηκαν δύο τέτοιες προσεγγίσεις: Ο ένας χάρτης είχε διακριτική ικανότητα ~1,5 εκατ bp Ο άλλος ~400,000 bp

73
Brown, Κεφάλαιο 3 Watson, Κεφάλαιο 10

Παρόμοιες παρουσιάσεις