Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Углевод-белковых комплексов
Химическая технология Углевод-белковых комплексов Краткий конспект лекций

2
Лекция№ 1. Биохимияның негізгі қалыптасуы кезеңдері. Aқуыздар.
Лекция жоспары. 1 Биохимияның негізгі қалыптасуы кезеңдері. 2 Aқуыздар, жіктелуі. 3 Aқуыздардыңод химиялық құрлысы, амин қышқылдарының жіктелуі. Билогиялық химия тіршілік туралы ғылым, яғни тіршлік ету процестерінде заттардың құрамын,энергия алмасуын және олардың бір бірімен тығыз байланыста екендігін молекулалық денгейде зертейтін ғылым. Биохимия XIX-ғасырдың аяғынада ғана жақын физиология мен органикалық химия ғылымдарының жеке ғылым ретінде болініп шықты. XIX-ғасырындағы аяғында биохимия медицина мен аул шаруашылығында көмегін тигізе отырып, тез дами бастады. Биохимия ғылымның дамуына Д.И. Менделеев, А.М. Бутлеров, И.И. Мечников, И.М. Сеченов, А.Я. Данилевский, И.П. Павлов, К.А. Темирязев, М.В. Ненцкий, В.И. Палладин секілді орыс ғалымдары оздерінің зор үлестерін қосты. , осы ғылымның дамуына Э. Фишер, Ф. Сенджер, Д. Уотсен, Ф. Крик, С.Очоа, Совет одағы кезінде биохимияның негізін салушы А.Я. Данилевский ақуыздардың құрамын, құрлысын, қасиеттерін, ферменттердің қасиеттерін, антиферменттерді алу жолдарын зерттеді. Орыс физиолгиясының негізін салушы И.М.Сеченов- Тыныс алу процесінің физиологиясы мен химиясын толық зерттеді. Қазақстанда медициналық биохимия саласында елеулі еңбек еткен ғалымдар Б.И. Ильин-Кукуев, В.П. Верболович, Т.Ш.Шарманов. Ал өсімдіктер биохимиясын дамыту барысында Т. Б. Дарханбаев, Н.А. Айтхожина, Р.М. Кунаева сияқты ғалымдардың қосқан үлестері зор. Сонымен қатарА. Корнберг, Ф. Липман, Г. Кребс сияқты шетел ғалымдарының қосқан үлестері айтарлықтай болды. Биохимия пәнің, оның міндеттеріне сәйкес жалпы биохимия, өсімдіктер биохимиясы, адам мен жануарлар биохимиясы, микроағзалар биохимисы медициналық биохимия деп бөлуге болады. Ф.Энгельс өзінің «Табиғат диалектикасы» еңбегінде: «Өмір дегеніміз- ақуызды денелердің өмір сүруі тәсілі яғни ағзадағы зат алмасудың қоршаған

3
ортасымен тығыз байланысы»- деп жазды
ортасымен тығыз байланысы»- деп жазды. Басқаша айтқанда ақуыз- барлық тірінің материалдық негізі болса ал олардын тіршілік ету тәсілі-өмір болып табылады. Ақуыздардын «ақуыз деп аталуы,олардын тауқ жұмартқысының қайнап, денатурацияланған кезде- ақ, тығыз массаға айналуынан шыққан. Оның басқаша атты протейін (protos-бірінші маңызды деген латын сөзінен алынған). Бұл атты Голландия ғалымы Ж.Мульдер берген. Ақуыз- күрделі заттардың жай,қарапайым заттарға дейін тотығуыны қатысатын және жай заттардан күрделі заттардың түзілуіне қатысатын ферменттер. Ақуыз-адам ағзасын микробтардан, ауру туғызушы агенттерден сақтайтын антидене. Актин, миозин сияқты ақуыздар бірімен-бірі АТФ-тың қатысуымен әрекеттесіп, бұлшықеттердің жиырылуы сияқты қызмет атқарды. Ақуыздардың элементтік құрамы: С-50-55% Н-15-18% О-21-24% Н-6,0-7,5% С-0,3-2,5% Бұлардан басқа Р (2%-ке дейін) Сu, Mn, Fe, Со және т.б. элементтер кездеседі. Ақуыздардын молекулалық салмағы мыңнан (жай ақуыздарда) бірнеше милионға дейін (күрделі ақуыздарда) жетеді. Элементтік қурамының қарапайымдылығына қарамастан, (барлығы бес элемент С,О,Н,N,S) ақуыздар өте күрделі полимерлік қосылыстар. Ақуыз макромолекулалары қарапайым амин қышқылдарынан тұрады. Бұл ең алғаш 1820 ж. А.Браконнонын жұмыстарынан көріне бастады. Бұл аминқышқылдарының ең қарапайым өкілі. Н Н2N-C-CООН жылдары Э. Фишер ақуыздардын қышқылдық (25% Н2 СО4 немесе 30% НСl). Қатысуымен cағат ішінде қалдырғанда гидролизге
. Бұл атты Голландия ғалымы Ж.Мульдер берген. Ақуыз- күрделі заттардың жай,қарапайым заттарға дейін тотығуыны қатысатын және жай заттардан күрделі заттардың түзілуіне қатысатын ферменттер. Ақуыз-адам ағзасын микробтардан, ауру туғызушы агенттерден сақтайтын антидене. Актин, миозин сияқты ақуыздар бірімен-бірі АТФ-тың қатысуымен әрекеттесіп, бұлшықеттердің жиырылуы сияқты қызмет атқарды. Ақуыздардың элементтік құрамы: С-50-55% Н-15-18% О-21-24% Н-6,0-7,5% С-0,3-2,5% Бұлардан басқа Р (2%-ке дейін) Сu, Mn, Fe, Со және т.б. элементтер кездеседі. Ақуыздардын молекулалық салмағы мыңнан (жай ақуыздарда) бірнеше милионға дейін (күрделі ақуыздарда) жетеді. Элементтік қурамының қарапайымдылығына қарамастан, (барлығы бес элемент С,О,Н,N,S) ақуыздар өте күрделі полимерлік қосылыстар. Ақуыз макромолекулалары қарапайым амин қышқылдарынан тұрады. Бұл ең алғаш 1820 ж. А.Браконнонын жұмыстарынан көріне бастады. Бұл аминқышқылдарының ең қарапайым өкілі. Н. Н2N-C-CООН жылдары Э. Фишер ақуыздардын қышқылдық (25% Н2 СО4 немесе 30% НСl). Қатысуымен cағат ішінде қалдырғанда гидролизге.")
4
түсетіндігін ашты, міне осы уақыттан бастап ақуыздын химиялық құрамын анықтау барысында жнйелі жұмастар жасала бастады. Аминқышқылдары – ақуыз молекулалаларының кұрамды бөлігі. Олар органикалық қышқылдардың аминді туындылары болып табылады мысалы: СН3- СООН СН3-СН-СООН СН3-(СН2)-СООН Сірке қышқылы пропан қышқылы май қышқылы Бұл қышқылдардын құрамындағы – СООН тобымен байланысқан α- көміртегіндегі сутегі атомы NH2- тобымен байланысқан. Сонымен адам ағзасындағы барлық ақуыз құрамындағы амин қышқылдары α,L- амин қышқылдары болып табылады. Ақуыз құрамында барлығы 20-ға жуық амин қышқылдары кездеседі. α – амин қышқылдары R H-C-COOH NH2 Амин қышқылдарының бірнеше жіктелу жолы бар. Аминқышқылдарының құрамы, атқаратын қызметі бойынша жіктеледі. Амин қышқылдық құрамы бойынша төмендегідей топтарға бөлуге болады: А) алмасуға жатпайтын қышқылдары олар басқа заттардан ағзада синтезделе алмайды, сондыктан да, тамақпен бірге түсуі қажет. Бұл амин қышқылдарына валин, лейцин, изолейцин (алифаттық амин қышқылдары); треонин, лизин, метионин (алифаттық орын басушы топтары бар); фенилаланин (ароматты); триптофан (гетероцклді). Б) жартылай алмасуға жататын амин қышқылдары. Олар ағзада синтезделеді, бірақ жеткіліксіз тнрде болғандықтан, булрдын да тамақпен түсуі қажет (аргинин,тирозин,гистидин). В) алмасуға жататын амин қышқылдары. Ағзада басқа заттардан жеткілікті түрде синтезделеді (аланин, аспарагин қышқылы, глутамин қышқылы, аспарагин, глутамин, серін, глицин).
-СООН. Сірке қышқылы пропан қышқылы май қышқылы. Бұл қышқылдардын құрамындағы – СООН тобымен байланысқан α- көміртегіндегі сутегі атомы NH2- тобымен байланысқан. Сонымен адам ағзасындағы барлық ақуыз құрамындағы амин қышқылдары α,L- амин қышқылдары болып табылады. Ақуыз құрамында барлығы 20-ға жуық амин қышқылдары кездеседі. α – амин қышқылдары R. H-C-COOH. NH2. Амин қышқылдарының бірнеше жіктелу жолы бар. Аминқышқылдарының құрамы, атқаратын қызметі бойынша жіктеледі. Амин қышқылдық құрамы бойынша төмендегідей топтарға бөлуге болады: А) алмасуға жатпайтын қышқылдары олар басқа заттардан ағзада синтезделе алмайды, сондыктан да, тамақпен бірге түсуі қажет. Бұл амин қышқылдарына валин, лейцин, изолейцин (алифаттық амин қышқылдары); треонин, лизин, метионин (алифаттық орын басушы топтары бар); фенилаланин (ароматты); триптофан (гетероцклді). Б) жартылай алмасуға жататын амин қышқылдары. Олар ағзада синтезделеді, бірақ жеткіліксіз тнрде болғандықтан, булрдын да тамақпен түсуі қажет (аргинин,тирозин,гистидин). В) алмасуға жататын амин қышқылдары. Ағзада басқа заттардан жеткілікті түрде синтезделеді (аланин, аспарагин қышқылы, глутамин қышқылы, аспарагин, глутамин, серін, глицин).")
