Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Τεχνικές Γονιδιωματικής Κλωνοποίησης
Βακτηριοφάγο Μ13 Ο Μ13 είναι ένας νηματοειδής φάγος που περιέχει μονόκλωνο DNA και αντιγράφεται χωρίς να θανατώνει τον ξενιστή του. Ώριμα σωματίδια του Μ13 απελευθερώνονται από τα κύτταρα του ξενιστή με μια διαδικασία εκβλάστησης και έτσι είναι εφικτή η απόκτηση μολυσμένων καλλιεργειών, οι οποίες αποτελούν διαρκή πηγή φαγικού DNA. Σημαντική ιδιότητα του Μ13 αποτελεί το μονόκλωνο DNA του. Για να χρησιμοποιηθεί ο Μ13 στην κλωνοποίηση, απαιτείται η ύπαρξη δίκλωνης μορφής του, επειδή τα περιοριστικά ένζυμα δρουν μόνο εναντίον δίκλωνου DNA. Δίκλωνο DNA του Μ13 απομονώνεται από μολυσμένα κύτταρα, διότι ο Μ13 αντιγράφεται στον ξενιστή ως δίκλωνο αντιγραφικό ενδιάμεσο.

3
Χάρτης του φορέα αυτού παρουσιάζεται στην κατωτέρω εικόνα .
Υπάρχει μια μικρή περιοχή που ονομάζεται διαγονιδιακή αλληλουχία και που μπορεί να χρησιμοποιηθεί ως θέση κλωνοποίησης. Εκεί μπορεί να κλωνοποιηθεί DNA ξένο προς τον ιό, με μήκος έως 5 ζεύγη κιλοβάσεων, χωρίς επίδραση στη βιωσιμότητα του φάγου. Όσο μεγαλώνει το γονιδίωμα, τόσο αυξάνεται το μήκος του ιοσώματος. Ο Μ13mp18 είναι παράγωγο του Μ13 στο οποίο η διαγονιδιακή περιοχή έχει τροποποιηθεί κατάλληλα ώστε να διευκολύνεται η κλωνοποίηση. Χάρτης του φορέα αυτού παρουσιάζεται στην κατωτέρω εικόνα . Η ένθεση ενός λειτουργικού τμήματος του lacZ, του γονιδίου του Escherichia coli κωδικοποιεί το ένζυμο β-γαλακτοζιδάση. Κατά συνέπεια, κύτταρα μολυσμένα με τον Μ13mp18 ανιχνεύονται εύκολα από το χρώμα των αποικιών τους σε τρυβλία-δείκτες. Το γονίδιο lacZ έχει τροποποιηθεί κατάλληλα ώστε να περιέχει τμήμα DNA 54 ζευγών βάσεων που, ονομάζεται θέση πολλαπλής κλωνοποίησης (polylinker). Η θέση πολλαπλής κλωνοποίησης περιέχει αρκετές θέσεις περιοριστικών ενζύμων που υπάρχουν μία μόνο φορά στον M13 και κατά συνέπεια μπορούν να χρησιμοποιηθούν στην κλωνοποίηση. Η θέση πολλαπλής κλωνοποίησης εισάγεται στην αρχή της περιοχής κωδικοποίησης του γονιδίου lacZ.
. Η θέση πολλαπλής κλωνοποίησης περιέχει αρκετές θέσεις περιοριστικών ενζύμων που υπάρχουν μία μόνο φορά στον M13 και κατά συνέπεια μπορούν να χρησιμοποιηθούν στην κλωνοποίηση. Η θέση πολλαπλής κλωνοποίησης εισάγεται στην αρχή της περιοχής κωδικοποίησης του γονιδίου lacZ.")
4
Η μικρή αυτή ένθεση γίνεται έτσι ώστε να βρίσκεται εντός πλαισίου ανάγνωσης, και τα 18 επιπλέον αμινοξέα δεν επηρεάζουν τη δραστικότητα του ενζύμου που κωδικοποιείται από το γονίδιο. Η κλωνοποίηση και άλλου τμήματος DNA στη θέση πολλαπλής κλωνοποίησης αδρανοποιεί το γονίδιο. Φάγοι στους οποίους εισήχθησαν πρόσθετα τμήματα DNA προκαλούν την εμφάνιση άχρωμων πλακών.

5
α. Ο φορέας διαθέτει τον υποκινητή lac και ένα γονίδιο το lacZ’ το οποίο κωδικοποιεί ένα τμήμα της β-γαλακτοζιδάσης.Στην αρχή του γονιδίου υπάρχει μια θέση πολλαπλής κλωνοποίησης περιοριστικών ενζύμων η οποία δεν τροποποιεί το πλαίσιο ανάγνωσης.Τα τμήματα DNA που κλωνοποιούνται στη θέση αυτή διακόπτουν την αλληλουχία του γονιδίου lacZ’ και σταματούν την δράση της β-γαλακτοζιδάσης . β & γ .Τρυβλίο με πλάκες που σχηματίσθηκαν από τον M13mp18 σε θρεπτικό υπόστρωμα που περιείχε την Χ-gal. Η β-γαλακτοζιδάση υδρολύει την Χ-gal απελευθερώνοντας μια αδιάλυτη μπλε χρωστική.Οι πλάκες στο τρυβλίο με μπλε χρώμα δηλώνουν την παρουσία φορέα χωρίς κλωνοποιημένο DNA .Οι άχρωμες πλάκες δηλώνουν ότι ξένο DNA έχει εισαχθεί και έχει διακόψει την αλληλουχία του γονιδίου lacZ’ .

