Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
ΕΙΣΑΓΩΓΗ Η επικοινωνία μπορεί να εκδηλώνεται από απόσταση ή από κοντά. Στην πρώτη περίπτωση αναφερόμαστε στην ύπαρξη ενός ενδιάμεσου μορίου που χρησιμοποιείται ως μήνυμα μεταφοράς της πληροφορίας ή της εντολής. Στη δεύτερη περίπτωση αναφερόμαστε σε σύζευξη κυττάρων που έχει διαπιστωθεί με διάφορους τρόπους, όπως με μικροηλεκτρόδια, με φθορίζουσες ουσίες ή με ραδιενεργές ουσίες. ΕΙΣΑΓΩΓΗ

2
ΣΥΝΔΕΣΜΟΙ ΕΠΙΚΟΙΝΩΝΙΑΣ (COMMUNICATING JUNCTIONS) 1
ΣΥΝΔΕΣΜΟΙ ΕΠΙΚΟΙΝΩΝΙΑΣ (COMMUNICATING JUNCTIONS) 1.Χασμοσύνδεσμοι (gap junctions) 2.Χημικές συνάψεις (chemical synapses) 3.Πλασμοδέσμες (plasmodesmata) - (μόνο στα φυτά) ΦΡΑΓΜΟΣΥΝΔΕΣΜΟΙ (OCCLUDING JUNCTIONS) 1.Στενοσύνδεσμοι (tight junctions) - (μόνο στα σπονδυλωτά) 2.Διαφραγματοσύνδεσμοι (septate junctions) - (κυρίως στα ασπόνδυλα) ΣΥΝΔΕΣΜΟΙ ΠΡΟΣΔΕΣΗΣ (ANCHORING JUNCTIONS) Συνδεόμενοι με ενδοκυττάρια ινίδια ακτίνης 1.Σύνδεσμος προσκόλλησης (adherens junctions) - ( cell-cell junctions) 2.Εστίες προσκόλλησης (focal adhesions) – (cell-matrix junctions) Συνδεόμενοι με ενδοκυττάρια ενδιάμεσα ινίδια 1.Δεσμοσώματα (desmosomes) - (cell-cell junctions) 2.Ημιδεσμοσώματα (hemidesmosomes)- (cell-matrix junctions)
 1.Χασμοσύνδεσμοι (gap junctions) 2.Χημικές συνάψεις (chemical synapses) 3.Πλασμοδέσμες (plasmodesmata) - (μόνο στα φυτά) ΦΡΑΓΜΟΣΥΝΔΕΣΜΟΙ (OCCLUDING JUNCTIONS) 1.Στενοσύνδεσμοι (tight junctions) - (μόνο στα σπονδυλωτά) 2.Διαφραγματοσύνδεσμοι (septate junctions) - (κυρίως στα ασπόνδυλα) ΣΥΝΔΕΣΜΟΙ ΠΡΟΣΔΕΣΗΣ (ANCHORING JUNCTIONS) Συνδεόμενοι με ενδοκυττάρια ινίδια ακτίνης 1.Σύνδεσμος προσκόλλησης (adherens junctions) - ( cell-cell junctions) 2.Εστίες προσκόλλησης (focal adhesions) – (cell-matrix junctions) Συνδεόμενοι με ενδοκυττάρια ενδιάμεσα ινίδια 1.Δεσμοσώματα (desmosomes) - (cell-cell junctions) 2.Ημιδεσμοσώματα (hemidesmosomes)- (cell-matrix junctions)")
3
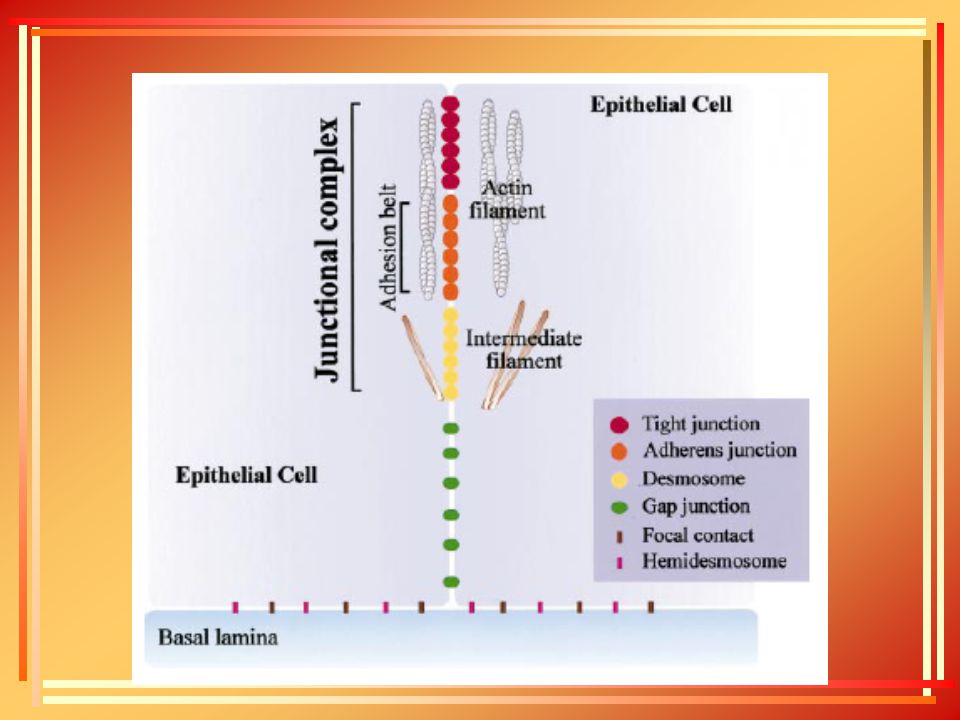
ΕΙΚΟΝΑ 1. ΣΥΝΟΨΗ ΤΩΝ ΔΙΑΦΟΡΩΝ ΚΥΤΤΑΡΙΚΩΝ ΣΥΝΔΕΣΜΩΝ ΠΟΥ ΠΑΡΑΤΗΡΟΥΝΤΑΙ ΣΕ ΕΝΑ ΕΠΙΘΗΛΙΑΚΟ ΚΥΤΤΑΡΟ ΤΟΥ ΛΕΠΤΟΥ ΕΝΤΕΡΟΥ ΣΠΟΝΔΥΛΩΤΟΥ.

4
Ο σχηματισμός όμως των συνδέσμων προϋποθέτει το σχηματισμό των αρχικών κυτταρικών επαφών που γίνονται με τη μεσολάβηση των πρωτεϊνών προσκόλλησης κυτταρικής επιφάνειας. Στις πρωτεϊνες κυτταρικής προσκόλλησης ανήκουν οι καδερίνες και οι N-CAM πρωτεϊνες. Η μεθοδολογία που έχει χρησιμοποιηθεί στις μελέτες αυτές είναι η ηλεκτρονική μικροσκοπία λεπτών τομών, η ψυκτοεξάχνωση και η περίθλαση με ακτίνες-Χ