5
Ферменттердін 6 классы бар
Лекция №2 Тірі ағзадағы химиялық реакциялардың жүруіне қатысатын ерекше ақуыздарды фермент немесе энзим деп атайды. Ферменттердің қасиетін құрлысын қызметін зерттейтін ғылымды энзимрлогия деп атайды. Ферменттер барлық мүшелерде, клеткаларда, субклеткалық құрылымдарда кездеседі. Кейбір ферменттер клетка ішінде синтезделіп, өзінің әсерін клеткадан тыс жүргізеді. Мысалы, пепсин,трипсин, липаза сияқты ас қорыту ферменттері. Олар асқазанның кілегейлі қабаттарында синтизделеді. Ферменттердін 6 классы бар I) Оксидоредуктаза классы тотығу – тотықсыдану реакцияларына қатысатын ферменттер. Олардың маңызды өкілдері: Аэробты дегидрогеназалар формула 2) Анаэробты дегидрогеназалар Формула 3) Цитрохромдар
 Оксидоредуктаза классы тотығу – тотықсыдану реакцияларына қатысатын ферменттер. Олардың маңызды өкілдері: Аэробты дегидрогеназалар. формула. 2) Анаэробты дегидрогеназалар. Формула. 3) Цитрохромдар.")
6
4) Пероксидаза және каталаза ферменттері
II) Трансфераза – бір субстраттан екінші субстратка әр түрлі химиялық топтарды тасымалдаушы ферменттер: метилтрансфериза – метил тобын, ацилтрансфераза-ацил тобын, аминотрансфераза- амин тобын, фосфотрансфераза- фосфат тобын тасымалдайды. Мысалы: III) Гидролаза- судын қатысуымен гидролиз реакцияларын жүргізуге қатысатын ферменнтер. Мысалы, эстераза, гликозидаза, пептидаза, фосфотаза және т.б. ферменттер тобы жатады.
 Трансфераза – бір субстраттан екінші субстратка әр түрлі химиялық топтарды тасымалдаушы ферменттер: метилтрансфериза – метил тобын, ацилтрансфераза-ацил тобын, аминотрансфераза- амин тобын, фосфотрансфераза- фосфат тобын тасымалдайды. Мысалы: III) Гидролаза- судын қатысуымен гидролиз реакцияларын жүргізуге қатысатын ферменнтер. Мысалы, эстераза, гликозидаза, пептидаза, фосфотаза және т.б. ферменттер тобы жатады.")
7
4) Пероксидаза және каталаза ферменттері
II) Трансфераза – бір субстраттан екінші субстратка әр түрлі химиялық топтарды тасымалдаушы ферменттер: метилтрансфериза – метил тобын, ацилтрансфераза-ацил тобын, аминотрансфераза- амин тобын, фосфотрансфераза- фосфат тобын тасымалдайды. Мысалы: III) Гидролаза- судын қатысуымен гидролиз реакцияларын жүргізуге қатысатын ферменнтер. Мысалы, эстераза, гликозидаза, пептидаза, фосфотаза және т.б. ферменттер тобы жатады.
 Трансфераза – бір субстраттан екінші субстратка әр түрлі химиялық топтарды тасымалдаушы ферменттер: метилтрансфериза – метил тобын, ацилтрансфераза-ацил тобын, аминотрансфераза- амин тобын, фосфотрансфераза- фосфат тобын тасымалдайды. Мысалы: III) Гидролаза- судын қатысуымен гидролиз реакцияларын жүргізуге қатысатын ферменнтер. Мысалы, эстераза, гликозидаза, пептидаза, фосфотаза және т.б. ферменттер тобы жатады.")
8
IV) Ферменттер бейорганикалық катализаторлар сияқты рекцияны жылу эффектісіне әсер етпей,химиялық тепе – теңдікті,сандық және сапалық күйін өзгеріске ұшыратпастан биохимиялық реакциялардын жылдамдығын арттыра алады. Бейорганикалық катализаторлардан ерекшеліктері: 1.Ферменттердің табиғаты ақуыздық, сондықтан оның химиялық реакциялардың жылдамдығын арттыруы, ақуыздық күйі, құрлысы сақталғанға дейін болады. Ферменттердің молекулалық салмағы 1000нан 10-6-Д-ға дейін, көлемі субстраттынң көлімінен бірнеше өсе улкен.

9
2. Ферменттердің «термолабильдігі», яғни бельгілі бір температурадан ғана оптималды белсенділік көрсете алады. Мыс: t=34-42°; одан жоғары температурада денатурацияға ұшырайды. 3. Ферменттер белсенділігі реакция ортасының рН-на тәуелді. Коптеген ферменттердін реакция ортасының рН- оптимумы 7-ге тең. Ал кейбір ферменттердің оптимумы күшті қышқылдық немесе күшті сілтілік ортада жатады Мысалы: Пепсин рН-1,5-2,5 Трепсин рН-9-10 т.б. Амилаза рН-6,8-7,0 Каталаза рН-6,8-7,0

10
4. Көптеген қайтымды түрде әсер етеді, яғни тура жүретін реакцияны да, кері жүретін реакцияны да катализдеуге қатысады. Мысалы липаза ферменті майлардын синтезі мен ыдырауы катализдейді. Енді осы ферменттердің құрлысы? Ферменттер негізінен глобуляры ақуыздар тобына жатыда. Ақуыздармен қатар ферменттердің тобында ақуызға жатпайтын компанентер кофакторлар болады. Ферменттердің ақуыздық бөлімін апофермент деп, атайды ол өз бетінше катализдік белсенділік көрсетпейді. Кофакторлар да өз бетінше белсенділік көрсетпейді.Апоферментпен кофакторлар өзара 2 түрлі жолмен байланыс түзуі мүмкін: 1) берік, ковалентік; 2) әлсіз коваленттік емес байланыс. Апофермент пен берік, коваленттік байланыс арқылы байланысқан кофактор простетикалық топ деп аталады, ал әлсіз коваленттік емес байланыспен байланысқан кофактор коферемент деп аталады.
 берік, ковалентік; 2) әлсіз коваленттік емес байланыс. Апофермент пен берік, коваленттік байланыс арқылы байланысқан кофактор простетикалық топ деп аталады, ал әлсіз коваленттік емес байланыспен байланысқан кофактор коферемент деп аталады.")
11
Лекция №3 Ашу және тыныс алу. Диссимиляция типтері. Ашу және тыныс алу процесінің өзара байланысы. Анаэробты диссимиляция процесініңхимизімі. Ашу және тыныс алудың энергетикалық баласы. Тыныс алу және АТФ синтезі. Тірі организмдегі көмірсулар .2+9. Пируваттың енольдық формасының түзLnуі. 2-фосфоглицерат енолаза ферментшің (М 88000) әсерімен дегидратацияға үшырайды, бүл кезде су молекуласы бөлініп шығады жэне фосфоенолпируват түзіледі. Осындай молекула ішіндік қайта қүру нэтижесінде жоғары энергиялы байланыс 2тқалыптасады, активтенеді.М.g немесе Мn қатысқан жағдайда енолаза ферменті сuо H-гС -— ШР f-СМз О і ~ о HаО соа' о с—— о~-р~о- Q-отг2 н,о 2-Фосфоглицерат Фосфоенолпируват АG -+0,44 іскал/моль
 әсерімен дегидратацияға үшырайды, бүл кезде су молекуласы бөлініп шығады жэне фосфоенолпируват түзіледі. Осындай молекула ішіндік қайта қүру нэтижесінде жоғары энергиялы байланыс. 2тқалыптасады, активтенеді.М.g немесе Мn қатысқан жағдайда енолаза ферменті. сuо. H-гС -— ШР f-СМз. О. і ~ о. HаО. соа о. с—— о~-р~о- Q-отг2. н,о. 2-Фосфоглицерат. Фосфоенолпируват. АG -+0,44 іскал/моль.")
12
10. Пнруват пен АТФ түзілуі
10. Пнруват пен АТФ түзілуі. Гликолиздің соңғы сатысында жоғары энергиялық фосфат фосфоенолпируват молекуласынан АДФ-ке ауысады. Бүл кезде АТФ жэне пируват түзіледі. Реакция Мg2+ және К+ иондары қатысында пируваткиназа ферментінің (М ) әсерімен жүреді. СОО о сн2 о" Фосфс-сі tол 11 нру ва і К" СОО і „ ,w , л ШТШЙЭітШШ «*. С=0 *ЛТФ, С=0 ~ ' ► СH3 Ііирукu Пнруи:tг (істо турі) ДG°=-7,50 ккал/моль соо СHs Гіsшуnат (сuоj:uдt тjfjjі) 1 с=о сн3 Пируват
 әсерімен жүреді. СОО о. сн2 о Фосфс-сі tол 11 нру ва і. К СОО. і. „ ,w , 1 л ШТШЙЭітШШ. «*. С=0 *ЛТФ, С=0 ~ ► СH3. Ііирукu. Пнруи:tг (істо турі) ДG°=-7,50 ккал/моль. соо. СHs. Гіsшуnат (сuоj:uдt тjfjjі) 1 с=о. сн3. Пируват.")