6
Για να κλωνοποιηθεί DNA σε φορείς του Μ13, απομονώνεται το δίκλωνο αντιγραφικό ενδιάμεσο από τον μολυσμένο φορέα, στο οποίο προστίθεται ένα περιοριστικό ένζυμο. Στο ξένο DNA, επίσης δίκλωνο, προστίθεται το ίδιο ένζυμο. Με τη συνένωση των δύο μορίων προκύπτουν δίκλωνα μόρια του Μ13 που περιέχουν το ξένο DNA. Όταν τα μόρια αυτά εισάγονται με μετασχηματισμό στο κύτταρο, αντιγράφονται και ύστερα από κάποιο χρονικό διάστημα παράγουν ώριμα σωμάτια βακτηριοφάγου που περιέχουν μονόκλωνα μόρια DNA. Μόνο μια αλυσίδα DNA «συσκευάζεται» μέσα σε έναν ώριμο φάγο. Το ποια από τις δύο αλυσίδες περιέχει ο φάγος εξαρτάται από τον προσανατολισμό με τον οποίο έγινε η ένθεση του DNA. Επειδή είναι δυνατόν να εισαχθεί ξένο DNA (σε χωριστά φαγικά σωμάτια) με οιονδήποτε προσανατολισμό, κλωνοποιούνται και οι δύο αλυσίδες. Το μονόκλωνο DNA του Μ13 που περιέχει ξένο DNA μπορεί κατόπιν να χρησιμοποιηθεί για προσδιορισμό της αλληλουχίας του. Δεδομένου ότι είναι γνωστή η βασική αλληλουχία όπου έγινε η σύνθεση του ξένου DNA (βάσει της εξειδίκευσης του περιοριστικού ενζύμου που χρησιμοποιήθηκε), μπορούμε να χρησιμοποιήσουμε ως εκκινητή ένα συνθετικό ολιγονουκλεοτίδιο συμπληρωματικό της περιοχής
 με οιονδήποτε προσανατολισμό, κλωνοποιούνται και οι δύο αλυσίδες. Το μονόκλωνο DNA του Μ13 που περιέχει ξένο DNA μπορεί κατόπιν να χρησιμοποιηθεί για προσδιορισμό της αλληλουχίας του. Δεδομένου ότι είναι γνωστή η βασική αλληλουχία όπου έγινε η σύνθεση του ξένου DNA (βάσει της εξειδίκευσης του περιοριστικού ενζύμου που χρησιμοποιήθηκε), μπορούμε να χρησιμοποιήσουμε ως εκκινητή ένα συνθετικό ολιγονουκλεοτίδιο συμπληρωματικό της περιοχής.")
7
αυτής ( Universal Sequencing Primer o οποίος υβριδίζεται σε μια θέση εκτός της θέσης πολλαπλής κλωνοποίησης περιοριστικών ενζύμων) και κατά συνέπεια να προσδιορίσουμε την αλληλουχία ολόκληρου του DNA πέραν του σημείου αυτού.
 και κατά συνέπεια να προσδιορίσουμε την αλληλουχία ολόκληρου του DNA πέραν του σημείου αυτού.")
8
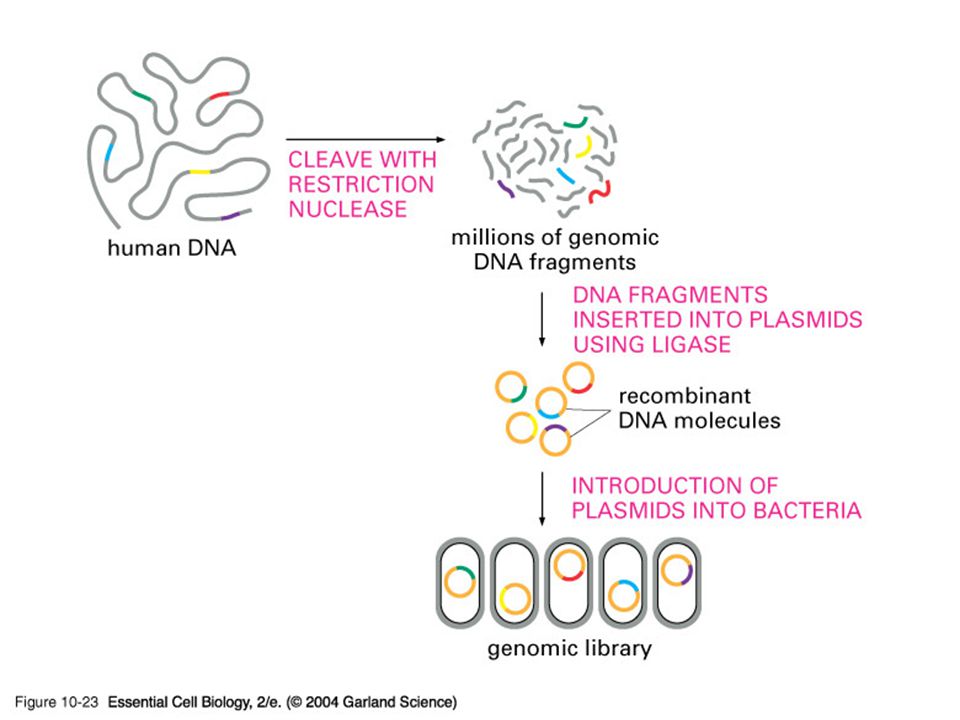
Τεχνητά χρωμοσώματα Μερικές φορές χρησιμοποιούνται φορείς του λ, οι οποίου δέχονται 20 κιλοβάσεις ή περισσότερο. Για την κατασκευή βιβλιοθηκών από DNA ευκαρυωτικών μικροοργανισμών ή ανώτερων ευκαρυωτών, όπως ο άνθρωπος, είναι χρήσιμο να υπάρχουν διαθέσιμοι φορείς που μπορούν να δεχτούν πολύ μεγάλα τμήματα DNA. Τέτοιοι φορείς έχουν αναπτυχθεί και ονομάζονται τεχνητά χρωμοσώματα. Το πλασμίδιο F της Escherichia coli αντιγράφεται στην E. coli με μεγάλη σταθερότητα, ενώ είναι γνωστές φυσικές παραλλαγές του, που ονομάζονται πλασμίδια F΄, τα οποία έχουν την ικανότητα να φέρουν μεγάλα τμήματα χρωμοσωματικού DNA. Το πλασμίδιο F έχει χρησιμοποιηθεί για να κατασκευασθούν φορείς κλωνοποίησης που ονομάζονται τεχνητά βακτηριακά χρωμοσώματα (bacterial artificial chromosomes ή BAC). Το BAC που φαίνεται στην κατωτέρω εικόνα έχει μέγεθος 6,7 kb, αντί των 99,2 kb του F.
. Το BAC που φαίνεται στην κατωτέρω εικόνα έχει μέγεθος 6,7 kb, αντί των 99,2 kb του F.")
9
Περιέχει λίγα μόνο γονίδια του F, στα οποία συμπεριλαμβάνονται τα oriS και repE, που είναι απαραίτητα για την αντιγραφή, και τα sopA και sopB, τα οποία διατηρούν τον αριθμό αντιγραφών του πλασμιδίου στο κύτταρο πολύ χαμηλό. Στο πλασμίδιο έχει εισαχθεί και το γονίδιο cat, το οποίο προσδίδει ανθεκτικότητα στη χλωραμφαινικόλη και περιέχει μια περιοχή κλωνοποίησης που περιλαμβάνει αρκετές θέσεις περιοριστικών ενζύμων για την κλωνοποίηση DNA. Σε φορείς BAC μπορεί να εισαχθεί και να διατηρηθεί σταθερά ξένο DNA μεγαλύτερο των 300 kb. Ο ξενιστής ενός BAC που περιέχει κλωνοποιημένο DNA είναι κατά κανόνα κάποιο μεταλλαγμένο στέλεχος της E. Coli, το οποίο δεν διαθέτει τα φυσιολογικά συστήματα περιορισμού και τροποποίησης DNA.