5
ΣΥΝΔΕΣΜΟΙ ΕΠΙΚΟΙΝΩΝΙΑΣ
ΣΥΝΔΕΣΜΟΙ ΕΠΙΚΟΙΝΩΝΙΑΣ

6
ΧΑΣΜΟΣΥΝΔΕΣΜΟΙ Τα περισσότερα κύτταρα στους ζωϊκούς ιστούς επικοινωνούν με τα γειτονικά τους μέσω των χασμοσυνδέσμων. Κάθε χασμοσύνδεσμος εμφανίζεται εκεί όπου οι μεμβράνες των δύο γειτονικών κυττάρων χωρίζονται από ένα χάσμα 2-4 nm περίπου. Στο χάσμα αυτό δημιουργούνται δίαυλοι (κοννεξόνια) από τις πρωτεϊνες κοννεξίνες, οι οποίοι επιτρέπουν σε ανόργανα ιόντα και σε άλλα μικρά υδρόφιλα μόρια όπως σάκχαρα, αμινοξέα, νουκλεοτίδια, βιταμίνες και διακυτταρικούς μεσολαβητές να περάσουν απ’ευθείας από το κυτταρόπλασμα του ενός κυττάρου στο κυτταρόπλασμα του άλλου. Βρέθηκε ότι ο πόρος του διαύλου στα κύτταρα θηλαστικών έχει διάμετρο περίπου nm . Έτσι, δεν επιτρέπεται η διέλευση μακρομορίων όπως πρωτεϊνες, νουκλεϊνικά οξέα και πολυσακχαρίτες. Γενικά μόρια με μοριακό βάρος μικρότερο του 1–1.2 kDa διέρχονται ελεύθερα, ενώ εκείνα από 2 kDa ή περισσότερο δεν περνάνε.
 από τις πρωτεϊνες κοννεξίνες, οι οποίοι επιτρέπουν σε ανόργανα ιόντα και σε άλλα μικρά υδρόφιλα μόρια όπως σάκχαρα, αμινοξέα, νουκλεοτίδια, βιταμίνες και διακυτταρικούς μεσολαβητές να περάσουν απ’ευθείας από το κυτταρόπλασμα του ενός κυττάρου στο κυτταρόπλασμα του άλλου. Βρέθηκε ότι ο πόρος του διαύλου στα κύτταρα θηλαστικών έχει διάμετρο περίπου nm . Έτσι, δεν επιτρέπεται η διέλευση μακρομορίων όπως πρωτεϊνες, νουκλεϊνικά οξέα και πολυσακχαρίτες. Γενικά μόρια με μοριακό βάρος μικρότερο του 1–1.2 kDa διέρχονται ελεύθερα, ενώ εκείνα από 2 kDa ή περισσότερο δεν περνάνε.")
7
ΕΙΚΟΝΑ 2. ΠΡΟΣΔΙΟΡΙΣΜΟΣ ΤΟΥ ΜΕΓΕΘΟΥΣ ΤΟΥ ΔΙΑΥΛΟΥ ΤΟΥ ΧΑΣΜΟΣΥΝΔΕΣΜΟΥ
ΕΙΚΟΝΑ 2. ΠΡΟΣΔΙΟΡΙΣΜΟΣ ΤΟΥ ΜΕΓΕΘΟΥΣ ΤΟΥ ΔΙΑΥΛΟΥ ΤΟΥ ΧΑΣΜΟΣΥΝΔΕΣΜΟΥ. Όταν φθορίζοντα μόρια διαφόρου μεγέθους ενέσονται σε δύο κύτταρα που συνδέονται με χασμοσυνδέσμους, μόρια με Μ.Β. μικρότερο των daltons μπορούν να διέλθουν από το ένα στο άλλο κύτταρο, ενώ μεγαλύτερα μόρια δεν μπορούν.

8
ΔΟΜΗ ΤΩΝ ΔΙΑΥΛΩΝ ΣΤΟΥΣ ΧΑΣΜΟΣΥΝΔΕΣΜΟΥΣ Ένας δίαυλος χασμοσυνδέσμου δημιουργείται από μόρια μιας διαμεμβρανικής πρωτεϊνης, της κοννεξίνης. Το Μ.Β. της κυμαίνεται από 30 εώς 42 kD. Έξι μόρια κοννεξίνης συναρμολογούνται για να δημιουργήσουν έναν δίαυλο, το κοννεξόνιο. Όταν τα κοννεξόνια στις πλασματικές μεμβράνες των δύο κυττάρων που βρίσκονται σε επαφή ευθυγραμμιστούν, σχηματίζουν έναν συνεχή δίαυλο ύδατος, ο οποίος συνδέει το εσωτερικό των δύο κυττάρων. Κάθε πολυπεπτίδιο κοννεξίνης έχει προταθεί ότι διέρχεται τέσσερις φορές από την πλασματική μεμβράνη και ότι μια συντηρητική ως προς την αμινοξική αλληλουχία διαμεμβρανική α-έλικα επενδύει το δίαυλο ύδατος.

9
ΕΙΚΟΝΑ 3. ΧΑΣΜΟΣΥΝΔΕΣΜΟΙ
ΕΙΚΟΝΑ 3. ΧΑΣΜΟΣΥΝΔΕΣΜΟΙ . (Α) Τρισδιάστατο σχέδιο που δείχνει την αλληλεπίδραση των πλασματικών μεβρανών των δύο γειτονικών κυττάρων που συνδέονται με χασμοσυνδέσμους. Οι παρατηθέμενες λιπιδικές διπλοστοιβάδες (κόκκινες) διαπερνούνται από συναρμολογούμενες πρωτεϊνες που ονομάζονται κοννεξόνια (πράσινα), καθένα από τα οποία αποτελείται από έξι υπομονάδες κοννεξίνης. Δύο κοννεξόνια ενώνονται για να δημιουργήσουν ένα συνεχή δίαυλο ύδατος που ενώνει τα δύο κύτταρα. (Β) Η οργάνωση των κοννεξινών στα κοννεξόνια στους διακυτταρικούς διαύλους. Τα κοννεξόνια μπορούν να είναι ομομερικά ή ετερομερικά και οι διακυτταρικοί δίαυλοι ομοτυπικοί ή ετεροτυπικοί.
 Τρισδιάστατο σχέδιο που δείχνει την αλληλεπίδραση των πλασματικών μεβρανών των δύο γειτονικών κυττάρων που συνδέονται με χασμοσυνδέσμους. Οι παρατηθέμενες λιπιδικές διπλοστοιβάδες (κόκκινες) διαπερνούνται από συναρμολογούμενες πρωτεϊνες που ονομάζονται κοννεξόνια (πράσινα), καθένα από τα οποία αποτελείται από έξι υπομονάδες κοννεξίνης. Δύο κοννεξόνια ενώνονται για να δημιουργήσουν ένα συνεχή δίαυλο ύδατος που ενώνει τα δύο κύτταρα. (Β) Η οργάνωση των κοννεξινών στα κοννεξόνια στους διακυτταρικούς διαύλους. Τα κοννεξόνια μπορούν να είναι ομομερικά ή ετερομερικά και οι διακυτταρικοί δίαυλοι ομοτυπικοί ή ετεροτυπικοί.")
10
ΕΙΚΟΝΑ 4. ΧΑΣΜΟΣΥΝΔΕΣΜΟΙ ΟΠΩΣ ΦΑΙΝΟΝΤΑΙ ΣΤΟ ΗΛΕΚΤΡΟΝΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ
ΕΙΚΟΝΑ 4. ΧΑΣΜΟΣΥΝΔΕΣΜΟΙ ΟΠΩΣ ΦΑΙΝΟΝΤΑΙ ΣΤΟ ΗΛΕΚΤΡΟΝΙΚΟ ΜΙΚΡΟΣΚΟΠΙΟ. (Α) Λεπτή τομή και (Β) Ηλεκτρονική μικρογραφία με ψυκτοεξάχνωση ενός μεγάλου και ενός μικρού χασμοσυνδέσμου μεταξύ ινοβλαστών σε καλλιέργεια.
 Λεπτή τομή και (Β) Ηλεκτρονική μικρογραφία με ψυκτοεξάχνωση ενός μεγάλου και ενός μικρού χασμοσυνδέσμου μεταξύ ινοβλαστών σε καλλιέργεια.")
11
ΕΦΑΡΜΟΓΕΣ ΤΩΝ ΧΑΣΜΟΣΥΝΔΕΣΜΩΝ
φαινόμενο της μεταβολικής σύζευξης ή μεταβολικής συνεργασίας: ένα κύτταρο μπορεί να μεταφέρει μόρια σε ένα γειτονικό κύτταρο που είναι ανίκανο να τα συνθέσει. Π.χ. το AMP, το ADP ή το ATP μπορούν να περάσουν μέσα από χασμοσυνδέσμους. η φυσιολογική ανάπτυξη των ωοθηκικών αδένων, εξαρτάται επίσης από την επικοινωνία μέσω των χασμοσυνδέσμων, μεταξύ των ωοκυττάρων και των κοκκιωδών κυττάρων που τα περιβάλλει. Μια μεταλλαγή στο γονίδιο που κωδικοποιεί την κοννεξίνη, που φυσιολογικά συνδέει αυτούς τους δύο τύπους κυττάρων, προκαλεί στειρότητα. η εφαρμογή της ένεσης του mRNA της κοννεξίνης, είναι πολύ χρήσιμη για τον προσδιορισμό καινούργιων χασμοσυνδετικών πρωτεϊνών. Γενετικές μελέτες στην μύγα Drosophila ταυτοποίησαν το γονίδιο shaking B. Η Shaking B έγινε το πρώτο μέλος μιας νέας οικογένειας χασμοσυνδετικών πρωτεϊνών των ασπονδύλων, που ονομάζονται ιννεξίνες. Υπάρχουν περισσότερα από 15 γονίδια ιννεξίνης στην Drosophila και 25 στον νηματοειδή C.elegans.