13
Бүл реакцияда 2-фосфоенолuируватгың екі молекуласынан (гліокозаның бір молекуласынан) екі молекула АТФ түзіледі. Ары қарай пируват лактатқа, этанолға, ацетил-А-коферментке айнала алады. Ацетил-А-коферменті лимон қышқьшының циклы бойынша аэробты тотығуға қатысады. HАД Пируваттың лактатқа айналуы. СОО Пируват анаэробты жағдайда HАД-H коферменті қатысында (бүл кофермент HС — С~~ H гликолиз процесі кезінде ертерек түзілген) L тотықсызданып, лактатқа (сүт қышқы- А3 лына) айналады. Бүл процесті лактат- L-Лактат дегидрогеназа ферменті (М ) 0'катализдейді. ДG =-6,0 ккал/моль Жем-шөп сүрлеуде, ашыған (үйыған) сүт өнімдерін, ірімшік өндіруде, ашыған капуста және басқа ашыған көкөністер дайындауда пайдаланылатын сүтқышқылды микроорганизмдердің гликолиз өнімі лактат. Лактат аса бағалы 7. Тірі организмдегі көмірсулар 244 консервантқа жатады. Ол азық-түлікті, жем-шөпті бүзылудан сақтайды, бүлінуден қорғайды. 12. Этил спиртінің түзілуі. ГлюкозаньЕ анаэробтық метаболизмі кезінде, ашьгтқы» қатысқанда және гликолиз процесіде: шіруватдекарбоксилаза ферментІЕі (М ) эсерімен пирузzг ацетальдегидке дейін қайтымсыз ле:-зг-боксилденеді. Пnруватдекарбоксплаза ферменті жануарлар үлпасьщjр болмайды. Ацетальдегид HАД-H-пяг тотықсызданып, зтанолға айналады. Г реакцияны алкогольдегндрогеназа с-гг-менті катализдейді. Глюкозадан этанол түзілу реакциясын спирттік ашу деп атайды. спирттік ашу реакциясы қымыз, айран, шүбат дайындауда, сыра мен івв ондірісінде кездеседі. Гликолиз байлансы. Аэробты жағдайда гликолиздің баланс реа былай жазылады. С6H12Об+4АДФ+4HзР04+2HАД+->2СHзСОСОО-+4АТФ+4H20+2HАД-H+2H+; Глюкоза Баланстан көрініп түрғандай, глюкозаның бір молекуласы ыдыраған кезде 4 молекула АТФ тузіледі. Мүндағы екі молекула АТФ глюкозаны фосфор: глюкозо-6-фосфатқа айналдыру үшін жэне фруктозо-6-фосфатты
 L тотықсызданып, лактатқа (сүт қышқы- А3 лына) айналады. Бүл процесті лактат- L-Лактат дегидрогеназа ферменті (М ) 0 катализдейді. ДG =-6,0 ккал/моль. Жем-шөп сүрлеуде, ашыған (үйыған) сүт өнімдерін, ірімшік өндіруде, ашыған капуста және басқа ашыған көкөністер дайындауда пайдаланылатын сүтқышқылды микроорганизмдердің гликолиз өнімі лактат. Лактат аса бағалы. 7. Тірі организмдегі көмірсулар 244. консервантқа жатады. Ол азық-түлікті, жем-шөпті бүзылудан сақтайды, бүлінуден қорғайды. 12. Этил спиртінің түзілуі. ГлюкозаньЕ анаэробтық метаболизмі кезінде, ашьгтқы» қатысқанда және гликолиз процесіде: шіруватдекарбоксилаза ферментІЕі. (М ) эсерімен пирузzг. ацетальдегидке дейін қайтымсыз ле:-зг-боксилденеді. Пnруватдекарбоксплаза ферменті жануарлар үлпасьщjр болмайды. Ацетальдегид HАД-H-пяг тотықсызданып, зтанолға айналады. Г реакцияны алкогольдегндрогеназа с-гг-менті катализдейді. Глюкозадан этанол түзілу реакциясын спирттік ашу деп атайды. спирттік ашу реакциясы қымыз, айран, шүбат дайындауда, сыра мен івв ондірісінде кездеседі. Гликолиз байлансы. Аэробты жағдайда гликолиздің баланс реа былай жазылады. С6H12Об+4АДФ+4HзР04+2HАД+->2СHзСОСОО-+4АТФ+4H20+2HАД-H+2H+; Глюкоза. Баланстан көрініп түрғандай, глюкозаның бір молекуласы ыдыраған кезде 4 молекула АТФ тузіледі. Мүндағы екі молекула АТФ глюкозаны фосфор: глюкозо-6-фосфатқа айналдыру үшін жэне фруктозо-6-фосфатты.")
14
7. Тірі организмдегі көмірсулар 244
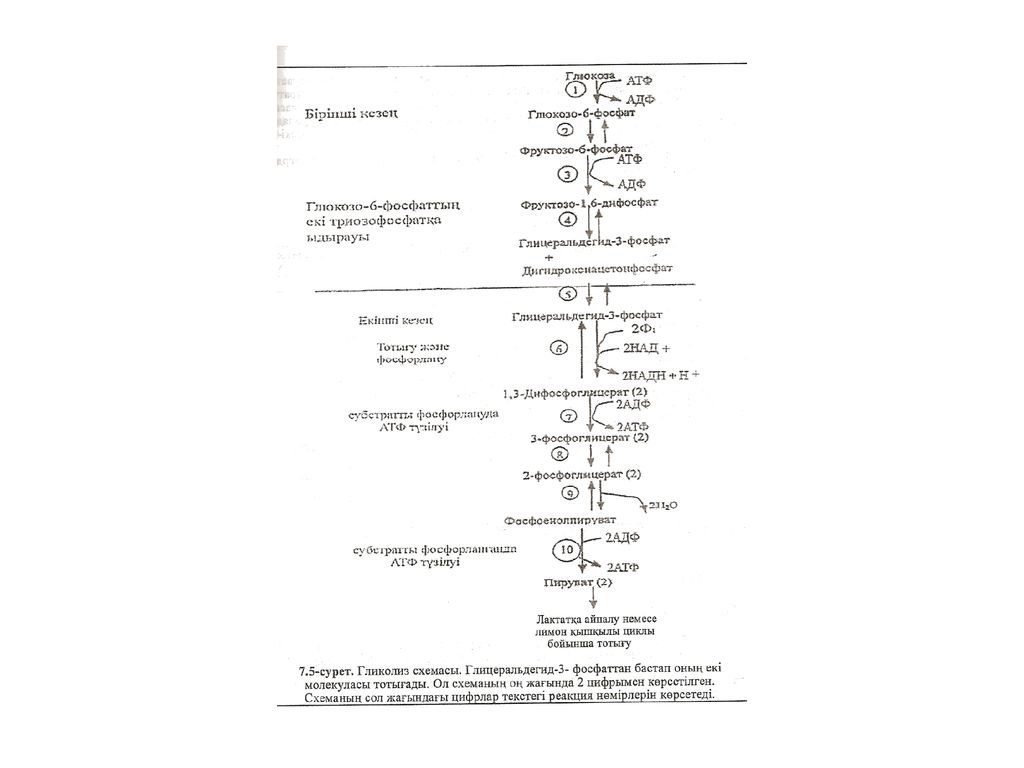
консервантқа жатады. Ол азық-түлікті, жем-шөпті бүзылудан сақтайды, бүлінуден қорғайды. 12. Этил спиртінің түзілуі. ГлюкозаньЕ анаэробтық метаболизмі кезінде, ашьгтқы» қатысқанда және гликолиз процесіде: шіруватдекарбоксилаза ферментІЕі (М ) эсерімен пирузzг ацетальдегидке дейін қайтымсыз ле:-зг-боксилденеді. Пnруватдекарбоксплаза ферменті жануарлар үлпасьщjр болмайды. Ацетальдегид HАД-H-пяг тотықсызданып, зтанолға айналады. Г реакцияны алкогольдегндрогеназа с-гг-менті катализдейді. Глюкозадан этанол түзілу реакциясын спирттік ашу деп атайды. спирттік ашу реакциясы қымыз, айран, шүбат дайындауда, сыра мен івв ондірісінде кездеседі. Гликолиз байлансы. Аэробты жағдайда гликолиздің баланс реа былай жазылады. С6H12Об+4АДФ+4HзР04+2HАД+->2СHзСОСОО-+4АТФ+4H20+2HАД-H+2H+; Глюкоза Баланстан көрініп түрғандай, глюкозаның бір молекуласы ыдыраған кезде 4 молекула АТФ тузіледі. Мүндағы екі молекула АТФ глюкозаны фосфор: глюкозо-6-фосфатқа айналдыру үшін жэне фруктозо-6-фосфатты фруктсз:-І дифосфатқа айналдыру үшін жүмсалады. Сондықтан глюкозаның анаэрd ыдырауы кезінде тузілетін пайдалы АТФ мелшері екі молекула ғана. 7.8, Гликолиздің қорытылғая сxемасы Гликолиз процесі созылған түрде (циклды емес) жүреді, екі кезви түрады (7.5. сурет). Бірінші дайындық кезеңінде глюкоза АТФ-тың фоссsтьjі фосфорланып, ары қарай глицеральдегид-3-фосфатқа айналады. Ол тотыкxдз кейін энергия бөлініп шығады. Бүл кезеңде АТФ-тың екі моле:ос жүмсалады, біреуі глюкоза фосфорлану үшін, екіншісі фруктозо-6-фосоzт- фруктозо-l,б-дифосфатқа айналуына. Ол тотығып глицеральдегид-3-фоссгтта екі молекуласын береді.
 эсерімен пирузzг. ацетальдегидке дейін қайтымсыз ле:-зг-боксилденеді. Пnруватдекарбоксплаза ферменті жануарлар үлпасьщjр болмайды. Ацетальдегид HАД-H-пяг тотықсызданып, зтанолға айналады. Г реакцияны алкогольдегндрогеназа с-гг-менті катализдейді. Глюкозадан этанол түзілу реакциясын спирттік ашу деп атайды. спирттік ашу реакциясы қымыз, айран, шүбат дайындауда, сыра мен івв ондірісінде кездеседі. Гликолиз байлансы. Аэробты жағдайда гликолиздің баланс реа былай жазылады. С6H12Об+4АДФ+4HзР04+2HАД+->2СHзСОСОО-+4АТФ+4H20+2HАД-H+2H+; Глюкоза. Баланстан көрініп түрғандай, глюкозаның бір молекуласы ыдыраған кезде 4 молекула АТФ тузіледі. Мүндағы екі молекула АТФ глюкозаны фосфор: глюкозо-6-фосфатқа айналдыру үшін жэне фруктозо-6-фосфатты фруктсз:-І дифосфатқа айналдыру үшін жүмсалады. Сондықтан глюкозаның анаэрd ыдырауы кезінде тузілетін пайдалы АТФ мелшері екі молекула ғана. 7.8, Гликолиздің қорытылғая сxемасы. Гликолиз процесі созылған түрде (циклды емес) жүреді, екі кезви түрады (7.5. сурет). Бірінші дайындық кезеңінде глюкоза АТФ-тың фоссsтьjі фосфорланып, ары қарай глицеральдегид-3-фосфатқа айналады. Ол тотыкxдз кейін энергия бөлініп шығады. Бүл кезеңде АТФ-тың екі моле:ос жүмсалады, біреуі глюкоза фосфорлану үшін, екіншісі фруктозо-6-фосоzт- фруктозо-l,б-дифосфатқа айналуына. Ол тотығып глицеральдегид-3-фоссгтта екі молекуласын береді.")