10
Ο όρος τεχνητό χρωμόσωμα προήλθε όχι από βακτήρια, αλλά από την ανάπτυξη τεχνητών χρωμοσωμάτων ζυμομύκητα (yeast artificial chromosome ή YAC). Προκειμένου να λειτουργήσουν όπως τα κανονικά ευκαρυωτικά χρωμοσώματα, τα YAC πρέπει να διαθέτουν μια αρχή αντιγραφής του DNA, τελομερή στα άκρα του χρωμοσώματος και ένα κεντρομερές (το τμήμα του χρωμοσώματος που είναι απαραίτητο για τον διαχωρισμό κατά τη μίτωση). Πρέπει επίσης να διαθέτουν μια θέση κλωνοποίησης και ένα γονίδιο επιλογής μετά τον μετασχηματισμό τους στον ξενιστή. Οι ίδιοι οι φορείς YAC έχουν μέγεθος μόνο 10 ζεύγη kb, μπορούν όμως να δεχτούν την ένθεση κλωνοποιημένου DNA μήκους ζεύγη Kb. Αν και τα YAC μπορούν να συγκρατήσουν μεγαλύτερες ενθέσεις DNA απ’ ό,τι τα BAC, υπάρχει μεγαλύτερο πρόβλημα ανασυνδυασμού και επαναδιάταξης του κλωνοποιημένου DNA στον ζυμομύκητα σε σχέση με την E. Coli.
. Πρέπει επίσης να διαθέτουν μια θέση κλωνοποίησης και ένα γονίδιο επιλογής μετά τον μετασχηματισμό τους στον ξενιστή. Οι ίδιοι οι φορείς YAC έχουν μέγεθος μόνο 10 ζεύγη kb, μπορούν όμως να δεχτούν την ένθεση κλωνοποιημένου DNA μήκους ζεύγη Kb. Αν και τα YAC μπορούν να συγκρατήσουν μεγαλύτερες ενθέσεις DNA απ’ ό,τι τα BAC, υπάρχει μεγαλύτερο πρόβλημα ανασυνδυασμού και επαναδιάταξης του κλωνοποιημένου DNA στον ζυμομύκητα σε σχέση με την E. Coli.")
12
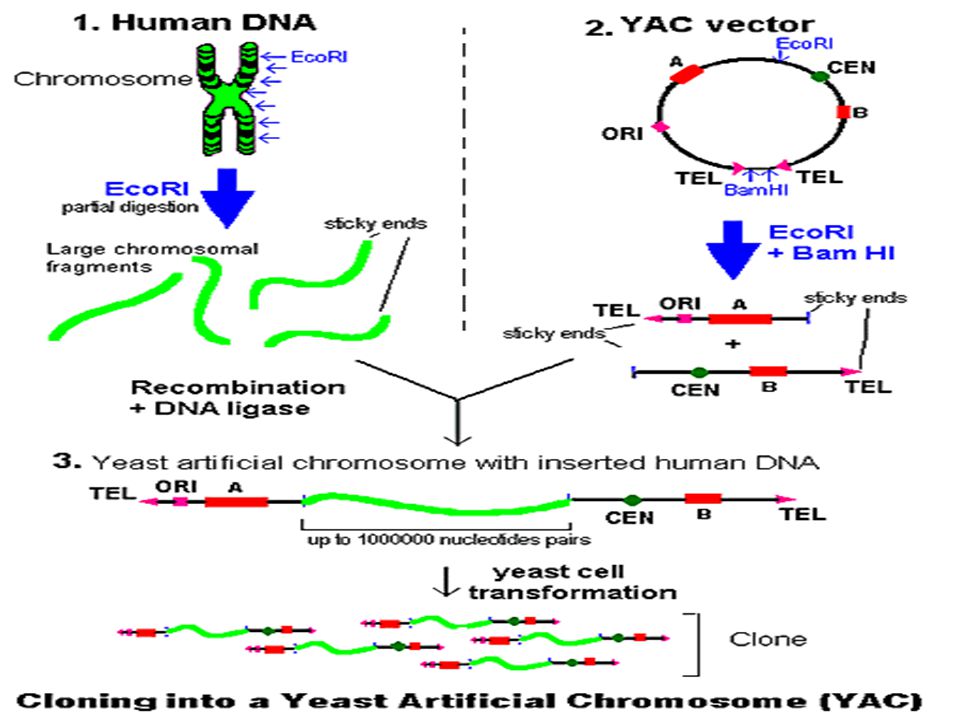
Cloning human genomic DNA into a YAC:
Genomic DNA is partially digested by the restriction enzyme EcoRI. Very large DNA fragments are obtained. The YAC is digested by the two restriction enzymes EcoRI and BamHI. Those two elements recombine at the EcoRI sites and are covalently linked by the DNA ligase. A recombinant YAC vector, a yeast artificial chromosome with genomic DNA inserted, is produced. This vector can be used to infect yeast cells and generated an unlimited number of copies. TEL: τελομερή τα οποία προστατεύουν τα άκρα του χρωμοσώματος από την δράση των νουκλεασών. CEN: το κεντρομερές ORI: αλληλουχία για την αρχή αντιγραφής του DNA A and B: markers για την επιλογή και την απομόνωση των ζυμομυκήτων. Αν ο ξενιστής είναι ουρακίλη - και εισάγεται με μετασχηματισμό σε αυτόν ο κλώνος ο οποίος είναι ουρακίλη + τότε θα αναπτυχθούν μόνο τα κύτταρα τα οποία περιέχουν το YAC και κατά συνέπεια είναι ουρακίλη + . Θέσεις αναγνώρισης για τα ένζυμα περιορισμού EcoRI και BamHI.

13
«Τυφλή» κλωνοποίηση βακτηριακού DNA

15
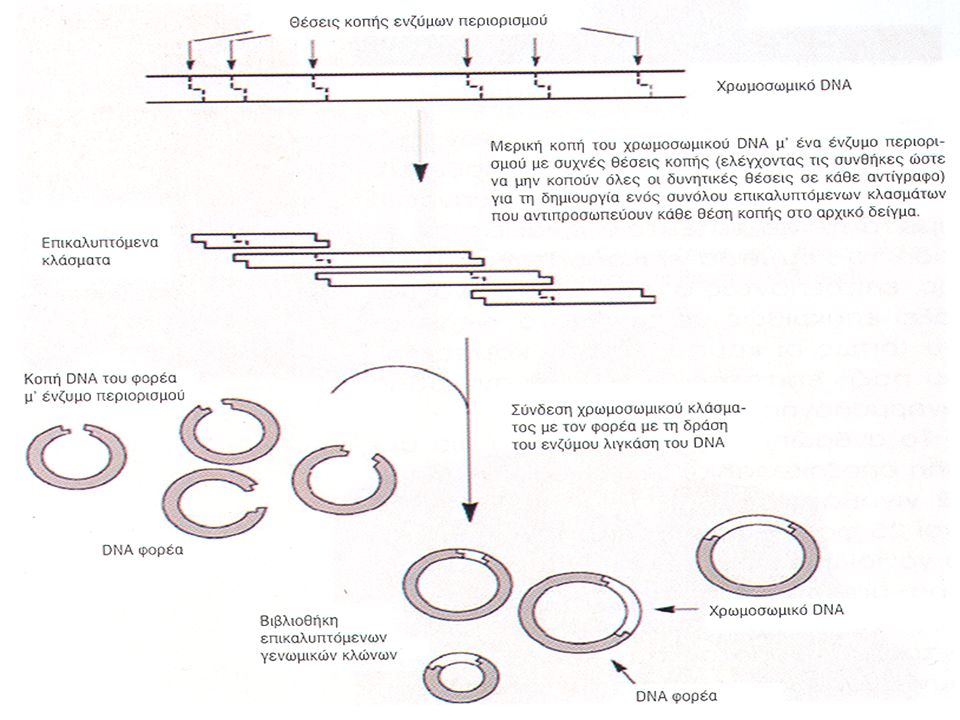
Μια άλλη μέθοδος, που ονομάζεται «τυφλός» προσδιορισμός αλληλουχίας ή «τυφλή» αλληλούχιση, η οποία έγινε εφικτή με την αυτοματοποίηση της διαδικασίας προσδιορισμού, τη ρομποτική και με ισχυρά υπολογιστικά προγράμματα, αφορά την κλωνοποίηση ολόκληρου γονιδιώματος κυριολεκτικά στην τύχη. Ο προσδιορισμός της αλληλουχίας των κλώνων γίνεται χωρίς να είναι γνωστή η σειρά ή η κατεύθυνση του κλωνοποιημένου DNA. Οι αλληλουχίες που προσδιορίζονται αναλύονται κατόπιν σε υπολογιστή, και το υπολογιστικό πρόγραμμα είναι αυτό που ψάχνει για επικαλυπτόμενες αλληλουχίες για να κατασκευάσει την πλήρη αλληλουχία του γονιδιώματος. Είναι αναγκαίο να προσδιοριστεί η αλληλουχία πολύ μεγάλου αριθμού κλώνων, πολλοί από τους οποίους είναι πανομοιότυποι. Αυτή η επταπλή ή δεκαπλή κάλυψη απομακρύνει επίσης, ή τουλάχιστον μειώνει δραστικά, την πιθανότητα σφαλμάτων κατά την αλληλούχιση.

18
Μερικές φορές, η «τυφλή» κλωνοποίηση και η τυχαία αλληλούχιση δεν μας οδηγούν σε μια πλήρη αλληλουχία. Μπορεί, δηλαδή, να υπάρχουν στην αλληλουχία κενά. Σε τέτοιες περιπτώσεις αναζητούνται κλώνοι που πιστεύεται ότι γεφυρώνουν τα κενά. Μια μέθοδος για να γίνει αυτό αν τα κενά δεν αριθμούν χιλιάδες ζεύγη βάσεων είναι η εκτέλεση αντιδράσεων PCR οι οποίες έχουν σχεδιασθεί έτσι ώστε οι δύο εκκινητές να συνδέονται σε ηδη γνωστές αλληλουχίες οι οποίες θα γεφυρώνουν τα κενά με σκοπό τον εντοπισμό και την κλωνοποίηση των τμημάτων που λείπουν. Το επόμενο βήμα στη χαρτογράφηση ενός γονιδιώματος είναι η ταυτοποίηση πιθανών γονιδίων και άλλων λειτουργικών περιοχών.

