Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
METABOLISMUL GLUCIDIC

2
INTRODUCERE Carbohidraţii: (CH2O)n sursă energetică formă de stocare a surplusului energetic comunicarea intercelulară rol structural: peretele bacterian

3
CLASIFICAREA CARBOHIDRAŢILOR
Monozaharidele pot fi clasificate astfel: Aldoze (-oza): exc. gliceraldehida Cetoze (-uloza): exc. dihidroxiacetona, fructoza, xiluloza Dizaharidele: 2 unităţi monozaharidice Oligozaharidele: unităţi monozaharidice Polizaharidele: > 10 unităţi monozaharidice (sute) ALDOZE CETOZE
: exc. gliceraldehida. Cetoze (-uloza): exc. dihidroxiacetona, fructoza, xiluloza. Dizaharidele: 2 unităţi monozaharidice. Oligozaharidele: unităţi monozaharidice. Polizaharidele: > 10 unităţi monozaharidice (sute) ALDOZE. CETOZE.")
4
IZOMERI ŞI EPIMERI Izomeri: compuşi cu formule chimice identice, dar cu structuri chimice diferite Ex. C6H12O6: fructoza, glucoza, galactoza şi manoza Epimeri: izomeri carbohidraţi care diferă prin configuraţia unui singur atom de C (exc. C din gruparea carbonil- anomeri); G şi Gal: epimeri la C4 DIZAHARID LACTOZA C1 C4 Galactoză legătură glicozidică Glucoză
; G şi Gal: epimeri la C4. DIZAHARID. LACTOZA. C1. C4. Galactoză. legătură. glicozidică. Glucoză.")
5
IZOMERI ŞI EPIMERI G şi Man: epimeri la C2
Gal şi Man nu sunt epimeri (structuri diferite la C2 şi C4); Gal şi Man sunt izomeri C2 Glucoză Manoză
; Gal şi Man sunt izomeri. C2. Glucoză. Manoză.")
6
ENANTIOMERI Glucidele din organism sunt D- glucide: gruparea -OH legată la C asimetric situat cel mai departe de C carbonilic este poziţionată dextrogir Racemazele asigură interconversia izomerilor D şi L ENANTIO- MERI D- GLUCOZA L- GLUCOZA

7
CICLIZAREA MONOZAHARIDELOR
Sub 1% din monozaharidele cu > 5 C au formă aciclică; majoritatea au configuraţie inelară prin interacţiunea dintre gruparea aldehidică (cetozică) cu un radical alcoolic al aceluiaşi glucid PIRANOZA: compus inelar hexa-atomic FURANOZA: compus inelar penta-atomic
 cu un radical alcoolic al aceluiaşi glucid. PIRANOZA: compus inelar. hexa-atomic. FURANOZA: compus inelar. penta-atomic.")
8
ATOMUL DE CARBON ANOMERIC
Ciclizarea crează un C anomeric (fostul C carbonilic) şi configuraţiile α şi β ale glucidului DIASTEREO- IZOMERI Gruparea OH a C anomeric de aceeaşi parte cu inelul
 şi configuraţiile α şi β ale glucidului. DIASTEREO- IZOMERI. Gruparea OH. a C anomeric de. aceeaşi parte cu. inelul.")
9
ATOMUL DE CARBON ANOMERIC
Enzimele deosebesc cele două structuri α şi β Glicogenul: sintetizat din α- D-glucopiranoză celuloza: sintetizată din β- D-glucopiranoză În soluţie, anomerii α şi β sunt în echilibru şi pot fi interschimbaţi spontan prin mutarotaţie

10
GLUCIDELE REDUCĂTOARE
Statusul atomului de oxigen legat de atomul anomeric dictează caracterul reducător sau nereducător al unui glucid, restul grupărilor OH nefiind implicate testele colorimetrice ce utilizează reactivi cromogeni detectează prezenţa glucidelor în urină identificarea exactă a glucidului necesită teste adiţionale specifice