12
ΡΥΘΜΙΣΗ ΤΗΣ ΔΙΑΠΕΡΑΤΟΤΗΤΑΣ ΤΩΝ ΧΑΣΜΟΣΥΝΔΕΣΜΩΝ
Οι δίαυλοι χασμοσυνδέσμων είναι δυναμικές δομές και μεταπηδούν συχνά από τη μια κατάσταση στην άλλη (ανοιχτοί δίαυλοι – κλειστοί δίαυλοι). Το πέρασμα διαμέσου των χασμοσυνδέσμων είναι γρήγορο (μέσα σε δευτερόλεπτα) και η διαπερατότητά τους αντιστρεπτή. Μειώνεται όταν ελαττώνεται το κυττοσολικό pH ή αυξάνεται κατά πολύ η κυττοσολική συγκέντρωση ελεύθερου Ca2+. Ο σκοπός της ρύθμισης του pH για τη διαπερατότητα των χασμοσυνδέσμων δεν είναι γνωστός. Σε μια περίπτωση, όμως, ο σκοπός του ελέγχου του Ca2+ μοιάζει να είναι ξεκάθαρος. Όταν ένα κύτταρο έχει καταστραφεί, η πλασματική του μεμβράνη γίνεται διάτρητη =>ιόντα σε υψηλές συγκεντρώσεις στο εξωκυτταρικό υγρό (π.χ Ca2+ και Na+) μεταφέρονται μέσα στο κύτταρο και πολύτιμοι μεταβολίτες εξέρχονται από αυτό. Αλλά η μεγάλη εισροή Ca2+ μέσα στο κατεστραμμένο κύτταρο => κλείσιμο χασμοσυνδετικών διαύλων => εμποδίζεται η καταστροφή και των γειτονικών κυττάρων. Η επικοινωνία μέσω χασμοσυνδέσμων μπορεί να ρυθμιστεί και μέσω εξωκυτταρικών σινιάλων. Για παράδειγμα η ντοπαμίνη, μια νευροδιαβιβαστική πρωτεϊνη, μειώνει την χασμοσυνδετική επικοινωνία μεταξύ μιας τάξης νευρώνων στον αμφιβληστροειδή, ως απόκριση στην αύξηση της φωτεινής έντασης.
. Το πέρασμα διαμέσου των χασμοσυνδέσμων είναι γρήγορο (μέσα σε δευτερόλεπτα) και η διαπερατότητά τους αντιστρεπτή. Μειώνεται όταν ελαττώνεται το κυττοσολικό pH ή αυξάνεται κατά πολύ η κυττοσολική συγκέντρωση ελεύθερου Ca2+. Ο σκοπός της ρύθμισης του pH για τη διαπερατότητα των χασμοσυνδέσμων δεν είναι γνωστός. Σε μια περίπτωση, όμως, ο σκοπός του ελέγχου του Ca2+ μοιάζει να είναι ξεκάθαρος. Όταν ένα κύτταρο έχει καταστραφεί, η πλασματική του μεμβράνη γίνεται διάτρητη =>ιόντα σε υψηλές συγκεντρώσεις στο εξωκυτταρικό υγρό (π.χ Ca2+ και Na+) μεταφέρονται μέσα στο κύτταρο και πολύτιμοι μεταβολίτες εξέρχονται από αυτό. Αλλά η μεγάλη εισροή Ca2+ μέσα στο κατεστραμμένο κύτταρο => κλείσιμο χασμοσυνδετικών διαύλων => εμποδίζεται η καταστροφή και των γειτονικών κυττάρων. Η επικοινωνία μέσω χασμοσυνδέσμων μπορεί να ρυθμιστεί και μέσω εξωκυτταρικών σινιάλων. Για παράδειγμα η ντοπαμίνη, μια νευροδιαβιβαστική πρωτεϊνη, μειώνει την χασμοσυνδετική επικοινωνία μεταξύ μιας τάξης νευρώνων στον αμφιβληστροειδή, ως απόκριση στην αύξηση της φωτεινής έντασης.")
13
ΣΥΝΑΨΕΙΣ Ειδική μορφή κυτταρικής επικοινωνίας αποτελούν οι συνάψεις που βρίσκονται σε επαφές νευρικών κυττάρων ή νευρικών με μυϊκά κύτταρα. Διακρίνονται σε ηλεκτρικές και σε χημικές συνάψεις.

14
ΕΙΚΟΝΑ 6. ΟΙ ΗΛΕΚΤΡΙΚΕΣ ΚΑΙ ΟΙ ΧΗΜΙΚΕΣ ΣΥΝΑΨΕΙΣ ΔΙΑΦΕΡΟΥΝ ΣΗΜΑΝΤΙΚΑ ΣΤΟΝ ΜΗΧΑΝΙΣΜΟ ΜΕΤΑΦΟΡΑΣ. (Α) Στις ηλεκτρικές συνάψεις, οι χασμοσύνδεσμοι μεταξύ προ- και μετασυναπτικών μεμβρανών επιτρέπουν να διέρχεται ρεύμα παθητικά μέσω των διακυτταρικών διαύλων. (Β) Στις χημικές συνάψεις, δεν υπάρχει κυτταροπλασματική συνέχεια και συνεπώς ούτε κατευθυνόμενη ροή ρεύματος από το προ- στο μετασυναπτικό κύτταρο.
 Στις ηλεκτρικές συνάψεις, οι χασμοσύνδεσμοι μεταξύ προ- και μετασυναπτικών μεμβρανών επιτρέπουν να διέρχεται ρεύμα παθητικά μέσω των διακυτταρικών διαύλων. (Β) Στις χημικές συνάψεις, δεν υπάρχει κυτταροπλασματική συνέχεια και συνεπώς ούτε κατευθυνόμενη ροή ρεύματος από το προ- στο μετασυναπτικό κύτταρο..")
15
Ηλεκτρικές συνάψεις Οι ηλεκτρικές συνάψεις εκδηλώνονται με τη μορφή «χασμοσυνδέσμων». Παρατηρούνται σε όλα τα νευρικά συστήματα, συμπεριλαμβανομένου και του ανθρώπινου εγκεφάλου. Επιτρέπουν τη παθητική ροή ιονικού ρεύματος διαμέσου των πόρων του χασμοσυνδέσμου από τον ένα νευρώνα στον άλλον. Ο ένας νευρώνας, ο οποίος είναι και η πηγή του ρεύματος ονομάζεται προσυναπτικός, ενώ ο νευρώνας στον οποίο διέρχεται το ρεύμα ονομάζεται μετασυναπτικός. η κατεύθυνση της ροής του ρεύματος μπορεί να είναι αμφίδρομη.