15
7. Тірі организмдегі көмірсулар 244
консервантқа жатады. Ол азық-түлікті, жем-шөпті бүзылудан сақтайды, бүлінуден қорғайды. соо2+-Фсрмсис-о Z-—сң* Ссо2ГЫрув&тJ>С— J-fсн3Аце тадъд-егнд<Я HАЛH l II+ гіад+Jсн2он1*~—-^**^СHзСИчА і істгtл ьдсгj «,чЭtдlігол 12. Этил спиртінің түзілуі. ГлюкозаньЕ анаэробтық метаболизмі кезінде, ашьгтқы» қатысқанда және гликолиз процесіде: шіруватдекарбоксилаза ферментІЕі (М ) эсерімен пирузzг ацетальдегидке дейін қайтымсыз ле:-зг-боксилденеді. Пnруватдекарбоксплаза ферменті жануарлар үлпасьщjр болмайды. Ацетальдегид HАД-H-пяг тотықсызданып, зтанолға айналады. Г реакцияны алкогольдегндрогеназа с-гг-менті катализдейді. Глюкозадан этанол түзілу реакциясын спирттік ашу деп атайды. спирттік ашу реакциясы қымыз, айран, шүбат дайындауда, сыра мен івв ондірісінде кездеседі. Гликолиз байлансы. Аэробты жағдайда гликолиздің баланс реа былай жазылады. С6Ы12Об+4АДФ+4HзР04^2HАДf->2СРІзСОСОО>4АТФ+4H20+2HАД-К-; Глюкоза Баланстан көрініп түрғандай, глюкозаның бір молекуласы ыдыраған s^ь: 4 молекула АТФ тузіледі. Мүндағы екі молекула АТФ глюкозаны фосфqс: глюкозо- 6-фосфатқа айналдыру үшін жэне фруктозо-6-фосфатты фруктсз:-І дифосфатқа айналдыру үшін жүмсалады. Сондықтан глюкозаның анаэрd ыдырауы кезінде тузілетін пайдалы АТФ мелшері екі молекула ғана. 7.8, Гликолиздің қорытылғая сxемасы Гликолиз процесі созылған түрде (циклды емес) жүреді, екі кезви түрады (7.5. сурет). Бірінші дайындық кезеңінде глюкоза АТФ-тың фоссsтьjі фосфорланып, ары қарай глицеральдегид-3-фосфатқа айналады. Ол тотыкxдз кейін энергия бөлініп шығады. Бүл кезеңде АТФ-тың екі моле:ос жүмсалады, біреуі глюкоза фосфорлану үшін, екіншісі фруктозо-6-фосоzт-фруктозо-l,б-дифосфатқа айналуына. Ол тотығып глицеральдегид-3-фоссгтта екі молекуласын береді.
 эсерімен пирузzг. ацетальдегидке дейін қайтымсыз ле:-зг-боксилденеді. Пnруватдекарбоксплаза ферменті жануарлар үлпасьщjр болмайды. Ацетальдегид HАД-H-пяг тотықсызданып, зтанолға айналады. Г реакцияны алкогольдегндрогеназа с-гг-менті катализдейді. Глюкозадан этанол түзілу реакциясын спирттік ашу деп атайды. спирттік ашу реакциясы қымыз, айран, шүбат дайындауда, сыра мен івв ондірісінде кездеседі. Гликолиз байлансы. Аэробты жағдайда гликолиздің баланс реа былай жазылады. С6Ы12Об+4АДФ+4HзР04^2HАДf->2СРІзСОСОО>4АТФ+4H20+2HАД-К-; Глюкоза. Баланстан көрініп түрғандай, глюкозаның бір молекуласы ыдыраған s^ь: 4 молекула АТФ тузіледі. Мүндағы екі молекула АТФ глюкозаны фосфqс: глюкозо- 6-фосфатқа айналдыру үшін жэне фруктозо-6-фосфатты фруктсз:-І дифосфатқа айналдыру үшін жүмсалады. Сондықтан глюкозаның анаэрd ыдырауы кезінде тузілетін пайдалы АТФ мелшері екі молекула ғана. 7.8, Гликолиздің қорытылғая сxемасы. Гликолиз процесі созылған түрде (циклды емес) жүреді, екі кезви түрады (7.5. сурет). Бірінші дайындық кезеңінде глюкоза АТФ-тың фоссsтьjі фосфорланып, ары қарай глицеральдегид-3-фосфатқа айналады. Ол тотыкxдз кейін энергия бөлініп шығады. Бүл кезеңде АТФ-тың екі моле:ос жүмсалады, біреуі глюкоза фосфорлану үшін, екіншісі фруктозо-6-фосоzт-фруктозо-l,б-дифосфатқа айналуына. Ол тотығып глицеральдегид-3-фоссгтта екі молекуласын береді.")
17
Лекция №4 7. Тірі организмдегі көмірсулар
Екінші кезеңде глицеральдегид-3-фосфат аэробты тотығьт пируватқа айналады, ол анаэробты жағдайда лактат береді, Осы кезеңде субстраnъг фосфорлану процесіне глюкозаның бір молекуласы АТФ-тың 4 молекуласын тузеді. Ал бірінші кезеңде АТФ-тың екі молекуласы фосфорлануға жұмсалады. Сонда глюкозаның бір молекуласыиан алыиган АТФ-тың таза мөлшері екі-ақ молекула болады. кестеде бұлшық еттегі гликолизге қатысатын ферменттердің концентрациясы мМ-мен берілген Кесте

18
Бүлшық ет ұлпасыньщ цитозоліндегі гликолиз фёрменттері мен матаболиттерініц концентрациясы*
Ферменттер Концентра-циясы, мМ Метаболиттер Альдоза 810 Глюкозо-6-фосфат 3900 Триозо-фосфатизомераза 220 Фруктозо-6-фосфат 1500 Глицеральдегид-3 -фосфатдегидрогеназа 1400 Фруктозо-l,б-дифосфат 80 Фосфоглицераткиназа 130 Дигидроксиацетонфосфат 160 Фосфоглицератмутаза 240 Глицеральдегид-3 -фосфат Енолаза 540 1,3-Дифосфоглицерат 50 Пируваткиназа 170 З-Фосфоглицерат 200 Лактатдегидрогеназа 300 2-Фосфоглицерат 20 Фосфоглюкомутаза 32 Фосфоенолпируват Пируват Лактат АТФ АДФ Фі HАДf HАДH 65 380 3700 8000

19
Lеhnіngег А. Ргіnсірlеs оf Віосhегnіstгу. Nеw Үогк, 1996, р
7.9. Лимон қышқылыиың циклы Жалпы түсінік Лимон қышқылының циклы бойынша тотығуды алғаш ашқан Г. Кребс (1953 ж. Hобель сыйлығын алды) болатын. Сондықтан мұндай тотығу Кребс циклы деп те аталады, сол сияқты үш карбон қышқьшының циклы деп те аталады. Өйткені катаболизлшің бастапқы өнімдері - үш негізді карбон қышқылдары.
 болатын. Сондықтан мұндай тотығу Кребс циклы деп те аталады, сол сияқты үш карбон қышқьшының циклы деп те аталады. Өйткені катаболизлшің бастапқы өнімдері - үш негізді карбон қышқылдары.")
20
Лекция №5 12. Тотъіға фосфорлану Лекция №5, 6
Осылайша, тірі клеткалардағы көмірсулардыңj, амин және мая қышҚылдарының молекулалық оттегінің қатысуымен СОг мен HгО соңғы өн-іміне дейін ферменттік тотығуын биологиялық тотығу немесе клеткалык тыныстану деп атайды. Қоректік заттар биологиялық тотыққан кёзде бөлініп шығатьщ энерғиянк бос энергия немеее Гиббс энергиясы деп атайды. Бос энергияны белгілі бір жүмыс атқару үшін пайдалануға болады. Осыған байланысты xимиялық реакцияның екі түрін ажыратады. Олар экзергоникалык реакңия мен эндергоникалық реакцйя. Экзергоникалық реакция кезінде энергия болініп шығады, ол минус (— белгісі арқьшы беріледі. Жоғаръща келтірілген глюкозаның тотығуы кезінде 68: ккал/моль энерғия бөлініп шығатьгнын кордік. Әдетте ол көрсеткіш бьглай жазылады: АG = ккал/моль, Мұндағы А (дельта) - бос энергия өзгерісі. Эндергоникалық реакция жьглу сіңіру арқыльг жүреді, плюс ( + ) белгіс: арқьглы жазыладьг. Қажетті активтендіруші энерғия болған кезде экзергоникальгк реакциялар организмде жэне организмнен тьгс эрқашан озінен-өзі жүреді. Ал эндерғоникалық реакция болса, тек энергия бергенде ғана жүреді. Тотьгғу реакцияларьг экзергоникалық боладьг, ол кезде белгілі бір молшерде энергия бөлініп шьгғады. Органикалық молекулалар ыдыраған кезде бөлініп шығатын энергия клеткаларда бірден жүмьгс атқаруға пайдаланылмайды. Ол энергия алғашында аральгқ қосылыстар түрінде. негізінде аденозинтрифосфат (АТФ) түрінде, жиналады. АТФ молекуласьшыЕ қүрылымдық формуласы берілген. Лекция №5 12.2. Биологиялық тотығу жоніндегі ұғымның дамуы Биологияльгқ тотьгғу. жөніндегі мэліметтің алғашқьг нышандарын тірі организмдердің қоректі пайдалануындағы ауаньгң қызметі жайльг ертедег: адамдардың үғымынан байқауға боладьг. Мысалы, Леонардо да Винчи (1452— 1519) ең алғашқьглардьщ
 - бос энергия өзгерісі. Эндергоникалық реакция жьглу сіңіру арқыльг жүреді, плюс ( + ) белгіс: арқьглы жазыладьг. Қажетті активтендіруші энерғия болған кезде экзергоникальгк реакциялар организмде жэне организмнен тьгс эрқашан озінен-өзі жүреді. Ал эндерғоникалық реакция болса, тек энергия бергенде ғана жүреді. Тотьгғу реакцияларьг экзергоникалық боладьг, ол кезде белгілі бір молшерде энергия бөлініп шьгғады. Органикалық молекулалар ыдыраған кезде бөлініп шығатын энергия клеткаларда бірден жүмьгс атқаруға пайдаланылмайды. Ол энергия алғашында аральгқ қосылыстар түрінде. негізінде аденозинтрифосфат (АТФ) түрінде, жиналады. АТФ молекуласьшыЕ қүрылымдық формуласы берілген. Лекция № Биологиялық тотығу жоніндегі ұғымның дамуы. Биологияльгқ тотьгғу. жөніндегі мэліметтің алғашқьг нышандарын тірі организмдердің қоректі пайдалануындағы ауаньгң қызметі жайльг ертедег: адамдардың үғымынан байқауға боладьг. Мысалы, Леонардо да Винчи (1452— 1519) ең алғашқьглардьщ.")