11
Cuplarea monozaharidelor
Monozaharidele se pot uni pt a forma: Dizaharide: ○ lactoza Gal + G ○ sucroza G + Fr ○ maltoza G + G Polizaharide: ○ glicogen ramificat (orig. animală) ○ amidon (orig. vegetală) ○ celuloza neramificată (orig. animală) Polimeri de glucoză
 ○ amidon (orig. vegetală) ○ celuloza neramificată (orig. animală) Polimeri. de. glucoză.")
12
LEGĂTURILE GLICOZIDICE
Legăturile dintre glucide- punţi glicozidice în prezenţa glicoziltransferazelor (S = glucide nucleotidice de tipul UDP-G) Hidroliza unei legături glicozidice: glicozidaze Se denumesc în funcţie de: ○ nr. atomilor de C conectaţi ○ poziţia grupării OH anomerice: OH în configuraţie β- legătură glicozidică β Ex. lactoza
 Hidroliza unei legături glicozidice: glicozidaze. Se denumesc în funcţie de: ○ nr. atomilor de C conectaţi. ○ poziţia grupării OH anomerice: OH în configuraţie β- legătură glicozidică β. Ex. lactoza.")
13
LEGĂTURILE GLICOZIDICE
Lactoza are caracter reducător (capătul anomeric al G nefiind implicat în formarea legăturii glicozidice) Legătură galactozil- β (1->4 )- glucoză
 Legătură. galactozil- β (1->4 )- glucoză.")
14
CARBOHIDRAŢI COMPLECŞI
Carbohidraţii se pot ataşa prin legături glicozidice unor structuri neglucidice: ○ baze purinice şi pirimidinice ( acizi nucleici) ○ nuclee aromatice (steroizi, bilirubină) ○ proteinele (glicoproteine, glicozaminoglicani) ○ lipidele (glicolipide) Legătură N- glicozidică: glucid ataşat la -NH2 Legătură O- glicozidică: glucid ataşat la -OH Legăturile dintre glucide sunt O-glicozidice
 ○ nuclee aromatice (steroizi, bilirubină) ○ proteinele (glicoproteine, glicozaminoglicani) ○ lipidele (glicolipide) Legătură N- glicozidică: glucid ataşat la -NH2. Legătură O- glicozidică: glucid ataşat la -OH. Legăturile dintre glucide sunt O-glicozidice.")
15
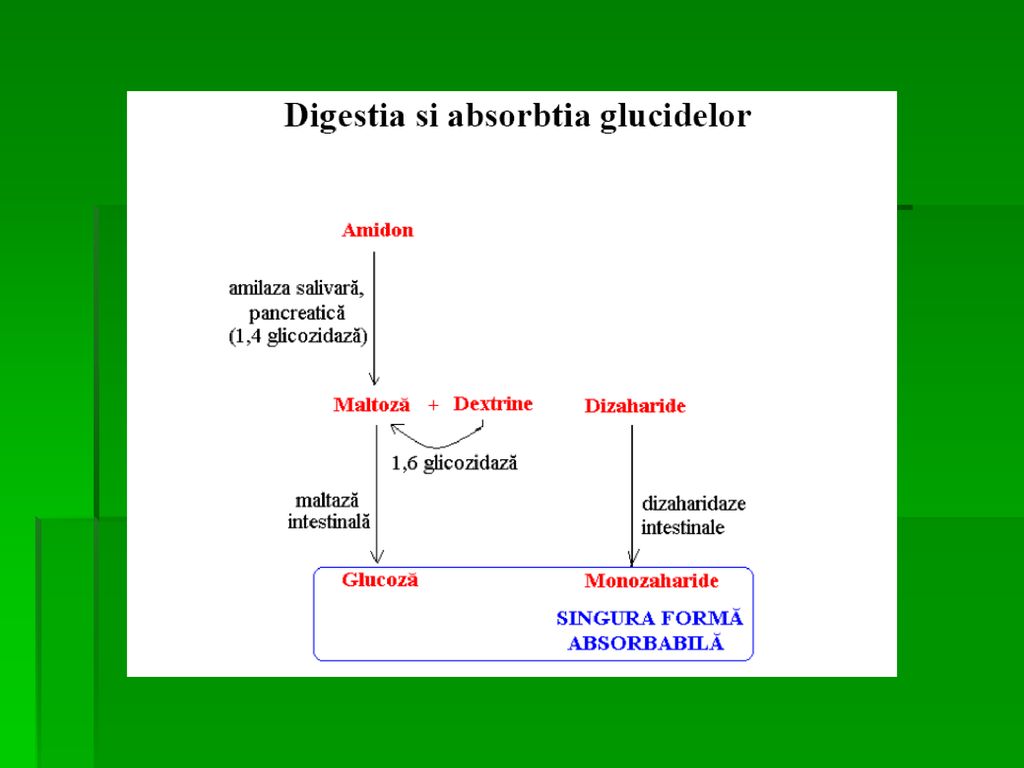
DIGESTIA CARBOHIDRAŢILOR
Digestia carbohidraţilor alimentari se desfăşoară în: ○ cavitatea orală ○ intestin ○ Procesul are loc rapid şi se încheie când chimul gastric ajunge la joncţiunea duodeno-jejunală Enzime: ○ Endoglicozidazele: hidroliza oligo- şi polizaharidelor ○ Dizaharidazele: hidroliza dizaharidelor ○ Glicozidazele specifice: hidroliza legăturilor glicozidice