16
Λειτουργίες χασμοσυνδέσμων μέσω ηλεκτρικών συνάψεων
Χαρακτηριστικό των ηλεκτρικών συνάψεων με τη μορφή χασμοσυνδέσμωνη υπερβολικά γρήγορη μεταφορά των μηνυμάτων διαμέσου αυτών. Τέτοιες συνάψεις διασυνδέουν πολλούς από τους νευρώνες που επιτρέπουν στα οστρακοειδή αλλά και σε έντομα να διαφύγουν από τους θηρευτές τους.

17
Χημικές συνάψεις – Μονόδρομη κυτταρική επικοινωνία
Κύριο χαρακτηριστικό τους η παρουσία μικρών μεμβρανο-περίβλητων οργανιδίωντα συναπτικά κυστίδια που βρίσκονται στο τελικό κομβίο του προσυναπτικού άξονα. Τα σφαιρικά αυτά οργανίδια είναι γεμάτα με νευροδιαβιβαστές χημικά συνιάλα που εκκρίνονται από τον προσυναπτικό νευρώνα. Υπάρχουν πολλών ειδών νευροδιαβιβαστές, η πιο μελετημένη είναι η ακετυλοχολίνη (Ach). Το κενό μεταξύ προσυναπτικών και μετασυναπτικών νευρώνων αρκετά μεγαλύτερο συναπτική σχισμή. Η διαδικασία διαβίβασης μέσω των χημικών συνάψεων ξεκινά όταν ένα δυναμικό ενέργειας εισέρχεται στο τελικό κομβίο του προσυναπτικού νευρώνα
. Το κενό μεταξύ προσυναπτικών και μετασυναπτικών νευρώνων αρκετά μεγαλύτερο συναπτική σχισμή. Η διαδικασία διαβίβασης μέσω των χημικών συνάψεων ξεκινά όταν ένα δυναμικό ενέργειας εισέρχεται στο τελικό κομβίο του προσυναπτικού νευρώνα.")
18
ΕΙΚΟΝΑ 8. ΣΕΙΡΑ ΤΩΝ ΓΕΓΟΝΟΤΩΝ ΠΟΥ ΠΑΙΡΝΟΥΝ ΜΕΡΟΣ ΚΑΤΑ ΤΗ ΔΙΑΒΙΒΑΣΗ ΣΕ ΜΙΑ ΤΥΠΙΚΗ ΧΗΜΙΚΗ ΣΥΝΑΨΗ.

19
ΠΛΑΣΜΟΔΕΣΜΕΣ Τα φυτικά κύτταρα έχουν μόνο ένα είδος διακυτταρικών συνδέσεων, τις πλασμοδέσμες δημιουργούν εξαιρετικά κυτταροπλασματικά κανάλια (διαμέτρου 20 – 40 nm) μέσω των παρεμβαλλόμενων κυτταρικών τοιχωμάτων (πάχος τουλάχιστον 0.1 μm) =>τα κύτταρα ενός φυτού φαίνονται ότι δημιουργούν ένα συγκύτιο στο οποίο πολλοί κυτταρικοί πυρήνες μοιράζονται ένα κοινό κυτταρόπλασμα. Κατά μήκος του κέντρου του καναλιού υπάρχει μια στενή κυλινδρική δομή, ο δεσμοσωλήνας βρίσκεται σε συνέχεια με στοιχεία του αδρού ενδοπλασματικού δικτύου σε κάθε ένα από τα συνδεόεμενα κύτταρα. Η κυτταροπλασματική δομή μεταξύ του δεσμοσωλήνα και της μεμβράνης της πλασμοδέσμης, ονομάζεται δακτύλιος (annulus) διατηρεί την κυτταροπλασματική συνέχεια των γειτονικών κυττάρων, επιτρέποντας στα μόρια να διαπερνούν από το ένα στο άλλο.
 μέσω των παρεμβαλλόμενων κυτταρικών τοιχωμάτων (πάχος τουλάχιστον 0.1 μm) =>τα κύτταρα ενός φυτού φαίνονται ότι δημιουργούν ένα συγκύτιο στο οποίο πολλοί κυτταρικοί πυρήνες μοιράζονται ένα κοινό κυτταρόπλασμα. Κατά μήκος του κέντρου του καναλιού υπάρχει μια στενή κυλινδρική δομή, ο δεσμοσωλήνας βρίσκεται σε συνέχεια με στοιχεία του αδρού ενδοπλασματικού δικτύου σε κάθε ένα από τα συνδεόεμενα κύτταρα. Η κυτταροπλασματική δομή μεταξύ του δεσμοσωλήνα και της μεμβράνης της πλασμοδέσμης, ονομάζεται δακτύλιος (annulus) διατηρεί την κυτταροπλασματική συνέχεια των γειτονικών κυττάρων, επιτρέποντας στα μόρια να διαπερνούν από το ένα στο άλλο.")
20
ΕΙΚΟΝΑ 9. ΠΛΑΣΜΟΔΕΣΜΕΣ.(Α)Τα κυτταροπλασματικά κανάλια των πλασμοδεσμών διαπερνούν το κυτταρικό τοίχωμα του φυτού και συνδέουν όλα τα κύτταρα ενός φυτού μαζί. (Β) Κάθε πλασμοδέσμη επενδύεται εσωτερικά με τη πλασματική μεμβράνη που είναι κοινή στα δύο συνδεόμενα κύτταρα. Συνήθως περιέχει επίσης και μια κυλινδρική δομή, το δεσμοσωλήνα, ο οποίος προέρχεται από το αδρό ενδοπλασματικό δίκτυο.
Τα κυτταροπλασματικά κανάλια των πλασμοδεσμών διαπερνούν το κυτταρικό τοίχωμα του φυτού και συνδέουν όλα τα κύτταρα ενός φυτού μαζί. (Β) Κάθε πλασμοδέσμη επενδύεται εσωτερικά με τη πλασματική μεμβράνη που είναι κοινή στα δύο συνδεόμενα κύτταρα. Συνήθως περιέχει επίσης και μια κυλινδρική δομή, το δεσμοσωλήνα, ο οποίος προέρχεται από το αδρό ενδοπλασματικό δίκτυο..")
21
Οι πλασμοδέσμες δημιουργούνται συγχρόνως με το κυτταρικό τοίχωμα κατά τη κυτταρική διαίρεση και παγιώνεται ο αριθμός και η θέση τους σε αυτή τη φάση, ενώ μικρότερες αλλαγές μπορούν να συμβούν αργότερα. Μπορούν όμως να εισαχθούν και de novo μέσω προϋπάρχοντων κυτταρικών τοιχωμάτων, οι οποίοι συνήθως βρίσκονται σε πυκνές δέσμες, που ονομάζονται pit fields (περιοχές βοθρίων). Όταν δε χρειάζονται πια, μπορούν να απομακρυνθούν αμέσως.
. Όταν δε χρειάζονται πια, μπορούν να απομακρυνθούν αμέσως..")
22
ΕΙΚΟΝΑ 10. ΠΟΙΚΙΛΕΣ ΟΨΕΙΣ ΤΩΝ ΠΛΑΣΜΟΔΕΣΜΩΝ
ΕΙΚΟΝΑ 10. ΠΟΙΚΙΛΕΣ ΟΨΕΙΣ ΤΩΝ ΠΛΑΣΜΟΔΕΣΜΩΝ. (Α) Ηλεκτρονική μικρογραφία επιμήκους τμήματος πλασμοδέσμης μιας φτέρας του νερού. Η πλασματική μεμβράνη επενδύει τον πόρο και είναι συνεχής από το ένα κύτταρο στο άλλο. Διαφαίνεται και το ενδοπλασματικό δίκτυο και η σχέση του με τον δεσμοσωλήνα. (Β) Μια παρόμοια πλασμοδέσμη σε διατομή. (Γ) Μικρά pit fields πλασμοδεσμών σε κυτταρικό τοίχωμα που έχει απομονωθεί από φύλλο καπνού.
 Ηλεκτρονική μικρογραφία επιμήκους τμήματος πλασμοδέσμης μιας φτέρας του νερού. Η πλασματική μεμβράνη επενδύει τον πόρο και είναι συνεχής από το ένα κύτταρο στο άλλο. Διαφαίνεται και το ενδοπλασματικό δίκτυο και η σχέση του με τον δεσμοσωλήνα. (Β) Μια παρόμοια πλασμοδέσμη σε διατομή. (Γ) Μικρά pit fields πλασμοδεσμών σε κυτταρικό τοίχωμα που έχει απομονωθεί από φύλλο καπνού.")
23
Παρά τις βασικές διαφορές στη δομή μεταξύ των πλασμοδεσμών και των χασμοσυνδέσμων, φαίνεται ότι ο τρόπος λειτουργίας τους είναι αρκετά όμοιος με αυτόν των χασμοσυνδέσμων : οι πλασμοδέσμες επιτρέπουν την είσοδο σε μόρια με Μ.Β.< 800 (παρόμοιο με το μέγιστο Μ.Β που επιτρέπουν οι χασμοσύνδεσμοι). η μεταφορά μέσω των πλασμοδεσμών μπορεί να ρυθμιστεί πειράματα με φθορίζοντες ουσίες δείχνουν ότι μπορεί να υπάρχουν εμπόδια στην μετακίνηση ακόμα και στα χαμηλού Μ.Β μόρια μεταξύ κυττάρων που συνδέονται με φαινομενικά φυσιολογικές πλασμοδέσμες. Οι μηχανισμοί που εμποδίζουν την επικοινωνία σ’αυτές τις περιπτώσεις δεν είναι κατανοητοί.
. η μεταφορά μέσω των πλασμοδεσμών μπορεί να ρυθμιστεί πειράματα με φθορίζοντες ουσίες δείχνουν ότι μπορεί να υπάρχουν εμπόδια στην μετακίνηση ακόμα και στα χαμηλού Μ.Β μόρια μεταξύ κυττάρων που συνδέονται με φαινομενικά φυσιολογικές πλασμοδέσμες. Οι μηχανισμοί που εμποδίζουν την επικοινωνία σ’αυτές τις περιπτώσεις δεν είναι κατανοητοί.")
24
Γονίδια που κωδικοποιούν ρυθμιστικές πρωτείνες που συμμετέχουν στη διαδικασία καθορισμού, περνούν από κύτταρο σε κύτταρο μέσω των πλασμοδεσμών, προσδένονται σε στοιχεία αυτών και ξεπερνούν το μηχανισμό που οριοθετεί το μέγεθος των μορίων, που κανονικά θα εμπόδιζε την είσοδό τους. Σε μερικές περιπτώσεις, ακόμη και το mRNA που κωδικοποιεί την πρωτεϊνη μπορεί να περάσει. Μερικοί ιοί φυτών εκμεταλλεύονται αυτή τη διαδικασία => μολυσματικά ιϊκά RNAs, ή ακόμη και κάποια ιϊκά τμήματα, μπορούν να περάσουν από κύτταρο σε κύτταρο μ’αυτό το τρόπο. Αυτοί οι ιοί παράγουν πρωτεϊνες που προσδένονται σε κάποια στοιχεία των πλασμοδεσμών και αυξάνουν δραματικά την αποτελεσματικότητα του μεγέθους του πόρου του καναλιού. Αυτά τα στοιχεία των πλασμοδεσμών παραμένουν άγνωστα => δεν είναι ξεκάθαρο πώς ενδογενή ή ιϊκά μακρομόρια ρυθμίζουν τη δυνατότητα μεταφοράς των καναλιών και διέρχονται μέσω αυτών.