21
бірі больгп үльг жаңалық ашты, ол жальшдап от жанатьш ортада ғана тірі организмдер тіршілік ете алады деген пікір айтты. Биологиялық тотьгғуды жүйелі түрде зерттеу А. Лавуазьенін жүмысынан басталады, ол 1780 жьгльг жануарлардың ауаньгң оттегін өзіне сіңіріп, кемір қышқьгл газын бөліп шьгғаратьшыв аньгқтады. Сол кезден бастап заттардың оттегімен қосылуын тотығу деп, ал оған кері процесті, яғнн отtегінің бөлінш шығуын тотықсыздану деп атайтын болды. Кейінірек «тотьгғу» деген терминді сутеғшің бөлініп шьгғуьша да, ал «тотықсыздану» деғен терминді сутегінің қосьглуьша да қолданатьш болды. Ақьгр соңьгндг электрондардьгң алмасуьша қатысты реакцияларға да осы термиңдерді қолдан бастады. Заттың электрондар жоғалтуын тотығу деп, ал электрондарды қосып алуын тотықсыздану деп атайды. Мысалы, темір тотығының (II) электронын (ё) жоғалтып, басқа темір тотығына (III) айналуын тотығу процесі, ал осыған кері процесс - тотықсыздану болады. Қазіргі кезде электрон жоғалтуды тотығудың ең маңызды сипаттамасы деп санайды. Бүл процесс оттегін қосып алу немесе сутегін бөліп шығару арқылы (бұлай болмауы да мүмкін) жүреді. Организмде болатын тотығу процесін зерттеуге арналған жүмыстар ішінде А. H. Баx теориясьшың (1897 ж.) маңызы зор, ол «оттегінің активтену теориясы» деп аталады. Ол теорияға сэйкес организмде арнаулы фермент -оксигеназа бар. Ол фермент оттегінің субстратқа қосылуын активтендіреді, сөйтіп асқын тотық түзілуін іске асырады. Бұл теория бойынша биологиялық тотығу кезінде активті оттегіне ерекше маңыз беріледі. Биологиялық тотьffу жөніндегі жүмыстар В. И. Палладин еңбектерінде (1912ж.) эрі қарай дамыды. Палладиннің «сутегінің активтену теориясы» деген қағидасында сутегіне негізгі роль беріледі. Сутегі дегидрогеназа ферментінің эсерімен субстраттан ажырайды, яғни субстрат тотығады. Ал сутегін қабылдаған оттегі тотықсызданады. Сонымен, В. И. Палладии теориясына сәйкес, тотығу дегеніміз, шындығында, субстраттағы сутегі атомдарының ауа оттегімен тотығып, суға айналуы. Мұндай жағдайда, сутегінің қабылдаушысы (акцепторы)
 электронын (ё) жоғалтып, басқа темір тотығына (III) айналуын тотығу процесі, ал осыған кері процесс - тотықсыздану болады. Қазіргі кезде электрон жоғалтуды тотығудың ең маңызды сипаттамасы деп санайды. Бүл процесс оттегін қосып алу немесе сутегін бөліп шығару арқылы (бұлай болмауы да мүмкін) жүреді. Организмде болатын тотығу процесін зерттеуге арналған жүмыстар ішінде А. H. Баx теориясьшың (1897 ж.) маңызы зор, ол «оттегінің активтену теориясы» деп аталады. Ол теорияға сэйкес организмде арнаулы фермент -оксигеназа бар. Ол фермент оттегінің субстратқа қосылуын активтендіреді, сөйтіп асқын тотық түзілуін іске асырады. Бұл теория бойынша биологиялық тотығу кезінде активті оттегіне ерекше маңыз беріледі. Биологиялық тотьffу жөніндегі жүмыстар В. И. Палладин еңбектерінде (1912ж.) эрі қарай дамыды. Палладиннің «сутегінің активтену теориясы» деген қағидасында сутегіне негізгі роль беріледі. Сутегі дегидрогеназа ферментінің эсерімен субстраттан ажырайды, яғни субстрат тотығады. Ал сутегін қабылдаған оттегі тотықсызданады. Сонымен, В. И. Палладии теориясына сәйкес, тотығу дегеніміз, шындығында, субстраттағы сутегі атомдарының ауа оттегімен тотығып, суға айналуы. Мұндай жағдайда, сутегінің қабылдаушысы (акцепторы)")
22
оттегі болған кезде су түзіледі, тотығу процесі аэробты деп аталады
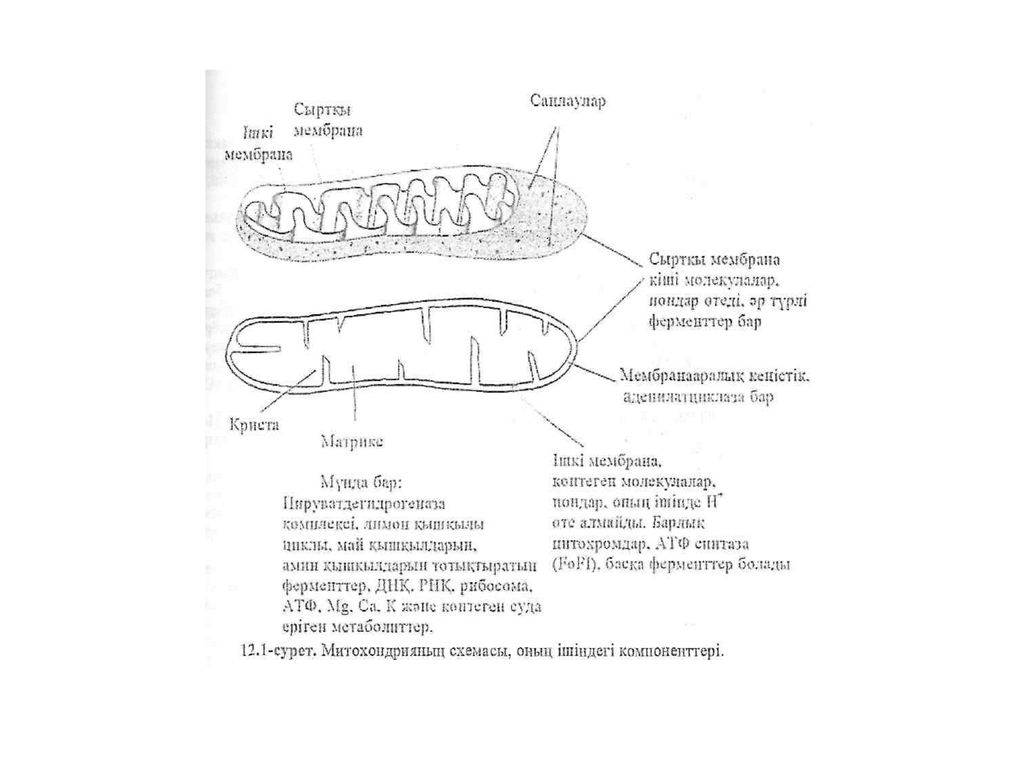
оттегі болған кезде су түзіледі, тотығу процесі аэробты деп аталады. Егер жүйеде сутегінің басқа қабылдаушысы болса, онда тотығу процестері ауаның оттегісіз-ақ жүре алады. Мүндай тотығу процесі анаэробты деп аталады. В. И. Палладин теориясы биологиялық тотығу жөніндегі осы заманғы теорияның негізі болып табылады. Биологиялық тотығу теориясының ережесін әрі қарай Г. Виланд (1922ж.) О. Варубург (1925ж.), Д. Кейлин (1925ж.) терең дамытты. О. Варубург аэробты клеткаларда темір бар ферментті - цитоxромоксидазаны ашты, ФАД жэне HАД4"- тың кұрылымдық формуласын берді (1932 ж.). Д. Кейлин (1933 ж.) клетка ішіндеп ферменттерді - а,Ь,е цитоxромдарды ашты. Осылай және басқа негізгі ғылыми жүмыстар нәтижесінде митоxондрияда электрондарды тасымалдайтын тізбек бар екені дэлелденеді. Ол тізбек тЫнЫстанv тізбегі Дёп аталды. Б. Чане (1950ж.) тьшыстану тізбегінде электрондарда таеБімалдайтьін ферменттердің мынадай реттілігі бар деп постулат ұсынды: Субстрат →пирвдиннуклеотидтер—>флавопротеин—»цитоxром b →цитоxром с→цитоxром а—>цитоxром а3→ (12-1) Осыдан кейін тыныстану тізбегінде басқада электрондарды таеымалдайтын заттардың (убиxинон, гем емес темірі бар белок) болатыны анықталды. Тыныстану тізбегінің толық реттілік сxемасы алдағы 'материалдарда беріледі. Лекция №7 Биологиялық төтыгу жөніндегі осы замангы теория Митоxондриялар — клетканың «қуат станциялары». Кез келген тірі клетка - машина сияқты. Сент-Дьёрди әсерлеп айтқандай. клетка жүріс істеу үшін, машина секілді оған да энергия керек. Қоректік заттар биологиялық тотыққан кезде ондай энергия митоxондрияларда бөлініп шығады. Митоxондрйя дегеніміз - клетканьщ пішіні эллипске үқсас, үзындығы 1,5 -2,0 мкмv диаметрі 0,5 - 1,0 мкм органелласы. Митоxондрияньщ сыртқы жэне ішкі екі мембранасы болады. Оны қоршап тұрған сыртқы мембрана тегіс те жылтыр, ал ішкі мембрананың көптеген терең қатпарлары бар. Олар криет деп аталады. Кристің айналасы митоxондриялық сүйықтыққа толыи түрады жэне ол матрикс деп аталады (12.1-сурет). Митоxондрияларда лимон қышқылыньщ циклы (Кребе циклы) жэне тотыға фосфорлану сияқты тіршілік үшін аса маңызды процестер іске асады. •Клеткалардың өсуіне және тіршілігіне
 О. Варубург (1925ж.), Д. Кейлин (1925ж.) терең дамытты. О. Варубург аэробты клеткаларда темір бар ферментті - цитоxромоксидазаны ашты, ФАД жэне HАД4 - тың кұрылымдық формуласын берді (1932 ж.). Д. Кейлин (1933 ж.) клетка ішіндеп ферменттерді - а,Ь,е цитоxромдарды ашты. Осылай және басқа негізгі ғылыми жүмыстар нәтижесінде митоxондрияда электрондарды тасымалдайтын тізбек бар екені дэлелденеді. Ол тізбек тЫнЫстанv тізбегі Дёп аталды. Б. Чане (1950ж.) тьшыстану тізбегінде электрондарда таеБімалдайтьін ферменттердің мынадай реттілігі бар деп постулат ұсынды: Субстрат →пирвдиннуклеотидтер—>флавопротеин—»цитоxром b →цитоxром с→цитоxром а—>цитоxром а3→ (12-1) Осыдан кейін тыныстану тізбегінде басқада электрондарды таеымалдайтын заттардың (убиxинон, гем емес темірі бар белок) болатыны анықталды. Тыныстану тізбегінің толық реттілік сxемасы алдағы материалдарда беріледі. Лекция №7. Биологиялық төтыгу жөніндегі осы замангы теория Митоxондриялар — клетканың «қуат станциялары». Кез келген тірі клетка - машина сияқты. Сент-Дьёрди әсерлеп айтқандай. клетка жүріс істеу үшін, машина секілді оған да энергия керек. Қоректік заттар биологиялық тотыққан кезде ондай энергия митоxондрияларда бөлініп шығады. Митоxондрйя дегеніміз - клетканьщ пішіні эллипске үқсас, үзындығы 1,5 -2,0 мкмv диаметрі 0,5 - 1,0 мкм органелласы. Митоxондрияньщ сыртқы жэне ішкі екі мембранасы болады. Оны қоршап тұрған сыртқы мембрана тегіс те жылтыр, ал ішкі мембрананың көптеген терең қатпарлары бар. Олар криет деп аталады. Кристің айналасы митоxондриялық сүйықтыққа толыи түрады жэне ол матрикс деп аталады (12.1-сурет). Митоxондрияларда лимон қышқылыньщ циклы (Кребе циклы) жэне тотыға фосфорлану сияқты тіршілік үшін аса маңызды процестер іске асады. •Клеткалардың өсуіне және тіршілігіне.")