17
DIGESTIA CARBOHIDRAŢILOR LA NIVELUL CAVITĂŢII ORALE
α- amilaza salivară hidrolizează legăturile α (1→4) din amidon si glicogen Celuloza nu poate fi digerată- legături β (1→4) α- amilaza salivară este inactivată de aciditatea gastrică locală GLICOGEN 1→4 1→6
 din amidon si glicogen. Celuloza nu poate fi digerată- legături β (1→4) α- amilaza salivară este inactivată de aciditatea gastrică locală. GLICOGEN. 1→4. 1→6.")
18
DIGESTIA CARBOHIDRAŢILOR LA NIVELUL CAVITĂŢII ORALE
Rezultă oligozaharide cu legături α (1→6) sau dextrine scurte, maltotrioze, dizaharide rezistente la actiunea α- amilazei
 sau dextrine scurte, maltotrioze, dizaharide rezistente la actiunea α- amilazei.")
19
DIGESTIA CARBOHIDRAŢILOR ÎN INTESTIN
Conţinutul gastric acid ajunge în lumenul intestinului subţire este neutralizat de bicarbonatul secretat de pancreas, iar α- amilaza pancreatică reia procesul de degradare a amidonului Etapele finale ale digestiei au loc în jejunul superior Enzime sintetizate la nivelul feţei luminale a marginii în perie a celulelor mucoasei intestinale Ex. Dizaharidaze şi oligozaharidaze: izomaltaza, maltaza, lactaza

20
DIGESTIA CARBOHIDRAŢILOR ÎN INTESTIN

21
ABSORŢIA MONOZAHARIDELOR
Absorţia glucidelor alimentare: în duoden şi jejun superior; insulina nu este necesară Mecanisme diferite de absorţie: ○ active 1.co-transportorul SGLT-1pt G şi Gal; 2. GLUT-5 pt Fr ○ G, Gal şi Fr sunt transportate din celulele mucoasei intestinale în circulaţia portală cu GLUT-2

22
DEGRADAREA ANORMALĂ A DIZAHARIDELOR
În mod normal, procesul global de digestie şi absorţie este eficient Disfuncţia dizaharidazelor specifice determină pătrunderea CH în lumenul intestinului gros cu producerea diarrei osmotice Fermentaţia bacteriană a CH cu 2 sau 3 atomi de C generează CO2 şi H2 activi cu apariţia crampelor abdominale, diareei, flatulenţei

23
DEFICIENŢELE ENZIMELOR DIGESTIVE CAUZE
Deficienţele congenitale ale dizaharidazelor: la sugari şi copii cu intoleranţă dizaharidică Malnutriţie, medicamente ce lezează mucoasa intestinală Deficienţă enzimatică temporară în diaree severă; nu se consumă lactate

24
DEFICIENŢELE ENZIMELOR DIGESTIVE CAUZE
Intoleranţa la lactoză > 3/4 din adulţi, 90% dintre adulţii africani şi asiatici au deficit de lactază reducere cantitativă a activităţii enzimatice Tratament: ○ reducerea consumului de lapte integral; ○ se preferă iaurt, branzeturi, broccoli pt Ca; ○ tablete cu lactază înainte de masă