25
Ο ΡΟΛΟΣ ΤΩΝ ΓΟΝΙΔΙΩΝ ΤΩΝ ΚΟΝΝΕΞΙΝΩΝ ΤΩΝ ΧΑΣΜΟΣΥΝΔΕΣΜΩΝ ΣΕ ΔΙΑΦΟΡΕΣ ΑΣΘΕΝΕΙΕΣ
Η διακυτταρική επικοινωνία μέσω των χασμοσυνδέσμων θεωρείται ότι παίζει σημαντικό ρόλο στη διατήρηση της ομοιόστασης στη μορφογένεση και στη κυτταρική ανάπτυξη και διαφοροποίηση σε πολυκύτταρους οργανισμούς.

26
Έλλειψη ή ανωμαλία διαφόρων γονιδίων κοννεξινών οδηγεί σε διάφορες δυσλειτουργίες, όπως :
καρκίνους καρδιακές ασθένειες καταρράκτη πολυνευροπάθειες κώφωση δερματικές παθήσεις και άλλες

27
Τα γονίδια των κοννεξινών μπορούν και ασκούν διπλή επίδραση στον έλεγχο των όγκων :
καταστολή όγκων Μεταφορά γονιδίων των κοννεξινών σε καρκινογενή κύτταρα επαναφέρει τη φυσιολογική κυτταρική ανάπτυξη => κοννεξίνες σχηματίζουν μια οικογένεια ογκοκατασταλτικών γονιδίων. Bystander effect (παρατυχούσα επίδραση) για τη θεραπεία του καρκίνου Στη γονιδιακή θεραπεία του HSV-tk / glanciclovir.
 για τη θεραπεία του καρκίνου Στη γονιδιακή θεραπεία του HSV-tk / glanciclovir.")
28
Μεταλλαγές στα γονίδια των κοννεξινών προκαλούν δερματικές παθήσεις, αλλά και κώφωση.
Έγιναν μελέτες που στόχευαν στο σύνδρομο KID (Keratitis-Ichthyosis-Deafness) μια σπάνια εκτοδερμική δυσπλασία που χαρακτηρίζεται από επιδερμική κερατίτιδα και αγγείωση του κερατοειδή χιτώνα, που πιθανώς οδηγεί σε τύφλωση, βαθιά νευροαισθητήρια απώλεια ακοής και πάχυνση του δέρματος επηρεάζει πολλούς επιθηλιακούς ιστούς και πιθανόν αντικατοπτρίζει διαταραχές στην διαδικασία της κυτταρικής διαφοροποίησης και ωρίμανσης. στον ρόλο των χασμοσυνδέσμων στην σηματοδότηση κυττάρων στους προσβαλλόμενους ιστούς, στην επιδερμίδα, στον κερατοειδή χιτώνα και το έσω αυτί.
 μια σπάνια εκτοδερμική δυσπλασία που χαρακτηρίζεται από επιδερμική κερατίτιδα και αγγείωση του κερατοειδή χιτώνα, που πιθανώς οδηγεί σε τύφλωση, βαθιά νευροαισθητήρια απώλεια ακοής και πάχυνση του δέρματος επηρεάζει πολλούς επιθηλιακούς ιστούς και πιθανόν αντικατοπτρίζει διαταραχές στην διαδικασία της κυτταρικής διαφοροποίησης και ωρίμανσης. στον ρόλο των χασμοσυνδέσμων στην σηματοδότηση κυττάρων στους προσβαλλόμενους ιστούς, στην επιδερμίδα, στον κερατοειδή χιτώνα και το έσω αυτί.")
29
Οι ερευνητές μελέτησαν μερικά διαφορετικά γονίδια κοννεξινών που εκφράζονται στους προσβαλλόμενους ιστούς-στόχους στο σύνδρομο KID και επιτυχώς ταυτοποίησαν μεταλλαγές που προκαλούν δυσλειτουργίες, σ’ένα γονίδιο που καλείται GJB2 το οποίο κωδικοποιεί την χασμοσυνδεσμοτική πρωτεϊνη Cx26, σε όλους τους ασθενείς. Αποτελέσματα : τα περισσότερα άτομα με το σύνδρομο KID κουβαλάνε μια νέα μεταλλαγή, η οποία μπορεί να μεταφερθεί στους απόγονους του προσβαλλόμενου ατόμου με αυτοσωμικό επικρατή τρόπο. Τώρα γίνονται έρευνες για μοριακά διαγνωστικά τέστ και προγεννετική διάγνωση του συνδρόμου αυτού.