20
Εντοπισμός πιθανών γονιδίων
Τα μικροβιακά γονιδιώματα αποτελούνται συνήθως από συνεχή ανοικτά πλαίσια ανάγνωσης (open reading frames, ORF). Ο απλούστερος τρόπος εντοπισμού γονιδίων που κωδικοποιούν πρωτεΐνες, ή γονιδίων που δυνητικά κωδικοποιούν πρωτεΐνες, είναι η χρήση υπολογιστή για την ανάλυση γονιδιώματος για ORF. Η ταυτοποίηση πιθανών λειτουργικών OR F όμως, είναι κατά κανόνα πιο σύνθετη από την απλή αναζήτηση κωδικονίων έναρξης και τερματισμού εντός πλαισίου. Μια ένδειξη ότι κάποιο ORF είναι λειτουργικό αποτελεί το μέγεθος. Οι περισσότερες πρωτεΐνες συνίστανται από 100 ή και περισσότερα αμινοξέα, άρα τα περισσότερα λειτουργικά ORF είναι μεγαλύτερα από 100 κωδικόνια. Παρ’ όλα αυτά, εάν ο υπολογιστής προγραμματισθεί έτσι ώστε να αγνοεί ORF μικρότερα από 100 κωδικόνια, θα χάσει πολλά λειτουργικά γονίδια. Δεδομένου ότι πολλοί οργανισμοί προτιμούν ορισμένα από τα συνώνυμα κωδικόνια κάποιου αμινοξέως, η προτίμηση κωδικονίων αποτελεί ένδειξη για το αν ένα ORF είναι λειτουργικό..
. Ο απλούστερος τρόπος εντοπισμού γονιδίων που κωδικοποιούν πρωτεΐνες, ή γονιδίων που δυνητικά κωδικοποιούν πρωτεΐνες, είναι η χρήση υπολογιστή για την ανάλυση γονιδιώματος για ORF. Η ταυτοποίηση πιθανών λειτουργικών OR F όμως, είναι κατά κανόνα πιο σύνθετη από την απλή αναζήτηση κωδικονίων έναρξης και τερματισμού εντός πλαισίου. Μια ένδειξη ότι κάποιο ORF είναι λειτουργικό αποτελεί το μέγεθος. Οι περισσότερες πρωτεΐνες συνίστανται από 100 ή και περισσότερα αμινοξέα, άρα τα περισσότερα λειτουργικά ORF είναι μεγαλύτερα από 100 κωδικόνια. Παρ’ όλα αυτά, εάν ο υπολογιστής προγραμματισθεί έτσι ώστε να αγνοεί ORF μικρότερα από 100 κωδικόνια, θα χάσει πολλά λειτουργικά γονίδια. Δεδομένου ότι πολλοί οργανισμοί προτιμούν ορισμένα από τα συνώνυμα κωδικόνια κάποιου αμινοξέως, η προτίμηση κωδικονίων αποτελεί ένδειξη για το αν ένα ORF είναι λειτουργικό..")
21
Τα προκαρυωτικά ριβοσώματα ξεκινούν τη μετάφραση όχι από το πρώτο κατά σειρά (προς το άκρο 5΄) πιθανό κωδικόνιο έναρξης, αλλά από το πρώτο κωδικόνιο αμέσως μετά την αλληλουχία Shine-Dalgarno του mRNA Θα ήταν σχεδόν αδύνατον να συναρμολογηθεί η αλληλουχία ακόμα και ενός προκαρυωτικού γονιδιώματος και να βρεθεί η θέση των γονιδίων και άλλων σημαντικών λειτουργικών αλληλουχιών του DNA χωρίς τη χρήση σύνθετων υπολογιστικών προγραμμάτων ικανών να χειριστούν μεγάλες βάσεις δεδομένων. Η εικόνα κατωτέρω παρουσιάζει μια περιοχή DNA που περιέχει τον τύπο των ORF που αναζητούνται από το υπολογιστικό πρόγραμμα. Αν και κάθε γονίδιο μεταγράφεται πάντοτε από μία έλικα, σε όλα τα γονιδιώματα –με την εξαίρεση των μικρότερων πλασμιδίων και ιών - μεταγράφονται σε κάποιο σημείο του γονιδιώματος και οι δύο έλικες. Η εικόνα παρουσιάζει διαγραμματικά μια περιοχή γονιδιώματος όπου ένα γονίδιο μεταγράφεται από τη μία έλικα και ένα από την άλλη έλικα.

22
Γονίδια tRNA και rRNA μπορούν εύκολα να βρεθούν σε μια γονιδιωματική αλληλουχία, επειδή οι αλληλουχίες αυτών των RNA είναι σε μεγάλο βαθμό συντηρημένες. Υπάρχει επίσης πιθανότητα να είναι ένα ORF λειτουργικό, αν η αλληλουχία του εμφανίζει ομοιότητες με αλληλουχίες ORF από γονιδιώματα άλλων οργανισμών , ή αν κάποιο τμήμα του ORF διαθέτει μια αλληλουχία που γνωρίζουμε ότι κωδικοποιεί έναν λειτουργικό πρωτεϊνικό τομέα.

23
Προκαρυωτικά γονιδιώματα
Ο κατάλογος του κατωτέρω πίνακα περιλαμβάνει Αρχαία (Α) και Βακτήρια (Β) των οποίων η αλληλούχιση του γονιδιώματός έχει ολοκληρωθεί.
 και Βακτήρια (Β) των οποίων η αλληλούχιση του γονιδιώματός έχει ολοκληρωθεί.")
24
Χωρίς τη βιοπληροφορική θα ήταν αδύνατον να αναλυθούν και να συγκριθούν γονιδιώματα. Ένα από τα πρώτα βήματα στη γονιδιωματική ανάλυση είναι η σύγκριση της αλληλουχίας που προσδιορίστηκε με αλληλουχίες άλλων γονιδιωμάτων και με αλληλουχίες γονιδίων γνωστής λειτουργίας. Αυτό βοηθά ώστε να προσδιορισθεί η λειτουργία ενός γονιδίου και το ποια θα πρέπει να είναι τα βασικά γονίδια ενός συγκεκριμένου οργανισμού . Στο καλά μελετημένο βακτήριο E. coli έχουν αποδοθεί λειτουργίες σε 2700 περίπου από τα σχεδόν 4300 γονίδια. Έχουν ταυτοποιηθεί πολλά από τα γονίδια που συμμετέχουν στη σύνθεση μακρομορίων και στον κύριο μεταβολισμό. Κατά συνέπεια, καθώς ανακαλύπτονται οι λειτουργίες και άλλων μη ταυτοποιημένων πλαισίων ανάγνωσης, είναι πιθανόν να διαπιστωθεί ότι τα περισσότερα από αυτά δεν είναι απαραίτητα, οπότε θα μειωθεί το ποσοστό γονιδίων της E. Coli που συμμετέχουν σε θεμελιώδεις λειτουργίες που αφορούν τη σύνθεση μακρομορίων ή τον κύριο μεταβολισμό .

25
ΠΟΣΟΣΤΟ ΓΟΝΙΔΙΩΝ ΑΝΑ ΚΑΤΗΓΟΡΙΑ
Υπάρχει τέλος και ένα ποσοστό 46% των γονιδίων των οποίων η λειτουργία δεν έχει αποσαφηνισθεί. Η γονιδιωματική ανάλυση μπορεί επίσης να οδηγήσει στη διατύπωση νέων απόψεων σχετικά με την οικολογία ενός οργανισμού. Το Helicobacter pylori κωδικοποιεί πρωτεΐνες που περιέχουν διπλάσια ποσότητα των βασικών αμινοξέων αργινίνη και λυσίνη απ’ ότι οι τυπικές πρωτεΐνες άλλων προκαρυωτών. Προφανώς αυτό βοηθάει τα κύτταρα του H. pylori να επιβιώσουν στο όξινο περιβάλλον του στομάχου