25
DEFICIENŢELE ENZIMELOR DIGESTIVE
Deficienţa de izomaltază- sucrază determină intoleranţa la sucroză Tratament: ○ restricţia alimentelor cu zahăr ○ substituţia enzimatică Diagnostic: teste de toleranţă orală la fiecare dizaharid măsurarea H2 respirator

26
METABOLISM- NOŢIUNI INTRODUCTIVE
La nivel celular reacţiile enzimatice decurg sub forma căilor metabolice multisecvenţiale (calea glicolizei) Într-o cale metabolică, P unei reacţii = S al reacţiei următoare; căile metabolice se intersectează →reţea Metabolismul sau reţeaua integrată a reacţiilor chimice = suma tuturor modificărilor chimice la nivel celular
 Într-o cale metabolică, P unei reacţii = S al reacţiei următoare; căile metabolice se intersectează →reţea. Metabolismul sau reţeaua integrată a reacţiilor chimice = suma tuturor modificărilor chimice la nivel celular.")
27
METABOLISM- NOŢIUNI INTRODUCTIVE
Căile metabolice: ○ catabolice (de degradare) ○ anabolice (de sinteză) ○ Cicluri de reacţie = căile care regenerează un anumit compus chimic ○ Harta metabolică = reuneşte conexiunile dintre diversele căi, permite vizualizarea metaboliţilor intermediari şi a efectului blocării uneia dintre căi
 ○ anabolice (de sinteză) ○ Cicluri de reacţie = căile care regenerează un anumit compus chimic. ○ Harta metabolică = reuneşte conexiunile dintre diversele căi, permite vizualizarea metaboliţilor intermediari şi a efectului blocării uneia dintre căi.")
28
Glicoliza- cale metabolică

29
METABOLISM- NOŢIUNI INTRODUCTIVE
Căile catabolice asigură: captarea şi stocarea energiei chimice rezultate prin degradarea moleculelor de combustibil biologic sub formă de ATP conversia moleculelor alimentare şi a rezervelor intracelulare de nutrienţi în substanţe de bază necesare în sinteza moleculelor complexe căi oxidative ce necesită coenzime, NAD+ 3 etape

30
METABOLISM- NOŢIUNI INTRODUCTIVE
1. Hidroliza moleculelor complexe în unităţile componente 2. Conversia unităţilor componente în acetil-CoA 3. Oxidarea acetil-CoA; fosforilarea oxidativă Catabolismul-proces divergent, obţinându-se numeroşi produşi complecşi ciclul acidului citric

31
METABOLISM- NOŢIUNI INTRODUCTIVE
Căile anabolice: asigură combinarea moleculelor mici pt a forma molecule complexe sunt endergonice, ATP fiind descompus la ADP şi Pi implică reduceri chimice, NADPH, o moleculă care cedează electroni Anabolismul- proces convergent, în final se obţine un nr redus de produşi

32
REGLAREA METABOLISMULUI
Producerea de energie şi sinteza produşilor finali corespund necesităţilor celulare Funcţiile organismului sunt coordonate printr-un sistem de comunicare elaborat Semnalele reglatoare ce informează fiecare celulă asupra statusului metabolic al organismului includ: ○ hormoni ○ neurotransmiţători ○ cantitatea de nutrienţi Semnalele reglatoare influenţează semnalele intracelulare

33
REGLAREA METABOLISMULUI
Semnalele intracelulare reglatoare pot controla viteza unei căi metabolice ce depinde de: ○ cantitatea de substrat disponibilă ○ inhibarea produsului de reacţie ○ modificări ale activatorilor şi inhibitorilor alosterici Comunicarea intercelulară= răspunsul la semnalele extracelulare esenţial în supravieţuire şi dezvoltare Determină un răspuns mai lent comparativ cu semnalizarea intracelulară fiind realizată prin: ○ contactul suprafeţelor ○ joncţiuni comunicante între citoplasmele adiacente