30
ΦΡΑΓΜΟΣΥΝΔΕΣΜΟΙ Λειτουργία: μεσοκυττάριος αποκλεισμός στήριξη
Φραγμοί μεταξύ κυττάρων Αύξηση αυξητικών παραγόντων απορρύθμιση φραγμών ασθένειες πχ. αγγειακός ενδοθηλιακός αυξητικός παράγοντας (VEGF)
")
31
Στενοσύνδεσμοι (σπονδυλόζωα) Διαφραγματοσύνδεσμοι (ασπόνδυλα)
ΦΡΑΓΜΟΣΥΝΔΕΣΜΟΙ Στενοσύνδεσμοι (σπονδυλόζωα) Διαφραγματοσύνδεσμοι (ασπόνδυλα)
 Διαφραγματοσύνδεσμοι (ασπόνδυλα)")
32
ΣΤΕΝΟΣΥΝΔΕΣΜΟΙ Περιορίζουν τις πρωτεΐνες μεταφοράς στις κατάλληλες μεμβρανικές περιοχές Εμποδίζουν τη διάχυση των μορίων πίσω στον αυλό

33
ΣΤΕΝΟΣΥΝΔΕΣΜΟΙ

34
Διαμεμβρανικές πρωτεΐνες προσκόλλησης
ΣΤΕΝΟΣΥΝΔΕΣΜΟΙ Διαμεμβρανικές πρωτεΐνες προσκόλλησης Οκλουδίνη Κλωδίνη JAM (junctional adhesion molecule) Αρκετές άλλες
 Αρκετές άλλες.")
35
ΣΤΕΝΟΣΥΝΔΕΣΜΟΙ

36
ΔΙΑΦΡΑΓΜΑΤΟΣΥΝΔΕΣΜΟΙ

37
ΣΥΝΔΕΣΜΟΙ ΠΡΟΣΔΕΣΗΣ Αποτελούνται από: Κυτταρικές πρωτεΐνες προσδεσης Διαμεμβρανικές πρωτεΐνες προσκόλλησης

38
ΣΥΝΔΕΣΜΟΙ ΠΡΟΣΔΕΣΗΣ (α) Σύνδεσμοι προσκόλλησης Δεσμοσώματα
(περιέχουν μόρια καδερίνης, συνδέουν δύο κύτταρα μεταξύ τους) (β) Εστίες προσκόλλησης Ημιδεσμοσώματα (περιέχουν μόρια ιντεγκρίνης, συνδέουν το κύτταρο με μόρια εξωκυττάριας ουσίας)
 (β) Εστίες προσκόλλησης. Ημιδεσμοσώματα. (περιέχουν μόρια ιντεγκρίνης, συνδέουν το κύτταρο με μόρια εξωκυττάριας ουσίας)")
39
ΣΥΝΔΕΣΜΟΙ ΠΡΟΣΚΟΛΛΗΣΗΣ

40
ΣΥΝΔΕΣΜΟΙ ΠΡΟΣΚΟΛΛΗΣΗΣ
Οι καδερίνες παίζουν ρόλο στη μορφογένεση και πιθανόν και στην οστεογένεση

41
ΔΕΣΜΟΣΩΜΑΤΑ Καδερίνες (δεσμογλεΐνη, δεσμοκολλίνη)
Armadillo (πλακοσφαιρίνη, πλακοφιλίνη) Πλακίνες (δεσμοπλακίνες, πλεκτίνες)
 Πλακίνες (δεσμοπλακίνες, πλεκτίνες)")
42
Παρέχουν μηχανική σταθερότητα σε ιστούς (πχ. καρδιά, επιδερμίδα)
ΔΕΣΜΟΣΩΜΑΤΑ Παρέχουν μηχανική σταθερότητα σε ιστούς (πχ. καρδιά, επιδερμίδα) Δέχονται ρυθμιστικά σινιάλα Συμμετέχουν στη μορφογένεση των ιστών και τη διαφοροποίηση
 Δέχονται ρυθμιστικά σινιάλα. Συμμετέχουν στη μορφογένεση των ιστών και τη διαφοροποίηση.")
43
ΕΣΤΙΕΣ ΠΡΟΣΚΟΛΛΗΣΗΣ