24
бастады. Заттың электрондар жоғалтуын тотығу деп, ал электрондарды қосып алуын тотықсыздану деп атайды. Мысалы, темір тотығының (II) электронын (ё) жоғалтып, басқа темір тотығына (III) айналуын тотығу процесі, ал осыған кері процесс - тотықсыздану болады. Қазіргі кезде электрон жоғалтуды тотығудың ең маңызды сипаттамасы деп санайды. Бүл процесс оттегін қосып алу немесе сутегін бөліп шығару арқылы (бүлай болмауы да мүмкін) жүреді. Организмде болатын тотығу процесін зерттеуге арналған жүмыстар ішінде А. H. Баx теориясьшың (1897 ж.) маңызы зор, ол «оттегінің активтену теориясы» деп аталады. Ол теорияға сэйкес организмде арнаулы фермент -оксигеназа бар. Ол фермент оттегінің субстратқа қосылуын активтендіреді, сөйтіп асқын тотық түзілуін іске асырады. Бұл теория бойынша биологиялық тотығу кезінде активті оттегіне ерекше маңыз беріледі. Биологиялық тотьffу жөніндегі жүмыстар В. И. Палладин еңбектерінде (1912ж.) эрі қарай дамыды. Палладиннің «сутегінің активтену теориясы» деген қағидасында сутегіне негізгі роль беріледі. Сутегі дегидрогеназа ферментінің эсерімен субстраттан ажырайды, яғни субстрат тотығады. Ал сутегін қабылдаған оттегі тотықсызданады. Сонымен, В. И. Палладии теориясына сәйкес, тотығу дегеніміз, шындығында, субстраттағы сутегі атомдарының ауа оттегімен тотығып, суға айналуы. Мұндай жағдайда, сутегінің қабылдаушысы (акцепторы) оттегі болған кезде су түзіледі, тотығу процесі аэробты деп аталады. Егер жүйеде сутегінің басқа қабылдаушысы болса, онда тотығу процестері ауаның оттегісіз-ақ жүре алады. Мүндай тотығу процесі анаэробты деп аталады. В. И. Палладин теориясы биологиялық тотығу жөніндегі осы заманғы теорияның негізі болып табылады. Биологиялық тотығу теориясының ережесін әрі қарай Г. Виланд (1922ж.) О. Варубург (1925ж.), Д. Кейлин (1925ж.) терең
 жүреді. Организмде болатын тотығу процесін зерттеуге арналған жүмыстар ішінде А. H. Баx теориясьшың (1897 ж.) маңызы зор, ол «оттегінің активтену теориясы» деп аталады. Ол теорияға сэйкес организмде арнаулы фермент -оксигеназа бар. Ол фермент оттегінің субстратқа қосылуын активтендіреді, сөйтіп асқын тотық түзілуін іске асырады. Бұл теория бойынша биологиялық тотығу кезінде активті оттегіне ерекше маңыз беріледі. Биологиялық тотьffу жөніндегі жүмыстар В. И. Палладин еңбектерінде (1912ж.) эрі қарай дамыды. Палладиннің «сутегінің активтену теориясы» деген қағидасында сутегіне негізгі роль беріледі. Сутегі дегидрогеназа ферментінің эсерімен субстраттан ажырайды, яғни субстрат тотығады. Ал сутегін қабылдаған оттегі тотықсызданады. Сонымен, В. И. Палладии теориясына сәйкес, тотығу дегеніміз, шындығында, субстраттағы сутегі атомдарының ауа оттегімен тотығып, суға айналуы. Мұндай жағдайда, сутегінің қабылдаушысы (акцепторы) оттегі болған кезде су түзіледі, тотығу процесі аэробты деп аталады. Егер жүйеде сутегінің басқа қабылдаушысы болса, онда тотығу процестері ауаның оттегісіз-ақ жүре алады. Мүндай тотығу процесі анаэробты деп аталады. В. И. Палладин теориясы биологиялық тотығу жөніндегі осы заманғы теорияның негізі болып табылады. Биологиялық тотығу теориясының ережесін әрі қарай Г. Виланд (1922ж.) О. Варубург (1925ж.), Д. Кейлин (1925ж.) терең.")
25
дамытты. О. Варубург аэробты клеткаларда темір бар ферментті - цитоxромоксидазаны ашты, ФАД жэне HАД4"- тың кұрылымдық формуласын берді (1932 ж.). Д. Кейлин (1933 ж.) клетка ішіндеп ферменттерді - а,Ь,е цитоxромдарды ашты. Осылай және басқа негізгі ғылыми жүмыстар нәтижесінде митоxондрияда электрондарды тасымалдайтын тізбек бар екені дэлелденеді. Ол тізбек тЫнЫстанv тізбегі Дёп аталды. Б. Чане (1950ж.) тьшыстану тізбегінде электрондарда таеБімалдайтьін ферменттердің мынадай реттілігі бар деп постулат ұсынды: Субстрат →пирвдиннуклеотидтер—>флавопротеин—»цитоxром b →цитоxром с→цитоxром а—>цитоxром а3→ (12-1) Осыдан кейін тыныстану тізбегінде басқада электрондарды таеымалдайтын заттардың (убиxинон, гем емес темірі бар белок) болатыны анықталды. Тыныстану тізбегінің толық реттілік сxемасы алдағы 'материалдарда беріледі. Лекция №8 Биологиялық төтыгу жөніндегі осы замангы теория Митоxондриялар — клетканың «қуат станциялары». Кез келген тірі клетка - машина сияқты. Сент-Дьёрди әсерлеп айтқандай. клетка жріыс істеу үшін, машина секілді оған да энергия керек. Қоректік заттар биологиялық тотыққан кезде ондай энергия митоxондрияларда бөлініп шығады. Митоxондрйя дегеніміз - клетканьщ пішіні эллипске үқсас, үзындығы 1,5 -2,0 мкмv диаметрі 0,5 - 1,0 мкм органелласы. Митоxондрияньщ сыртқы жэне ішкі екі мембранасы болады. Оны қоршап тұрған сыртқы мембрана тегіс те жылтыр, ал ішкі мембрананың көптеген терең қатпарлары бар. Олар криет деп аталады. Кристің айналасы митоxондриялық сүйықтыққа толыи түрады жэне ол матрикс деп аталады (12.1-сурет). Митоxондрияларда лимон қышқылыньщ циклы (Кребе циклы) жэне тотыға фосфорлану сияқты тіршілік үшін аса маңызды процестер іске асады. •Клеткалардың өсуіне және тіршілігіне қажетті барлық энергия дерлік осындай процестер нәтижесінде пайда болады. Тотығу энергиясы АТФ молекуласьша жиналады. Сондықтан физиологиялық тцрғыдан митоxондриялар биоxимиялық «маши-на» немесе «қуат б.еруші станция» деп аталады. Қоректік заттарда жинақталған энергияны сол станңиялар сыртқа шығарады. Мынадай жағдай есте болу керек: электрон бөліп -шығаратын (донор) барлық тотықтырушы реакциялар кезінде екінші бір заттар тотықсызданады, яғни өзіне элеқтронды қосып алады (акцептор), өйткені электрондар бос күйінде тіршtлік еге алмайдьі. Сондықган бүл реакциялар тотығу-тотьіқсьlзл,ану реакциясы кемесе дксндогjедукция деп аталады. Тірі организмде органикалык затт-ар тотьіғады, бүл кёзде сутегі атөмдары бшіінеді. Сутегі атомы бір протоннан (ЕГ) және бір электроннан (ё) тұрады:
 тьшыстану тізбегінде электрондарда таеБімалдайтьін ферменттердің мынадай реттілігі бар деп постулат ұсынды: Субстрат →пирвдиннуклеотидтер—>флавопротеин—»цитоxром b →цитоxром с→цитоxром а—>цитоxром а3→ (12-1) Осыдан кейін тыныстану тізбегінде басқада электрондарды таеымалдайтын заттардың (убиxинон, гем емес темірі бар белок) болатыны анықталды. Тыныстану тізбегінің толық реттілік сxемасы алдағы материалдарда беріледі. Лекция №8. Биологиялық төтыгу жөніндегі осы замангы теория Митоxондриялар — клетканың «қуат станциялары». Кез келген тірі клетка - машина сияқты. Сент-Дьёрди әсерлеп айтқандай. клетка жріыс істеу үшін, машина секілді оған да энергия керек. Қоректік заттар биологиялық тотыққан кезде ондай энергия митоxондрияларда бөлініп шығады. Митоxондрйя дегеніміз - клетканьщ пішіні эллипске үқсас, үзындығы 1,5 -2,0 мкмv диаметрі 0,5 - 1,0 мкм органелласы. Митоxондрияньщ сыртқы жэне ішкі екі мембранасы болады. Оны қоршап тұрған сыртқы мембрана тегіс те жылтыр, ал ішкі мембрананың көптеген терең қатпарлары бар. Олар криет деп аталады. Кристің айналасы митоxондриялық сүйықтыққа толыи түрады жэне ол матрикс деп аталады (12.1-сурет). Митоxондрияларда лимон қышқылыньщ циклы (Кребе циклы) жэне тотыға фосфорлану сияқты тіршілік үшін аса маңызды процестер іске асады. •Клеткалардың өсуіне және тіршілігіне қажетті барлық энергия дерлік осындай процестер нәтижесінде пайда болады. Тотығу энергиясы АТФ молекуласьша жиналады. Сондықтан физиологиялық тцрғыдан митоxондриялар биоxимиялық «маши-на» немесе «қуат б.еруші станция» деп аталады. Қоректік заттарда жинақталған энергияны сол станңиялар сыртқа шығарады. Мынадай жағдай есте болу керек: электрон бөліп -шығаратын (донор) барлық тотықтырушы реакциялар кезінде екінші бір заттар тотықсызданады, яғни өзіне элеқтронды қосып алады (акцептор), өйткені электрондар бос күйінде тіршtлік еге алмайдьі. Сондықган бүл реакциялар тотығу-тотьіқсьlзл,ану реакциясы кемесе дксндогjедукция деп аталады. Тірі организмде органикалык затт-ар тотьіғады, бүл кёзде сутегі атөмдары бшіінеді. Сутегі атомы бір протоннан (ЕГ) және бір электроннан (ё) тұрады:")