26
Πρωτεωμική Πρωτέωμα είναι το σύνολο των πρωτεϊνών που υπάρχουν σε ένα κύτταρο, ιστό, ή οργανισμό σε οιαδήποτε χρονική στιγμή. Αν και τα γονίδια ενός οργανισμού είναι συνεχώς παρόντα και σχετικά σταθερά, λόγω της ρύθμισης της γονιδιακής έκφρασης ο αριθμός και οι τύποι των πρωτεϊνών που είναι παρούσες αλλάζουν συνεχώς αποκρινόμενες στο περιβάλλον του οργανισμού ή στην ανάπτυξή του. Η μελέτη (σε ολόκληρο το γονιδίωμα) της δομής, λειτουργίας και ρύθμισης των πρωτεϊνών ονομάζεται πρωτεωμική ή λειτουργική γονιδιωματική. Από τις πρώτες μεθόδους της πρωτεωμικής ήταν η χρήση ηλεκτροφόρησης δύο διαστάσεων σε πήκτωμα πολυακρυλαμίδης για τον διαχωρισμό, την ταυτοποίηση και την καταμέτρηση όλων των πρωτεϊνών που υπάρχουν σε ένα δείγμα κυττάρων.
 της δομής, λειτουργίας και ρύθμισης των πρωτεϊνών ονομάζεται πρωτεωμική ή λειτουργική γονιδιωματική. Από τις πρώτες μεθόδους της πρωτεωμικής ήταν η χρήση ηλεκτροφόρησης δύο διαστάσεων σε πήκτωμα πολυακρυλαμίδης για τον διαχωρισμό, την ταυτοποίηση και την καταμέτρηση όλων των πρωτεϊνών που υπάρχουν σε ένα δείγμα κυττάρων.")
27
Ένα πήκτωμα δύο διαστάσεων όπου διαχωρίστηκαν πρωτεΐνες της Escherichia coli φαίνεται στην κατωτέρω εικόνα . Στην πρώτη διάσταση (την οριζόμτια διάσταση της εικόνας), οι πρωτεΐνες διαχωρίζονται βάσει των ισοηλεκτρικών τους σημείων, δηλαδή μετακινούνται σε μια βαθμίδωση pH έως ότου το καθαρό φορτίο κάποιας συγκεκριμένης πρωτεΐνης φθάσει στο 0. Στη δεύτερη διάσταση, οι πρωτεΐνες αποδιατάσσονται κατά τρόπο ώστε κάθε αμινοξύ να αποκτά σταθερό φορτίο. Οι πρωτεΐνες τότε διαχωρίζονται βάσει μεγέθους.
, οι πρωτεΐνες διαχωρίζονται βάσει των ισοηλεκτρικών τους σημείων, δηλαδή μετακινούνται σε μια βαθμίδωση pH έως ότου το καθαρό φορτίο κάποιας συγκεκριμένης πρωτεΐνης φθάσει στο 0. Στη δεύτερη διάσταση, οι πρωτεΐνες αποδιατάσσονται κατά τρόπο ώστε κάθε αμινοξύ να αποκτά σταθερό φορτίο. Οι πρωτεΐνες τότε διαχωρίζονται βάσει μεγέθους..")
28
Μετά την αλληλούχιση του γονιδιώματος ενός οργανισμού, η προκαταρτική ανάλυση θα συγκρίνει την αλληλουχία του με αλληλουχίες άλλων οργανισμών για τη χωροθέτηση και ταυτοποίηση γονιδίων που είναι ήδη γνωστά. Η «αλληλουχία» που είναι πιο σημαντική είναι η αλληλουχία αμινοξέων της πρωτεΐνης. Λόγω του εκφυλισμού του γενετικού κώδικα, διαφορές στην αλληλουχία του DNA δεν σημαίνουν απαραιτήτως και διαφορές στην αλληλουχία των αμινοξέων . Σύγκριση αλληλουχιών νουκλεϊκών οξέων και αμινοξέων, για τον εντοπισμό ομοιοτήτων. Παρουσιάζονται τρεις διαφορετικές αλληλουχίες νουκλεοτιδίων. Τόσο η αλληλουχία 2 όσο και η 3 διαφέρουν από την αλληλουχία 1 σε τρεις θέσεις. Ωστόσο, η αλληλουχία των αμινοξέων που κωδικοποιούνται από τις 1 και 2 είναι πανομοιότυπες, ενώ αυτή που κωδικοποιείται από την 3 δεν έχει σχέση με τις άλλες δύο

29
Δομική και λειτουργική γονιδιωματική
Υπάρχουν ισχυρές ενδείξεις ότι πρωτεΐνες με αλληλουχίες όμοιες σε ποσοστό άνω του 60 – 70 % συνήθως έχουν παρεμφερείς λειτουργίες. Δομική και λειτουργική γονιδιωματική Είναι ο προσδιορισμός της τριτοταγούς δομής πρωτεϊνών και των λειτουργιών τους. Τελικός στόχος είναι η συγκέντρωση πληροφοριών όσον αφορά τη δομή, έτσι ώστε με τη βοήθεια της βιοπληροφορικής να μπορεί να προβλεφθεί η πιθανή δομή και η λειτουργία σχεδόν κάθε πρωτεΐνης, με αφετηρία την προσδιορισμένη αλληλουχία που την κωδικοποιεί. Ένας από τους σκοπούς της πρωτεωμικής είναι η μελέτη της ρύθμισης της γονιδιακής έκφρασης.

30
Στις αρχές της δεκαετίας του 1990 ανακαλύφθηκε ότι η ίδια μέθοδος
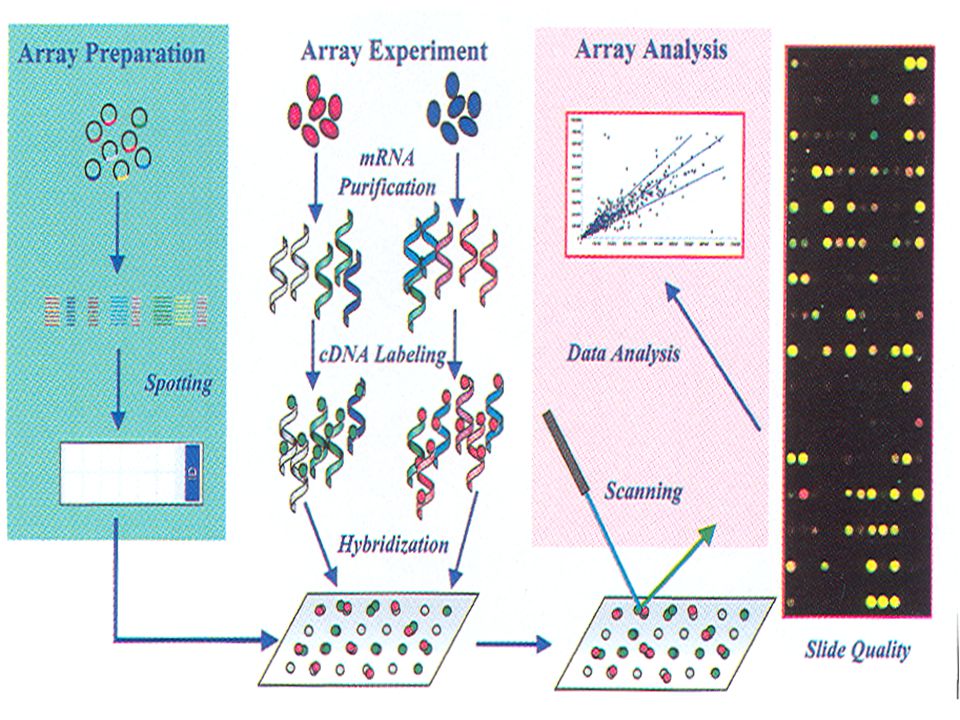
Μικροσυστοιχίες Υπάρχει πλέον διαθέσιμη μια νέα τεχνική, με μεγάλες δυνατότητες, που μας επιτρέπει να αναλύσουμε τη μεταγραφή ολόκληρου του γονιδιώματος , οι μικροσυστοιχίες. Προκειμένου να εκφραστεί ένα γονίδιο, πρέπει να μεταγραφεί. Γνωρίζοντας υπό ποιες συνθήκες μεταγράφεται ένα γονίδιο, μπορεί να αποκτήσουμε πληροφορίες και για τη λειτουργία του γονιδίου. Οι τεχνικές υβριδοποίησης έχουν επίσης χρησιμοποιηθεί επί μακρόν στη μέτρηση της γονιδιακής έκφρασης με υβριδοποίηση mRNA . Η τεχνική αυτή ενισχύεται σε μεγάλο βαθμό με την ανάπτυξη μικροσυστοιχιών, που αποτελούνται από τσιπ DNA , ( πρόκειται για τμήματα γονιδίων ή ολιγονουκλεοτιδίων αντιπροσωπευτικών του συνολικού γονιδιώματος, διατεταγμένα σε στερεή επιφάνεια ). Τα γονίδια αυτά (ή τα τμήματά τους) υβριδίζονται με διάφορους ανιχνευτές, σαρώνονται, και αναλύονται από υπολογιστή. Υπάρχουν διάφορες μέθοδοι παρασκευής μικροσυστοιχιών, μία εκ των οποίων είναι η φωτολιθογραφία . Στις αρχές της δεκαετίας του 1990 ανακαλύφθηκε ότι η ίδια μέθοδος
. Τα γονίδια αυτά (ή τα τμήματά τους) υβριδίζονται με διάφορους ανιχνευτές, σαρώνονται, και αναλύονται από υπολογιστή. Υπάρχουν διάφορες μέθοδοι παρασκευής μικροσυστοιχιών, μία εκ των οποίων είναι η φωτολιθογραφία . Στις αρχές της δεκαετίας του 1990 ανακαλύφθηκε ότι η ίδια μέθοδος.")
31
παραγωγής τσιπ ηλεκτρονικών υπολογιστών μπορούσε να προσαρμοσθεί κατάλληλα για την κατασκευή τσιπ πυριτίου, το καθένα από τα οποία μπορούσε να περιέχει μερικές χιλιάδες διαφορετικά τμήματα DNA. ΑΡΧΗ ΤΗΣ ΜΕΘΟΔΟΥ