44
ΗΜΙΔΕΣΜΟΣΩΜΑΤΑ

46
ΚΥΤΤΑΡΙΚΗ ΠΡΟΣΚΟΛΛΗΣΗ

47
ΞΑΝΑΣΧΗΜΑΤΙΣΜΟΣ ΛΕΙΤΟΥΡΓΙΚΟΥ ΙΣΤΟΥ ΥΣΤΕΡΑ ΑΠΟ ΔΙΑΧΩΡΙΣΜΟ ΚΥΤΤΑΡΩΝ ΣΕ ΣΠΟΝΔΥΛΩΤΟ.
ΥΣΤΕΡΑ ΑΠΌ ΚΑΤΕΡΓΑΣΙΑ ΕΜΒΡΥΪΚΟΥ ΙΣΤΟΥ ΜΕ ΠΡΩΤΕΟΛΥΤΙΚΑ ΕΝΖΥΜΑ(ΘΡΥΨΙΝΗ), ΑΦΑΙΡΕΣΗ ΙΟΝΤΩΝ ΑΣΒΕΣΤΙΟΥ ΚΑΙ ΜΑΓΝΗΣΙΟΥ ΚΑΙ ΠΡΟΣΘΗΚΗ EDTA ΤΑ ΔΙΑΧΩΡΙΣΜΕΝΑ ΚΥΤΤΑΡΑ ΣΕ IN VITRO ΔΟΚΙΜΕΣ ΞΑΝΑΣΥΓΚΕΝΤΡΩΝΟΝΤΑΙ ΣΕ ΔΟΜΕΣ ΠΟΥ ΜΟΙΑΖΟΥΝ ΜΕ ΤΟΝ ΑΡΧΙΚΟ ΙΣΤΟ. Γενικότερα τα διαχωρισμένα κύτταρα βρίσκουν να προσκολληθούν ευκολότερα σε συσσωματώματα του ίδιου τους του οργάνου παρά σε συσσωματώματα διαφορετικών οργάνων. Άρα υπάρχει ένα σύστημα αναγνώρισης το οποίο προτρέπει κύτταρα του ίδιου διαφοροποιημένου ιστού να προτιμήσει την προσκόλληση σε ένα άλλο κύτταρο του ίδιου ιστού. Αυτή η προτίμηση που υπάρχει στην προσκόλληση είναι πιθανώς σημαντική στην σταθεροποίηση της δομής του ιστού .
, ΑΦΑΙΡΕΣΗ ΙΟΝΤΩΝ ΑΣΒΕΣΤΙΟΥ ΚΑΙ ΜΑΓΝΗΣΙΟΥ ΚΑΙ ΠΡΟΣΘΗΚΗ EDTA ΤΑ ΔΙΑΧΩΡΙΣΜΕΝΑ ΚΥΤΤΑΡΑ ΣΕ IN VITRO ΔΟΚΙΜΕΣ ΞΑΝΑΣΥΓΚΕΝΤΡΩΝΟΝΤΑΙ ΣΕ ΔΟΜΕΣ ΠΟΥ ΜΟΙΑΖΟΥΝ ΜΕ ΤΟΝ ΑΡΧΙΚΟ ΙΣΤΟ. Γενικότερα τα διαχωρισμένα κύτταρα βρίσκουν να προσκολληθούν ευκολότερα σε συσσωματώματα του ίδιου τους του οργάνου παρά σε συσσωματώματα διαφορετικών οργάνων. Άρα υπάρχει ένα σύστημα αναγνώρισης το οποίο προτρέπει κύτταρα του ίδιου διαφοροποιημένου ιστού να προτιμήσει την προσκόλληση σε ένα άλλο κύτταρο του ίδιου ιστού. Αυτή η προτίμηση που υπάρχει στην προσκόλληση είναι πιθανώς σημαντική στην σταθεροποίηση της δομής του ιστού .")
48
Τα κύτταρα προσκολλούνται το ένα στο άλλο και στο εξωκυττάριο στρώμα δια μέσου πρωτεϊνών κυτταρικής επιφανείας οι οποίες λέγονται μόρια κυτταρικής προσκόλλησης (CAM). Μερικά μόρια κυτταρικής προσκόλλησης εξαρτώνται από ιόντα ασβεστίου ενώ κάποια άλλα είναι ανεξάρτητα από τα ιόντα ασβεστίου. Όσα μόρια κυτταρικής προσκόλλησης εξαρτώνται από ιόντα ασβεστίου είναι κυρίως υπεύθυνα για την ειδική για τον ιστό προσκόλληση κύτταρο με κύτταρο στα έμβρυα σπονδυλωτών εξηγώντας το γιατί αυτά τα κύτταρα μπορούν να αποσυναθροιστούν παρουσία ιόντων ασβεστίου και χειλικού παράγοντα. Αρχικά τα μόρια κυτταρικής προσκόλλησης αναγνωρίστηκαν φτιάχνοντας αντισώματα έναντι των μορίων κυτταρικής επιφανείας και μετά δοκίμασαν τα αντισώματα ως προς την ικανότητα τους να αναστέλλουν την κύτταρο με κύτταρο προσκόλληση στον δοκιμαστικό σωλήνα. Αυτά λοιπόν τα αντισώματα τα οποία αναστέλλουν την προσκόλληση χρησιμοποιήθηκαν έπειτα για να χαρακτηριστούν τα μόρια προσκόλλησης και να απομονωθούν.

49
ΚΑΤΗΓΟΡΙΕΣ ΠΡΩΤΕΪΝΩΝ ΚΥΤΤΑΡΙΚΗΣ ΠΡΟΣΚΟΛΛΗΣΗΣ
ΚΑΔΕΡΙΝΕΣ ΣΕΛΕΚΤΙΝΕΣ ΥΠΕΡΟΙΚΟΓΕΝΕΙΑ ΑΝΟΣΟΣΦΑΙΡΙΝΩΝ

50
ΤΑ ΚΥΡΙΑ ΜΟΡΙΑ ΚΥΤΤΑΡΙΚΗΣ ΠΡΟΣΚΟΛΛΗΣΗΣ
ΟΙ ΚΑΔΕΡΙΝΕΣ ΕΞΑΡΤΩΝΤΑΙ ΑΠΟ ΤΗΝ ΔΙΑΜΕΣΟΛΑΒΗΣΗ ΤΩΝ ΙΟΝΤΩΝ ΑΣΒΕΣΤΙΟΥ ΓΙΑ ΤΗΝ ΚΥΤΤΑΡΙΚΗ ΠΡΟΣΚΟΛΛΗΣΗ ΤΑ ΚΥΡΙΑ ΜΟΡΙΑ ΚΥΤΤΑΡΙΚΗΣ ΠΡΟΣΚΟΛΛΗΣΗΣ ΧΩΡΙΖΟΝΤΑΙ ΣΕ ΚΛΑΣΙΚΕΣ ΚΑΙ ΜΗ ΚΛΑΣΙΚΕΣ ΚΑΔΕΡΙΝΕΣ ΕΚΦΡΑΖΟΝΤΑΙ ΣΕ ΣΠΟΝΔΥΛΩΤΑ ΚΑΙ ΑΣΠΟΝΔΥΛΑ ΟΙ ΠΕΡΙΣΣΟΤΕΡΕΣ ΚΑΔΕΡΙΕΝΕΣ ΕΙΝΑΙ ΜΟΝΗΣ ΔΙΕΛΕΥΣΗΣ ΜΗΚΟΥΣ ΑΜΙΝΟΞΕΑ ΣΧΗΜΑΤΙΖΟΥΝ ΔΙΜΕΡΕΣ Ή ΜΕΓΑΛΥΤΕΡΟ ΟΛΙΓΟΜΕΡΕΣ ΤΟ ΕΞΩΚΥΤΤΑΡΙΟ ΚΟΜΜΑΤΙ ΤΗΣ ΠΟΛΥΠΕΠΤΙΔΙΚΗΣ ΑΛΥΣΙΔΑΣ ΔΙΠΛΩΝΕΤΑΙ ΣΕ 5 Ή 6 ΚΑΔΕΡΙΝΙΚΕΣ ΕΠΑΝΑΛΛΗΨΕΙΣ (ΔΟΜΙΚΑ ΣΧΕΤΙΖΟΝΤΑΙ ΜΕ ΤΙΣ ΠΕΡΙΟΧΕΣ ΤΩΝ ΑΝΟΣΟΣΦΑΙΡΙΝΩΝ)
")
51
ΡΟΛΟΣ ΤΩΝ ΙΟΝΤΩΝ ΑΣΒΕΣΤΙΟΥ ΣΤΗΝ ΛΕΙΤΟΥΡΓΙΑ ΤΩΝ ΚΑΔΕΡΙΝΩΝ

52
ΟΙ ΚΑΔΕΡΙΝΕΣ ΔΙΑΜΕΣΟΛΑΒΟΥΝ ΓΙΑ ΤΗΝ ΚΥΤΤΑΡΟ ΜΕ ΚΥΤΤΑΡΟ ΠΡΟΣΚΟΛΛΗΣΗ ΜΕ ΕΝΑΝ ΟΜΟΦΙΛΙΚΟ ΜΗΧΑΝΙΣΜΟ
ΟΜΟΦΙΛΙΚΗ:ΜΟΡΙΑ ΤΟΥ ΕΝΟΣ ΚΥΤΤΑΡΟΥ ΣΥΝΔΕΟΝΤΑΙ ΜΕ ΜΟΡΙΑ ΤΟΥ ΙΔΙΟΥ ΤΥΠΟΥ ΣΕ ΓΕΙΤΟΝΙΚΑ ΚΥΤΤΑΡΑ ΕΤΕΡΟΦΙΛΙΚΑ:ΜΟΡΙΑ ΤΟΥ ΕΝΌΣ ΚΥΤΤΑΡΟΥ ΔΕΣΜΕΥΟΝΤΑΙ ΜΕ ΜΟΡΙΑ ΔΙΑΦΟΡΕΤΙΚΟΥ ΤΥΠΟΥ ΣΕ ΔΙΠΛΑΝΑ ΚΥΤΤΑΡΑ ΤΑ ΓΕΙΤΟΝΙΚΑ ΚΥΤΤΑΡΑ ΣΥΝΔΕΟΝΤΑΙ ΧΑΡΗ ΣΕ ΈΝΑ ΕΚΡΡΙΝΟΜΕΝΟ ΜΟΡΙΟ ΣΥΝΔΕΤΗ

53
ΔΙΑΦΟΡΕΤΙΚΕΣ ΚΑΔΕΡΙΝΕΣ ΠΡΟΤΙΜΟΥΝ ΝΑ ΔΕΣΜΕΥΟΝΤΑΙ ΣΤΟΝ ΙΔΙΟ ΤΥΠΟ ΠΟΥ ΑΝΗΚΟΥΝ

54
ΟΙ ΚΑΔΕΡΙΝΕΣ ΣΥΝΔΕΟΝΤΑΙ ΣΤΟ ΚΥΤΤΑΡΟΣΚΕΛΕΤΟ ΑΚΤΙΝΗΣ ΜΕ ΚΑΤΕΝΙΝΕΣ.