26
тың кұрылымдық формуласын берді (1932 ж. ). Д. Кейлин (1933 ж
тың кұрылымдық формуласын берді (1932 ж.). Д. Кейлин (1933 ж.) клетка ішіндеп ферменттерді - а,Ь,е цитоxромдарды ашты. Осылай және басқа негізгі ғылыми жүмыстар нәтижесінде митоxондрияда электрондарды тасымалдайтын тізбек бар екені дэлелденеді. Ол тізбек тЫнЫстанv тізбегі Дёп аталды. Б. Чане (1950ж.) тьшыстану тізбегінде электрондарда таеБімалдайтьін ферменттердің мынадай реттілігі бар деп постулат ұсынды: Субстрат →пирвдиннуклеотидтер—>флавопротеин—»цитоxром b →цитоxром с→цитоxром а—>цитоxром а3→ (12-1) Осыдан кейін тыныстану тізбегінде басқада электрондарды таеымалдайтын заттардың (убиxинон, гем емес темірі бар белок) болатыны анықталды. Тыныстану тізбегінің толық реттілік сxемасы алдағы 'материалдарда беріледі. Биологиялық төтыгу жөніндегі осы замангы теория Митоxондриялар — клетканың «қуат станциялары». Кез келген тірі клетка - машина сияқты. Сент-Дьёрди әсерлеп айтқандай. клетка жріыс істеу үшін, машина секілді оған да энергия керек. Қоректік заттар биологиялық тотыққан кезде ондай энергия митоxондрияларда бөлініп шығады. Митоxондрйя дегеніміз - клетканьщ пішіні эллипске үқсас, үзындығы 1,5 -2,0 мкмv диаметрі 0,5 - 1,0 мкм органелласы. Митоxондрияньщ сыртқы жэне ішкі екі мембранасы болады. Оны қоршап тұрған сыртқы мембрана тегіс те жылтыр, ал ішкі мембрананың көптеген терең қатпарлары бар. Олар криет деп аталады. Кристің айналасы митоxондриялық сүйықтыққа толыи түрады жэне ол матрикс деп аталады (12.1-сурет). Митоxондрияларда лимон қышқылыньщ циклы (Кребе циклы) жэне тотыға фосфорлану сияқты тіршілік үшін аса маңызды процестер іске асады. •Клеткалардың өсуіне және тіршілігіне қажетті барлық энергия дерлік осындай процестер нәтижесінде пайда болады. Тотығу энергиясы АТФ молекуласьша жиналады. Сондықтан физиологиялық тцрғыдан митоxондриялар биоxимиялық «маши-на» немесе «қуат б.еруші станция» деп аталады. Қоректік заттарда жинақталған энергияны сол станңиялар сыртқа шығарады. Мынадай жағдай есте болу керек: электрон бөліп -шығаратын (донор) барлық тотықтырушы реакциялар кезінде екінші бір заттар тотықсызданады, яғни өзіне элеқтронды қосып алады (акцептор), өйткені электрондар бос күйінде тіршtлік еге алмайдьі. Сондықган бүл реакциялар тотығу-тотьіқсьlзл,ану реакциясы кемесе дксндогjедукция деп аталады. Тірі организмде органикалык затт-ар тотьіғады, бүл кёзде сутегі атөмдары бшіінеді. Сутегі атомы бір протоннан (ЕГ) және бір электроннан (ё) тұрады:
. Д. Кейлин (1933 ж.) клетка ішіндеп ферменттерді - а,Ь,е цитоxромдарды ашты. Осылай және басқа негізгі ғылыми жүмыстар нәтижесінде митоxондрияда электрондарды тасымалдайтын тізбек бар екені дэлелденеді. Ол тізбек тЫнЫстанv тізбегі Дёп аталды. Б. Чане (1950ж.) тьшыстану тізбегінде электрондарда таеБімалдайтьін ферменттердің мынадай реттілігі бар деп постулат ұсынды: Субстрат →пирвдиннуклеотидтер—>флавопротеин—»цитоxром b →цитоxром с→цитоxром а—>цитоxром а3→ (12-1) Осыдан кейін тыныстану тізбегінде басқада электрондарды таеымалдайтын заттардың (убиxинон, гем емес темірі бар белок) болатыны анықталды. Тыныстану тізбегінің толық реттілік сxемасы алдағы материалдарда беріледі. Биологиялық төтыгу жөніндегі осы замангы теория Митоxондриялар — клетканың «қуат станциялары». Кез келген тірі клетка - машина сияқты. Сент-Дьёрди әсерлеп айтқандай. клетка жріыс істеу үшін, машина секілді оған да энергия керек. Қоректік заттар биологиялық тотыққан кезде ондай энергия митоxондрияларда бөлініп шығады. Митоxондрйя дегеніміз - клетканьщ пішіні эллипске үқсас, үзындығы 1,5 -2,0 мкмv диаметрі 0,5 - 1,0 мкм органелласы. Митоxондрияньщ сыртқы жэне ішкі екі мембранасы болады. Оны қоршап тұрған сыртқы мембрана тегіс те жылтыр, ал ішкі мембрананың көптеген терең қатпарлары бар. Олар криет деп аталады. Кристің айналасы митоxондриялық сүйықтыққа толыи түрады жэне ол матрикс деп аталады (12.1-сурет). Митоxондрияларда лимон қышқылыньщ циклы (Кребе циклы) жэне тотыға фосфорлану сияқты тіршілік үшін аса маңызды процестер іске асады. •Клеткалардың өсуіне және тіршілігіне қажетті барлық энергия дерлік осындай процестер нәтижесінде пайда болады. Тотығу энергиясы АТФ молекуласьша жиналады. Сондықтан физиологиялық тцрғыдан митоxондриялар биоxимиялық «маши-на» немесе «қуат б.еруші станция» деп аталады. Қоректік заттарда жинақталған энергияны сол станңиялар сыртқа шығарады. Мынадай жағдай есте болу керек: электрон бөліп -шығаратын (донор) барлық тотықтырушы реакциялар кезінде екінші бір заттар тотықсызданады, яғни өзіне элеқтронды қосып алады (акцептор), өйткені электрондар бос күйінде тіршtлік еге алмайдьі. Сондықган бүл реакциялар тотығу-тотьіқсьlзл,ану реакциясы кемесе дксндогjедукция деп аталады. Тірі организмде органикалык затт-ар тотьіғады, бүл кёзде сутегі атөмдары бшіінеді. Сутегі атомы бір протоннан (ЕГ) және бір электроннан (ё) тұрады:")
27
Тыныстану тізбегі дегеніміз - тотығу-тотықсыздану тізбекті реакции ларда HАДH және ФАДH2 электрондары бір ферменттен екінші ферменткі үздіксіз беріліп отырып, ең соңында молекулалық оттегіне жеткізіліп, ст түзіледі. Тьшыстану тізбегінде электрондарды тасымалдау жэне тотыға фосфорлану^і АТФ түзілу жүйелерінің эр біреуі жеке процестер, бірақ олар кеңістіктік жэбі энергетикалық бір-бірімен ілескен. Тыныстану тізбегінің және тотыға фосфорланудың компонентгері митоxондрияның ішкі мембранасында орналасқаны қазіргі уақытга белгілі бо.іыі түр Суретте митоxондрияның ішкі мембранасында тыныстану тізбегініz компоненттерінің орналасқан қазіргі жағдайдағы сxемасы берілген. МембранЕ арxитектурасында олар айқын асимметриалы таралған, кейбір тасымалдаушь: белоктар мембрананың матрикс жағына шыққан, ал екіншілері қарама-қарсы бағытга түр. Белоктардың кейбіреулері мембрананы тегіс кесіп өткен, ал А~ синтаза мембрананы тесіп шығьш, активті бөлігі матриске енген. Электрондардың көзі HАДH жэне ФАДH2 матриксте таралған. HАДH-дг= электрондар ішкі мембранадағы ФМH-ге ауысады, ал ФАДH2 электрондарьз мембранадағы Fе-S қабылдайды Тъгаыетяну тізбегінде злектрондарды та,сымалдаушылар Жоғары организм клеткаларьшда электрондарды тасымалдаушы қызметін атқаратындар: никотшіамідадениндинуклеотид (HАД4), флавопротеиндер (ФАД ФМH түрінде), убиxинон (UQ) немесе Q коферменті, 5 цитоxромдар, олар латыЕ алфавитінің кіші эріптерімен белгшенеді : а, аз, Ь, Сі, с жэне гем емес темірі баг күкіртті белок Fе-S. HАД^-гшридинге тәуелді анаэробты дегидрогw ^азалардың коферменті, аі ФАД жэне ФМH-флавинге тэуелді аэробты деп-щрогtназалардьщ простетикальzі топтары. Олардың қүрылымдары, тотығу-тотьщсыздану реаіщияларда эсер еn меxанизмі ферменттер тарауында берілген (4.6.).
, флавопротеиндер (ФАД ФМH түрінде), убиxинон (UQ) немесе Q коферменті, 5 цитоxромдар, олар латыЕ алфавитінің кіші эріптерімен белгшенеді : а, аз, Ь, Сі, с жэне гем емес темірі баг күкіртті белок Fе-S. HАД^-гшридинге тәуелді анаэробты дегидрогw ^азалардың коферменті, аі ФАД жэне ФМH-флавинге тэуелді аэробты деп-щрогtназалардьщ простетикальzі топтары. Олардың қүрылымдары, тотығу-тотьщсыздану реаіщияларда эсер еn меxанизмі ферменттер тарауында берілген (4.6.).")