34
SEMNALIZARE INTERCELULARĂ SINAPTICĂ
CELULĂ NERVOASĂ NEUROTRANSMIŢĂTOR Ach CELULĂ ŢINTĂ Mecanism sinaptic

35
SEMNALIZARE INTERCELULARĂ ENDOCRINĂ ŞI PRIN CONTACT DIRECT
Mecanism endocrin Mecanism prin contact direct

36
SISTEMUL MESAGERILOR SECUNZI
Pentru metabolismul energetic cea mai importantă este semnalizarea chimică dintre celule prin: ○ hormoni ○ neurotransmiţători Receptorii acestora= detectori de semnale Fiecare componentă realizează legătura cu una dintre modificările chimice extra- şi intracelulare Mesagerii secunzi asigură legătura dintre: ○ mesagerul primar ○ răspunsul celular Mesageri primari

37
SISTEMUL MESAGERILOR SECUNZI
Sistemul calciu/ fosfatidil- inozitol Sistemul adenilat-ciclazei Adenilat ciclaza (AC): activitatea ei este crescută/ scăzută prin recunoaşterea semnalului chimic de R β şi α2 adrenergici Toxina produsă de Vibrio cholerae + AC; toxina produsă de Bordetella pertussis - AC Enzimă membranară
: activitatea ei este crescută/ scăzută prin recunoaşterea semnalului chimic de R β şi α2 adrenergici. Toxina produsă de Vibrio cholerae + AC; toxina produsă de Bordetella pertussis - AC. Enzimă membranară.")
38
SISTEMUL MESAGERILOR SECUNZI
1. Proteinele reglatoare GTP- dependente= proteine membranare specializate care leaga GDP sau GTP Heterotrimere: α (contine situsul de legarea al GDP), β, γ GDP-αβγ= forma inactiva a proteinei; cH-R actioneaza asupra pG inactive pe care o activează GTP- α se deplasează de pe R pe AC-ază activând-o.
, β, γ. GDP-αβγ= forma inactiva a proteinei; cH-R actioneaza asupra pG inactive pe care o activează. GTP- α se deplasează de pe R pe AC-ază activând-o.")
39
SISTEMUL MESAGERILOR SECUNZI
1 R activat conduce la formarea mai multor molecule de proteină G activă proteinele Gs stimulează AC-aza proteinele Gi inhibă enzima Efectele proteinei G- GTP sunt de scurtă durată deoarece proteina G are activitate GTP-azică cu formarea de GDP şi inactivarea proteinei G

40
SISTEMUL MESAGERILOR SECUNZI
Protein-kinazele AMPc dependente PKA este o holoenzima alcatuita din dimerul subunitatii reglatoare (R), având fiecare monomer legat la o subunitatea catalitica. La concentratii mici de cAMP, holoenzima rămâne intactă şi inactiva catalitic Când [cAMP] creste (activarea AC-azei de R cuplati proteinei G atasati la Gs, inhibarea PDE care degradeaza cAMP), cAMP se fixeaza la cele doua situsuri de legare de pe subunitatile reglatoare, cu eliberarea subunitatilor catalitice
, având fiecare monomer legat la o subunitatea catalitica. La concentratii mici de cAMP, holoenzima rămâne intactă şi inactiva catalitic. Când [cAMP] creste (activarea AC-azei de R cuplati proteinei G atasati la Gs, inhibarea PDE care degradeaza cAMP), cAMP se fixeaza la cele doua situsuri de legare de pe subunitatile reglatoare, cu eliberarea subunitatilor catalitice.")
41
SISTEMUL MESAGERILOR SECUNZI
AMPc activează pkA prin legare la subunităţile reglatoare R, cu eliberarea celor catalitice C. Acestea catalizează transferul ionului fosfat de pe ATP pe Ser sau Thr din substraturile proteice. Fosfoproteinele fie acţionează direct la nivelul canalelor ionice, ale celulei fie se transformă în enzime activate sau inhibate

42
SISTEMUL MESAGERILOR SECUNZI
pK-azele fosforilează proteinele specifice ce aderă la regiunile promotoare ale ADN cu creşterea expresiei unor gene specifice Nu toate pK-azele răspund la prezenţa AMPc. Ex pK C Defosforilarea proteinelor se realizează cu ajutorul protein-fosfatazelor enzime ce produc clivarea hidrolitică a esterilor fosfat Ca urmare, modificările enzimatice induse de fosforilarea proteinelor sunt temporare.