55
Μερικά κύτταρα μπορούν να ρυθμίζουν την ενεργότητα προσκόλλησης των καδερινών τους.

56
ΟΙ ΣΕΛΕΚΤΙΝΕΣ ΜΕΣΟΛΑΒΟΥΝ ΓΙΑ ΤΗΝ ΠΑΡΟΔΙΚΗ ΚΥΤΤΑΡΟ ΜΕ ΚΥΤΤΑΡΟ ΠΡΟΣΚΟΛΛΗΣΗ ΣΤΗ ΡΟΗ ΤΟΥ ΑΙΜΑΤΟΣ
L-ΣΕΛΕΚΤΙΝΕΣ:ΛΕΥΚΑ ΑΙΜΟΣΦΑΙΡΙΑ P-ΣΕΛΕΚΤΙΝΕΣ:ΑΙΜΟΠΕΤΑΛΙΑ ΚΑΙ ΕΝΔΟΘΗΛΙΑΚΑ ΚΥΤΤΑΡΑ Ε-ΣΕΛΕΚΤΙΝΕΣ:ΕΝΕΡΓΟΠΟΙΗΜΕΝΑ ΚΥΤΤΑΡΑ ΕΝΔΟΘΗΛΙΟΥ ΕΞΑΡΤΩΝΤΑΙ ΑΠΌ ΙΟΝΤΑ ΑΣΒΕΣΤΙΟΥ ΣΥΝΕΡΓΑΖΟΝΤΑΙ ΜΕ ΤΙΣ ΙΝΤΕΓΚΡΙΝΕΣ

57
ΤΑ ΙΟΝΤΑ ΑΣΒΕΣΤΙΟΥ ΔΕΝ ΔΙΑΜΕΣΟΛΑΒΟΥΝ ΣΤΗΝ ΥΠΕΡΟΙΚΟΓΕΝΕΙΑ ΤΩΝ ΑΝΟΣΟΣΦΑΙΡΙΝΩΝ
ΠΕΡΙΕΧΟΥΝ ΜΙΑ Ή ΠΕΡΙΣΣΟΤΕΡΕΣ ΠΕΡΙΟΧΕΣ ΧΑΡΑΚΤΗΡΙΣΤΙΚΕΣ ΓΙΑ ΤΑ ΑΝΤΙΣΩΜΑΤΑ ΟΜΟΦΙΛΙΚΟ ΜΗΧΑΝΙΣΜΟ(ΜΕΡΙΚΕΣ ΕΤΕΡΟΦΙΛΊΚΟ) 5 ΠΕΡΙΟΧΕΣ ΤΟΥ ΕΞΩΚΥΤΤΑΡΙΟΥ ΜΕΡΟΥΣ Ν-CAM ΠΕΡΙΕΧΟΥΝ ΜΕΓΑΛΕΣ ΠΟΣΟΤΗΤΕΣ ΣΙΑΛΙΚΟΥ ΟΞΕΟΣ ΓΙΑ ΠΑΡΑΚΩΛΗΣΕΙ ΤΗΣ ΣΥΝΔΕΣΗΣ ΣΙΝΙΑΛΑ ΣΤΟ ΕΣΩΤΕΡΙΚΟ ΤΟΥ ΚΥΤΤΑΡΟΥ
 5 ΠΕΡΙΟΧΕΣ ΤΟΥ ΕΞΩΚΥΤΤΑΡΙΟΥ ΜΕΡΟΥΣ. Ν-CAM ΠΕΡΙΕΧΟΥΝ ΜΕΓΑΛΕΣ ΠΟΣΟΤΗΤΕΣ ΣΙΑΛΙΚΟΥ ΟΞΕΟΣ ΓΙΑ ΠΑΡΑΚΩΛΗΣΕΙ ΤΗΣ ΣΥΝΔΕΣΗΣ. ΣΙΝΙΑΛΑ ΣΤΟ ΕΣΩΤΕΡΙΚΟ ΤΟΥ ΚΥΤΤΑΡΟΥ.")
58
ΧΗΜΕΙΟΤΑΚΤΙΣΜΟΣ ΜΑΣΤΙΓΙΟ ΕΛΑΣΤΙΚΟ ΕΛΑΤΗΡΙΟ ΠΡΩΤΕΪΝΙΚΟ ΔΙΣΚΟ
ΚΙΝΗΤΗΡΙΟΣ ΜΗΧΑΝΙΣΜΟΣ ΕΝΕΡΓΕΙΑ ΑΠΌ ΔΙΑΜΕΜΒΡΑΝΙΚΗ ΚΛΙΣΗ ΙΟΝΤΩΝ ΥΔΡΟΓΟΜΟΥ

59
ΑΡΙΣΤΕΡΟΣΤΡΟΦΗ ΠΕΡΙΣΤΡΟΦΗ:ΟΜΟΙΟΜΟΡΦΑ ΠΡΟΣ ΜΙΑ ΚΑΤΕΥΘΥΝΣΗ ΔΕΞΙΟΣΤΡΟΦΗ ΠΕΡΙΣΤΡΟΦΗ:ΣΤΡΙΦΟΓΥΡΙΖΟΥΝ

60
ΥΠΟΔΟΧΕΙΣ ΧΗΜΕΙΟΤΑΚΤΙΣΜΟΥ:ΔΙΜΕΡΙΣ ΔΙΑΜΕΜΒΡΑΝΙΚΕΣ ΠΡΩΤΕΪΝΕΣ
CheA:ΚΙΝΑΣΗ ΙΣΤΙΔΙΝΗΣ CheW:ΠΡΩΤΕΪΝΗ ΠΡΟΣΑΡΜΟΓΕΑΣ CheY:ΠΡΩΤΕΪΝΗ ΑΓΓΕΛΙΟΦΟΡΟΣ ΚΑΙ ΠΡΟΚΑΛΕΙ ΤΗΝ ΔΕΞΙΟΣΤΡΟΦΗ ΠΕΡΙΣΤΡΟΦΗ CheZ:ΒΟΗΘΑΕΙ ΣΤΗΝ ΑΠΟΦΩΣΦΟΡΥΛΙΩΣΕΙ ΤΗΣ CheY

61
ΧΗΜΕΙΟΤΑΚΤΙΣΜΟΣ ΣΤΟ Dictyostelium
ΌΤΑΝ ΕΙΣΕΛΘΕΙ ΣΤΗΝ ΦΑΣΗ ΤΗΣ ΠΕΙΝΑΣ ΕΧΟΥΜΕ ΤΗΝ ΣΥΣΣΩΜΑΤΩΣΗ ΠΡΟΣ ΕΝΑ ΚΕΝΤΡΟ ΣΥΣΣΩΜΑΤΩΣΗΣ ΧΗΜΕΙΟΤΑΞΗ ΠΡΟΚΑΛΕΙΤΑΙ ΑΠΌ ΤΟ cAMP

Παρόμοιες παρουσιάσεις