32
Η δημιουργία ενός DNA ανιχνευτή και στη συνέχεια ο υβριδισμός σε μια επιφάνεια ή σε ένα διάλυμα με χιλιάδες αλληλουχίες ολιγονουκλεοτιδίων θα επιτρέψει την αναγνώριση και την σύνδεσή του με την συμπληρωματική του αλληλουχία . Η σήμανση του ανιχνευτή π.χ. με μια φθορίζουσα ουσία όταν αυτή διεγερθεί θα επιτρέψει μέσω του μήκους κύματος εκπομπής της να εντοπίσουμε τον ανιχνευτή και την συμπληρωματική του αλληλουχία

34
Αν η ανωτέρω αντίδραση πραγματοποιειθεί σε μια επιφάνεια ενός μικροτσιπ όπου σε κάθε mm2 έχουμε εκ των προτέρων τοποθετήσει μια σειρά γνωστών σε εμάς αλληλουχιών (δηλ. διάφορα ολιγονουκλεοτίδια ) , τότε ανάλογα με τις θέσεις όπου ο ανιχνευτής θα συνδεθεί θα μας αποκαλύψει και την αλληλουχία του.
 , τότε ανάλογα με τις θέσεις όπου ο ανιχνευτής θα συνδεθεί θα μας αποκαλύψει και την αλληλουχία του..")
36
Η εναπόθεση των “spots” γίνεται με την βοήθεια τριχοειδών πάνω στο μικροτσιπ και κάτω από αυστηρά ελεγχόμενες συνθήκες θερμοκρασίας και υγρασίας .Το σημείο εναπόθεσης και καταγραφής των “spots” καθοδηγείται και καταγράφεται από κατάλληλο λογισμικό. Σε κάθε μικροτσιπ μπορούν να τοποθετηθούν τουλάχιστον “spots”.

38
Το μεγάλο πλεονέκτημα της τεχνικής έγκειται στην δυνατότητα πραγματοποίησης μερικών δεκάδων χιλιάδων αναλύσεων ταυτόχρονα σε μια επιφάνεια η οποία δεν ξεπερνά το μέγεθος μιας γυάλινης πλάκας μικροσκοπίου , διαστάσεων 7cm X 3cm. Χρησιμοποιώντας συγχρόνως ανιχνευτές οι οποίοι έχουν σημανθεί με 2 ή 3 διαφορετικές φθορίζουσες ουσίες οι οποίες εκπέμπουν μετά την σύνδεσή τους με την συμπληρωματική τους αλληλουχία σε διαφορετικό μήκος κύματος , είναι δυνατόν ο αριθμός των αναλύσεων ταυτόχρονα να πολλαπλασιασθεί επί 2 ή 3 .

39
η σύνθεση in situ ολιγονουκλεοτιδίων μέσω τεχνικών φωτολιθογραφείας.
Ο μόνος σημαντικός προς το παρόν περιοριστικός παράγων είναι η ανάλυση της εικόνας με την υποστήριξη των κατάλληλων λογισμικών. Δύο είναι οι πιο σημαντικές μέθοδοι για την δημιουργία των μικροσυστοιχιών : Σύνδεση ολιγονουκλεοτιδίων ή τμημάτων DNA ( π.χ. αποδιατεταγμένων προιόντων PCR , cDNA , DNA ανιχνευτών ) μέσω σύνδεσης των σε επιφάνεια η οποία προηγουμένως έχει καλυφθεί από poly-L- Lysine π.χ. γυάλινη πλάκα μικροσκοπίου και η σύνθεση in situ ολιγονουκλεοτιδίων μέσω τεχνικών φωτολιθογραφείας. Το πλεονέκτημα της 1ης μεθόδου είναι το σχετικά χαμηλό κόστος κατασκευής των μικροσυστοιχειών το οποίο μπορεί να πραγματοποιηθεί σε ένα κατάλληλα επανδρωμένο εργαστήριο με εναπόθεση «spots» που κυμαίνονται περίπου στις ανά πλάκα μικροσκοπίου. Το μεγάλο μειονέκτημα της μεθόδου αυτής είναι η δημιουργία και αποθήκευση χιλιάδων ολιγονουκλεοτιδίων , προιόντων PCR , cDNA και DNA ανιχνευτών . Η 2η μέθοδος είναι καθαρά βιομηχανική και μπορεί να επιτύχει εναπόθεση «spots» της τάξης των σε μια επιφάνεια 1,2Χ1,2 cm.
 μέσω σύνδεσης των σε επιφάνεια η οποία προηγουμένως έχει καλυφθεί από poly-L- Lysine π.χ. γυάλινη πλάκα μικροσκοπίου και. η σύνθεση in situ ολιγονουκλεοτιδίων μέσω τεχνικών φωτολιθογραφείας. Το πλεονέκτημα της 1ης μεθόδου είναι το σχετικά χαμηλό κόστος κατασκευής των μικροσυστοιχειών το οποίο μπορεί να πραγματοποιηθεί σε ένα κατάλληλα επανδρωμένο εργαστήριο με εναπόθεση «spots» που κυμαίνονται περίπου στις ανά πλάκα μικροσκοπίου. Το μεγάλο μειονέκτημα της μεθόδου αυτής είναι η δημιουργία και αποθήκευση χιλιάδων ολιγονουκλεοτιδίων , προιόντων PCR , cDNA και DNA ανιχνευτών . Η 2η μέθοδος είναι καθαρά βιομηχανική και μπορεί να επιτύχει εναπόθεση «spots» της τάξης των σε μια επιφάνεια 1,2Χ1,2 cm.")
40
Κατασκευή μιας «in house» μικροσυστοιχίας με εναπόθεση 17
Κατασκευή μιας «in house» μικροσυστοιχίας με εναπόθεση spots cDNA σε μικροπλάκα μικροσκοπίου. Το USA penny στα αριστερά προσδιορίζει την κλίμακα.