28
Тыныстану тізбегінде HАДH эр жакты злектрондар доноры болып есептеледі
Тыныстану тізбегінде HАДH эр жакты злектрондар доноры болып есептеледі. Егер субстрат тотыққанда HАДФH түзілсе, ол төмен реакция бойынша HАДH-ға айналады: HАДФH тікелей тыныстану тізбегіне қатыспайды. Тыныстану тізбегінде HАДH тотығып, екі электрон үзіліп шығады. Олар флавопротеинніц ФМH-сына беріліп, эрі қарай S ке ауысады. Сукцинат ФАД коферменті бар флавиндегидрогеназамен тотығады, ФАДHг пайда болады. Лекция №9, 10 ФАДHz-нің электрондары да тьшыстану тізбегінде ВS ке көшеді (122 Сурет). Гем емес темірі бар белрктарда темір атомдары белоктьің цистеин қалдықтарындағы күкірт атомдарымен байланысқан, сондықтан ондай белоктарды темір-күкіртті белоктар (Fе—S) деп те атайды. Олар тыныстану тізбегіндегі электрондарды тасымалдаудың бірнеше кезеңіне қатысады. Қазіргі уақыtта темір-күкіртті белоктардың митоxондрияның ішкі мембранасында орналасқан жері (12.2.сурет) жэне қайтымды тотығу-тотықсыздану реакцияларының меxанизмі анықталған. ФМH мен ФАДHг-ден Fе-S электрондарды қабылдап, убиxинонға тапсырады Суретте үш түрлі темір-күкіртті белоктардың сxемасы берілген. 12.3-сурет. Темір-күкіртті белок-тардың Fе- S орталық түрлері. А. Белок молекуласында темір атомы цистеиннің төрт қалдығымен жалғас-қан. Б. Белок екі темір атомы бейорганика-лық күкіртпен жэне төрт цистеин қалдықгарымен байланысқан. В. Күрделі орталығы бар 4Fе-4S белок. Убиxинон (UQ) митоxоыдрияда жэне плазмалық мембранада орналасқан Элеісгрондарды қабьшдап, қайтымды тотықсызданады, гидроxинонға (убиxинолға) айналады, осындай түрінде тыныстану тізбегінде электрондар тасымалдау жүйесінде қызмет атқарады. Төменде убиxинонның тотығу жэне тотықсыздану реакциялары берілген.
. Гем емес темірі бар белрктарда темір атомдары белоктьің цистеин қалдықтарындағы күкірт атомдарымен байланысқан, сондықтан ондай белоктарды темір-күкіртті белоктар (Fе—S) деп те атайды. Олар тыныстану тізбегіндегі электрондарды тасымалдаудың бірнеше кезеңіне қатысады. Қазіргі уақыtта темір-күкіртті белоктардың митоxондрияның ішкі мембранасында орналасқан жері (12.2.сурет) жэне қайтымды тотығу-тотықсыздану реакцияларының меxанизмі анықталған. ФМH мен ФАДHг-ден Fе-S электрондарды қабылдап, убиxинонға тапсырады Суретте үш түрлі темір-күкіртті белоктардың сxемасы берілген сурет. Темір-күкіртті белок-тардың Fе- S орталық түрлері. А. Белок молекуласында темір атомы цистеиннің төрт қалдығымен жалғас-қан. Б. Белок екі темір атомы бейорганика-лық күкіртпен жэне төрт цистеин қалдықгарымен байланысқан. В. Күрделі орталығы бар 4Fе-4S белок. Убиxинон (UQ) митоxоыдрияда жэне плазмалық мембранада орналасқан. Элеісгрондарды қабьшдап, қайтымды тотықсызданады, гидроxинонға (убиxинолға) айналады, осындай түрінде тыныстану тізбегінде электрондар тасымалдау жүйесінде қызмет атқарады. Төменде убиxинонның тотығу жэне тотықсыздану реакциялары берілген.")
29
Убиздаион (UQ)тотық-қан түрі
Радикал Убихинол тотықсызданған (UQH2) түрі, гидрохинон Тьныстану тізбегінде убиxинон электрондары Fе-S-тен қабылдап, цитоxром h тапсырады (12.2.Сурет). Бұдан эрі қарай цитоxромдық жүйе электрон акцепторы болып табьшады. Цитоxромдар дегеніміз - қүрамында темір бар ферменттер тобы, олар электрондарды Q коферментінен молекулалық оттегіне тасымалдайды. Митоxондрияларда 20 шамасындай өзара үқсас цитоxромдар анықталған. Олардьщ ішінде эсірессе а, а3, Ь, с және с^ цитоxромдар жақсы зерттелген. Цитоxромдар -күрделі ферменттер, олардьщ қүрамында простетикалық топ ретінде гем бар. Гем сонымен қатар гемоглобин мен миоглобин құрамына да кіреді. Цитоxромдар бір-бірімен апоферменттердің қүрамымен, молекулалык массасьшен, гемнің бүйірлік тобының қүрьшымьшен жэне оптикалык қасиеттерімен айырылады. Ең кіші, М 12500, жануарлардың, ашытқыньщ митоxондриясынан бөліп алынған с цитоxром болады. Цитоxромоксядазаның қүрылымы анықталған, ол бірнеше суббөліктерден қүралған олигомерлік белок екен. Бактерия цитоxромоксидазасы 2-3 суббөліктен, ашытқьшьщ 7-8, жоғары сатылы жануарлардың цитоxромоксидазасы суббөліктен қүралған, М Активті орталығында екі гем және екі мыс ионы бар, Ь цитоxромның М 25000, сі Цитоxромдар эр түрлі сіңіру спектрымен сипатталады. Олардьщ а-алқап (полоса) толықын ұзындық мөлшері мынадай (нм): а цитоxром-600, а3-605, Ь -562, с-550 жэне Сі цитоxром-554. Цитоxромдардьщ қызметі - тыныстану тізбегінде электрондарды тасымалдау, ал сутек атомдарьш тасымалдамайды. Электрондарды тасымалдау кезінде цитоxром геміндегі темірдің валенттілігі қайтымды өзгереді. Тотыққан цитоxромда темір үш валентті (Fе3+), ал тотықсынданған цитохромда ол екі валентті (Fe2+) (Fe3+ Fe2+) Цитоxромдардың бір ерекшелігі - гемоглобин мен миоглобинде оттегін қосып алу жэне бөліп шығару кезінде темірдің валенттілігі өзгермейді, эрқашанда екі валентті күйінде қалады.Тыныстану тізбегінде бір мезгілде екі электрон беріліп отырады.Бірақ цитоxромньщ әр бір молекуласы бір мезгілде тек қана бір электронды алып-бере алады
 түрі, гидрохинон. Тьныстану тізбегінде убиxинон электрондары Fе-S-тен қабылдап, цитоxром h тапсырады (12.2.Сурет). Бұдан эрі қарай цитоxромдық жүйе электрон акцепторы болып табьшады. Цитоxромдар дегеніміз - қүрамында темір бар ферменттер тобы, олар электрондарды Q коферментінен молекулалық оттегіне тасымалдайды. Митоxондрияларда 20 шамасындай өзара үқсас цитоxромдар анықталған. Олардьщ ішінде эсірессе а, а3, Ь, с және с^ цитоxромдар жақсы зерттелген. Цитоxромдар -күрделі ферменттер, олардьщ қүрамында простетикалық топ ретінде гем бар. Гем сонымен қатар гемоглобин мен миоглобин құрамына да кіреді. Цитоxромдар бір-бірімен апоферменттердің қүрамымен, молекулалык массасьшен, гемнің бүйірлік тобының қүрьшымьшен жэне оптикалык қасиеттерімен айырылады. Ең кіші, М 12500, жануарлардың, ашытқыньщ митоxондриясынан бөліп алынған с цитоxром болады. Цитоxромоксядазаның қүрылымы анықталған, ол бірнеше суббөліктерден қүралған олигомерлік белок екен. Бактерия цитоxромоксидазасы 2-3 суббөліктен, ашытқьшьщ 7-8, жоғары сатылы жануарлардың цитоxромоксидазасы суббөліктен қүралған, М Активті орталығында екі гем және екі мыс ионы бар, Ь цитоxромның М 25000, сі Цитоxромдар эр түрлі сіңіру спектрымен сипатталады. Олардьщ а-алқап (полоса) толықын ұзындық мөлшері мынадай (нм): а цитоxром-600, а3-605, Ь -562, с-550 жэне Сі цитоxром-554. Цитоxромдардьщ қызметі - тыныстану тізбегінде электрондарды тасымалдау, ал сутек атомдарьш тасымалдамайды. Электрондарды тасымалдау кезінде цитоxром геміндегі темірдің валенттілігі қайтымды өзгереді. Тотыққан цитоxромда темір үш валентті (Fе3+), ал тотықсынданған цитохромда ол екі валентті (Fe2+) (Fe3+ Fe2+) Цитоxромдардың бір ерекшелігі - гемоглобин мен миоглобинде оттегін қосып алу жэне бөліп шығару кезінде темірдің валенттілігі өзгермейді, эрқашанда екі валентті күйінде қалады.Тыныстану тізбегінде бір мезгілде екі электрон беріліп отырады.Бірақ цитоxромньщ әр бір молекуласы бір мезгілде тек қана бір электронды алып-бере алады.")
30
Лекция №11 Тотыға фосфорлану
Бір мезгілде екі электронның тасымалдауын қамтамасыз ету үшін, тыныстану тізбегінің ішінде ерекше жоғары құрылыс болуы керек, ол бір уақытта екі электронды HАДH-дан немесе ФАДH2-ден альш, эрі қарай жібере алатындай. Тыныстану тізбегінде цитоxромдардың орналасу реті 12.4-суретте берілген. Тыныстану тізбегіндегі соңғы фермент а және а3 цитоxромымен байлаиысқан мыс бар, оның да валентгілігі злектрондар тасымалы кезінде өзгеріп отырады. Cu Cu2+ + -e а жэне аз комплексінің бірегей ерекшелігі мынадай: ол молекулалық оттегімен тікелей реакцияласады, сондықтан а жэне аз комплексін цито-xромоксидаза дзп атайды. Бүл комплексте мыс иондары өзінің электрондарын отгегіне береді. Басқа цитоxромдар молекулалы оттегшея өзара эрекеттесе алмайды. Молекулалық оттегі электрондарды қабылдайды да, анион түзіп, тотықсызданады: ½O2+2e O Тотықсызданған оттегі бос протондармен реакцияласады да, су түзеді. 12,4*2.Тыныстану тізбегінің төрт комплексі Тыныстану тізбегі өз бетімен жеке жүретін митоxондрияның бір бөліn. Бүтін митоxондрияны жұмсақ бөлшектеп зерттегенде, оның кейбір

34
Лекция №13

37
Лекция №14

42
Лекция №15

Παρόμοιες παρουσιάσεις











>")
