42
ΦΡΙΖΟΥΣΕΣ ΟΥΣΙΕΣ ΟΙ ΟΠΟΙΕΣ ΧΡΗΣΙΜΟΠΟΙΟΥΝΤΑΙ ΣΤΙΣ ΜΙΚΡΟΣΥΣΤΟΙΧΙΕΣ ΚΑΙ ΤΑ ΧΑΡΑΚΤΗΡΙΣΤΙΚΑ ΑΥΤΩΝ

43
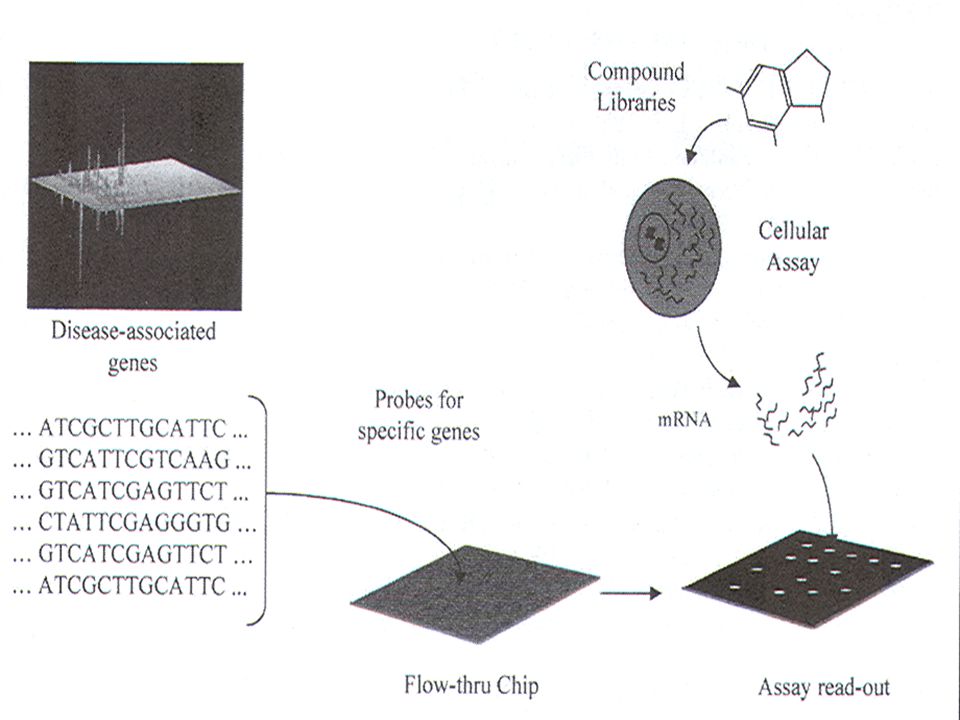
Κατωτέρω απεικονίζεται η δημιουργία μικροσυστοιχιών διαφορετικών τμημάτων DNA , γνωστής αλληλουχίας , σε μια συστοιχία τσιπ πυριτίου. Μονόκλωνα ολιγονουκλεοτίδια μικρού μήκους, που αντιστοιχούν σε όλα τα γονίδια ενός οργανισμού, συντίθενται μεμονωμένα και προσκολλώνται σε προκαθορισμένες και ταυτοποιήσιμες θέσεις . Στην κατωτέρω εικόνα χρησιμοποιείται ένα μόνο γονίδιο. Αυτά τα τσιπ DNA αναλύονται με τη χρήση σημασμένων ανιχνευτών από τον υπό μελέτη οργανισμό, υπό ποικίλες συνθήκες, με στόχο τον προσδιορισμό προτύπων γονιδιακής έκφρασης.

44
Οι μικροσυστοιχίες υβριδίζονται με ανιχνευτές DNA ή RNA, σημασμένους με φθορίζουσα χρωστική . Το σημασμένο DNA ή RNA προσδένεται μόνο στην αλληλουχία του τσιπ που είναι συμπληρωματική του. Το τσιπ σαρώνεται με λέιζερ και αναλύεται σε ηλεκτρονικό υπολογιστή με την βοήθεια των κατάλληλων λογισμικών. Ανάλογα με το ποιες συμπληρωματικές αλληλουχίες DNA ή RNA υπάρχουν στο δείγμα, θα παρατηρηθούν και συγκεκριμένα πρότυπα υβριδισμού

45
Για παράδειγμα, μπορεί κάποιος να θέσει το ερώτημα ποια από τα 5000 και πλέον γονίδια του Saccharomyces cerevisae εκφράζονται υπό συγκεκριμένες συνθήκες, χρησιμοποιώντας το σύνολο του mRNA ως ανιχνευτή. Βεβαίως, μπορεί κάποιος να συγκρίνει και την έκφραση διαφορετικών γονιδίων υπό διαφορετικές συνθήκες. Στην κατωτέρω εικόνα απεικονίζεται ένα τμήμα μικροσυστοιχίας η οποία χρησιμοποιήθηκε για την ανάλυση της έκφρασης του γονιδιώματος του Saccharomyces cerevisiae.

46
Ο μέγιστος υβριδισμός 100% δίνει λευκό χρώμα.
Κάθε γονίδιο υπάρχει σε πολλαπλά αντίγραφα και έχει ανιχνευθεί με mRNA σημασμένο με φθορίζουσα χρωστική το οποίο απομονώθηκε από κύτταρα ζύμης που αναπτύχθηκαν κάτω από διάφορες συνθήκες. Οι θέσεις όπου το RNA υβριδίζεται με το DNA εμφανίζονται με διαφορετική απόχρωση . Ο μέγιστος υβριδισμός 100% δίνει λευκό χρώμα. Δεδομένου ότι είναι γνωστή η θέση των διαφόρων γονιδίων πάνω στο μικροτσιπ , μετά την σάρωσή του και την επεξεργασία της εικόνας με το κατάλληλο λογισμικό θα φανεί ποια γονίδια εκφράζονται και κάτω από ποίες συνθήκες.

47
Παρακάτω παρουσιάζεται ένα παράδειγμα μελέτης της έκφρασης γονιδίων χρησιμοποιώντας cDNA το οποίο έχει σημανθεί κατά την διάρκεια της αντίστροφης μεταγραφής ενσωματώνοντας είτε την φθορίζουσα ουσία Cy3 ( πράσινο χρώμα ) είτε την Cy5 ( κόκκινο χρώμα). Τα δυο cDNA υβριδίζονται στην μικροσυστοιχία η οποία σκανάρεται και αναλύεται με το κατάλληλο λογισμικό.Οταν το πράσινο χρώμα αναμειγνύεται με το κόκκινο σε ίση συγκέντρωση τότε προκύπτει κίτρινο χρώμα . Το λογισμικό αναλύει την ένταση του πράσινου, κόκκινου , κίτρινου και των διαβαθμίσεων του και υπολογίζει τον log του λόγου της έντασης του Cy5/Cy3. Θετικός λογάριθμος σημαίνει περισσότερη έκφραση του μετάγραφου που έχει σημανθεί με το Cy5 , αρνητικός λογάριθμος σημαίνει περισσότερη έκφραση του μετάγραφου που έχει σημανθεί με το Cy3 και λογάριθμος = 0 σημαίνει ίδια έκφραση και των 2 μετάγραφων.

48
Παρουσιάζεται επίσης μια βαθμίδωση της έντασης των ανωτέρω χρωμάτων
Πραγματοποιώντας ανάλογα πειράματα δεκάδες φορές είναι δυνατόν πλέον τα αποτελέσματα να παρουσιασθούν υπό μορφή ανάλυσης CLUSTER. Tα κόκκινα τετράγωνα απεικονίζουν θετικό λογάριθμο του λόγου Cy5/Cy3 , τα πράσινα αρνητικό λογάριθμο του λόγου Cy5/Cy3 , τα μαύρα ότι ο ανωτέρω λόγος =0 . Παρουσιάζεται επίσης μια βαθμίδωση της έντασης των ανωτέρω χρωμάτων

49
Το clustering των γονιδίων απεικονίζεται στον κάθετο άξονα και τα διάφορα πειράματα στον οριζόντιο άξονα.Μέσω αυτής της ανάλυσης είναι δυνατόν να εντοπισθούν γονίδια τα οποία συν-ρυθμίζουν την έκφρασή τους.

50
Κατωτέρω απεικονίζεται η έκφραση 600 γονιδίων ( οριζόντιος άξονας) κατά την διάρκεια 21 πειραμάτων ( κάθετος άξονας). Το clustering της έκφρασης των γονιδίων σε σχέση με τις συνθήκες των 21 πειραμάτων απεικονίζονται μέσω των δενδρογραμμάτων ( οριζόντια και κάθετα ).
..")
51
Η δυνατότητα σχετικά εύκολης ανάλυσης της ταυτόχρονης έκφρασης χιλιάδων γονιδίων, τόσο ποιοτικά όσο και ποσοτικά, προσφέρει τεράστιες δυνατότητες στο να κατανοήσουμε την απίστευτη περιπλοκότητα και τη ρύθμιση οργανισμών τόσο «απλών» όσο η Escherichia coli με τα περίπου γονίδιά της, ή τόσο σύνθετους οργανισμούς με περίπου γονίδια όπως ο άνθρωπος. Μερικές Εφαρμογές Σχεδιάζοντες εκκινητές οι οποίοι ανιχνεύουν ORFs και λαμβάνοντας τα αντίστοιχα προιόντα PCR είναι δυνατόν να κατασκευάσουμε μικροσυστοιχίες και να εντοπίσουμε κατ’αρχάς τα γονίδια και εν συνεχεία μελετήσουμε την έκφραση των γονιδίων ενός μικροοργανισμού ή και ενός ανώτερου οργανισμού κάτω από συγκεκριμένες πειραματικές συνθήκες. Η προσέγγιση αυτή έχει ήδη εφαρμοσθεί για τον εντοπισμό των γονιδίων της Ε. coli και των γονιδίων του Mycobacterium tuberculosis .Μελέτες ανάπτυξης της Ε. coli σε πλούσιο και φτωχό θρεπτικό υπόστρωμα απέδειξαν ότι 344 γονίδια εκφράζονται διαφορετικά ανάλογα το θρεπτικό υπόστρωμα , γεγονώς που δηλώνει ότι στο άμεσο μέλλον από το “pattern” μικροσυστοιχίας θα προβλέπουμε την φυσιολογική κατάσταση του εκάστοτε εξεταζόμενου μικροοργανισμού.

52
εκάστοτε εξεταζόμενου μικροοργανισμού.
Δεδομένου ότι η έκφραση των γονιδίων τα οποία συνδέονται με την λοιμογόνο δύναμη παθογόνων μικροοργανισμών είναι συνάρτησει του μικροπεριβάλλοντος του μικροοργανισμού , οι μικροσυστοιχίες προσφέρουν την δυνατότητα όχι μόνο να εντοπισθούν νέα τέτοια λοιμογόνα γονίδια αλλά και κάτω από ποιες συνθήκες αυτά εκφράζονται. Οι μικροσυστοιχίες μπορούν να χρησιμοποιηθούν επίσης στην ταυτοποίηση μικροοργανισμών. Χρησιμοποιώντας ολιγονουκλεοτίδια «υπογραφές» καθώς και ολιγονουκλεοτίδια από μερικώς συντηρημένες και ευμετάβλητες αλληλουχίες είναι δυνατόν μέσω διαφορών στα πρότυπα υβριδισμού να επιτευχθεί άμεσα ο προσδιορισμός του γένους, του είδους και να γίνει ακόμη και διαφοροποίηση στενά συγγενικών στελεχών. Αυτό επιτρέπει την ταχύτατη ταυτοποίηση παθογόνων στελεχών ιών ή βακτηρίων και ακόμη περαιτέρω την ανίχνευση μεταλλαγμάτων των συγκεκριμένων μικροοργανισμών χωρίς να απαιτείται η χρονοβόρος διαδικασία ανάπτυξης του μικροοργανισμού σε κατάλληλα θρεπτικά υποστρώματα ή κυτταροκαλλιέργειες

53
Μελέτη της παθογένειας των μικροοοργανισμών
Μελέτη της παθογένειας των μικροοοργανισμών. Οι μικροσυστοιχίες επιτρέπουν , προς το παρόν τουλάχιστον in vitro , την ομαδοποίηση γονιδίων μικροοργανισμών τα οποία είτε ενεργοποιούνται είτε καταστέλλονται κατά την διάρκεια μιας μόλυνσης.Π.χ. η μόλυνση μιας ευαίσθητης κυτταρικής σειράς (MRC5) για την ανάπτυξη του CMV η οποία μελετήθηκε με μικροσυστοιχίες 8 και ώρες μετά την μόλυνση με τον CMV , έδειξε την υπερ-έκφραση 258 εκ των γονιδίων τα οποία εμελετήθηκαν σε σχέση με τα ίδια γονίδια στην κυτταρική σειρά η οποία δεν είχε μολυνθεί από τον ιό. Τέτοιες μελέτες ανοίγουν τον δρόμο για την Φαρμακογενωμική. Μελέτη της επίδρασης διαφόρων φαρμακευτικών παρασκευασμάτων μέσω του “pattern” έκφρασης των διαφόρων γονιδίων του μικροοργανισμού πριν ,κατά την διάρκεια χορήγησης και μετά το πέρας χορήγησης του παρασκευάσματος. Πρόσφατα η μελέτη αυτή πραγματοποιήθηκε για το Mycobacterium tuberculosis έναντι των αναστολέων της σύνθεσης των μυκολικών οξέων ISONIAZID και ETHIONAMIDE. Οι ανωτέρω αναστολείς παρουσίασαν ένα παρόμοιο “pattern” έκφρασης το οποίο χαρακτηρίσθηκε από την επαγωγή της μεταγραφής 5 συνεχόμενων γονιδίων τα οποία εμπλέκονται στην βιοσύνθεση ενζύμων των λιπαρών οξέων.
 για την ανάπτυξη του CMV η οποία μελετήθηκε με μικροσυστοιχίες 8 και 24 ώρες μετά την μόλυνση με τον CMV , έδειξε την υπερ-έκφραση 258 εκ των γονιδίων τα οποία εμελετήθηκαν σε σχέση με τα ίδια γονίδια στην κυτταρική σειρά η οποία δεν είχε μολυνθεί από τον ιό. Τέτοιες μελέτες ανοίγουν τον δρόμο για την Φαρμακογενωμική. Μελέτη της επίδρασης διαφόρων φαρμακευτικών παρασκευασμάτων μέσω του pattern έκφρασης των διαφόρων γονιδίων του μικροοργανισμού πριν ,κατά την διάρκεια χορήγησης και μετά το πέρας χορήγησης του παρασκευάσματος. Πρόσφατα η μελέτη αυτή πραγματοποιήθηκε για το Mycobacterium tuberculosis έναντι των αναστολέων της σύνθεσης των μυκολικών οξέων ISONIAZID και ETHIONAMIDE. Οι ανωτέρω αναστολείς παρουσίασαν ένα παρόμοιο pattern έκφρασης το οποίο χαρακτηρίσθηκε από την επαγωγή της μεταγραφής 5 συνεχόμενων γονιδίων τα οποία εμπλέκονται στην βιοσύνθεση ενζύμων των λιπαρών οξέων.")
54
Κατά συνέπεια το “pattern” έκφρασης μιας μικροσυστοιχίας μπορεί να μας καθοδηγήσει για την ανάπτυξη νέων φαρμακευτικών παρασκευασμάτων όπως στο ανωτέρω παράδειγμα όπου θα πρέπει να αναζητηθούν και νέοι ειδικοί αναστολής της σύνθεσης των λιπαρών οξέων του Mycobacterium tuberculosis . Και βέβαια η τεχνική αυτή προσφέρεται για τον μαζικό έλεγχο της φαρμακευτικής δράσης χιλιάδων παρασκευασμάτων και τον ταχύτατο εντοπισμό μεταλλάξεων σε γονίδια οι οποίες μεταλλάξεις προσδίδουν ανθεκτικότητα σε αντιβιοτικά (π.χ. μεταλλάξεις στο γονίδιο rpoB του Mycobacterium tuberculosis προσδίδει ανθεκτικότητα στην rifampicin ).
..")
Παρόμοιες παρουσιάσεις























 - Γ' ΦΑΣΗ Αποθετήρια και Επιστημονικά Ηλεκτρονικά Περιοδικά Ανοικτής Πρόσβασης»>")