43
SISTEMUL MESAGERILOR SECUNZI
Hidroliza AMPc la 5'- AMP sub acţiunea AMPc-fosfodiesterazei (PDE) ce scindează legăturile 3'5'- fosfodiesterice; Molecula de 5'- AMP = semnal intracelular
 ce scindează legăturile fosfodiesterice; Molecula de 5 - AMP = semnal intracelular.")
44
SISTEMUL MESAGERILOR SECUNZI
PDE este inhibată de : ○ derivaţi metilxantinici (teofilina, cafeina) Efectele creşterii [cAMP] sub acţiunea neurotransmiţătorilor şi hormonilor sunt rapid epuizate prin îndepărtarea stimulului extracelular Inactivarea pKA prin mecanism feedback: PDE, unul dintre substratele activate de kinaza, converteşte cAMP la AMP reducând cantitatea de cAMP care ar putea activa protein kinaza A Astfel, pKA este controlata de cAMP. Subunitatea catalitică este inactivată prin fosforilare.
 Efectele creşterii [cAMP] sub acţiunea neurotransmiţătorilor şi hormonilor sunt rapid epuizate prin îndepărtarea stimulului extracelular. Inactivarea pKA prin mecanism feedback: PDE, unul dintre substratele activate de kinaza, converteşte cAMP la AMP reducând cantitatea de cAMP care ar putea activa protein kinaza A. Astfel, pKA este controlata de cAMP. Subunitatea catalitică este inactivată prin fosforilare.")
48
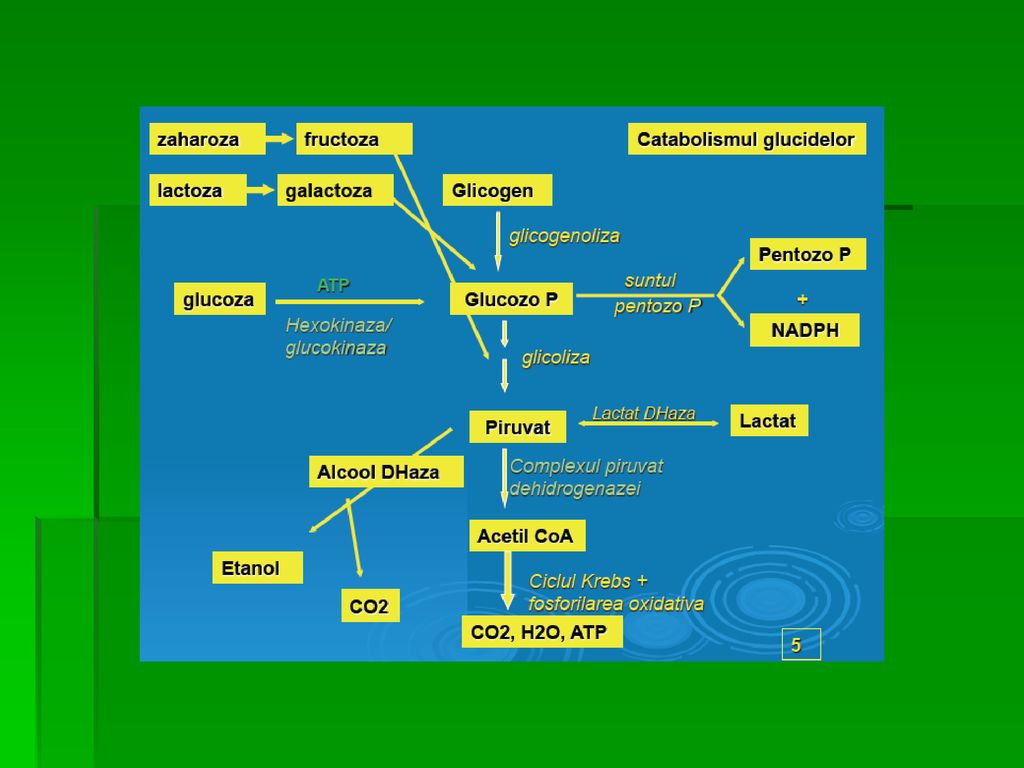
GLICOLIZA

49
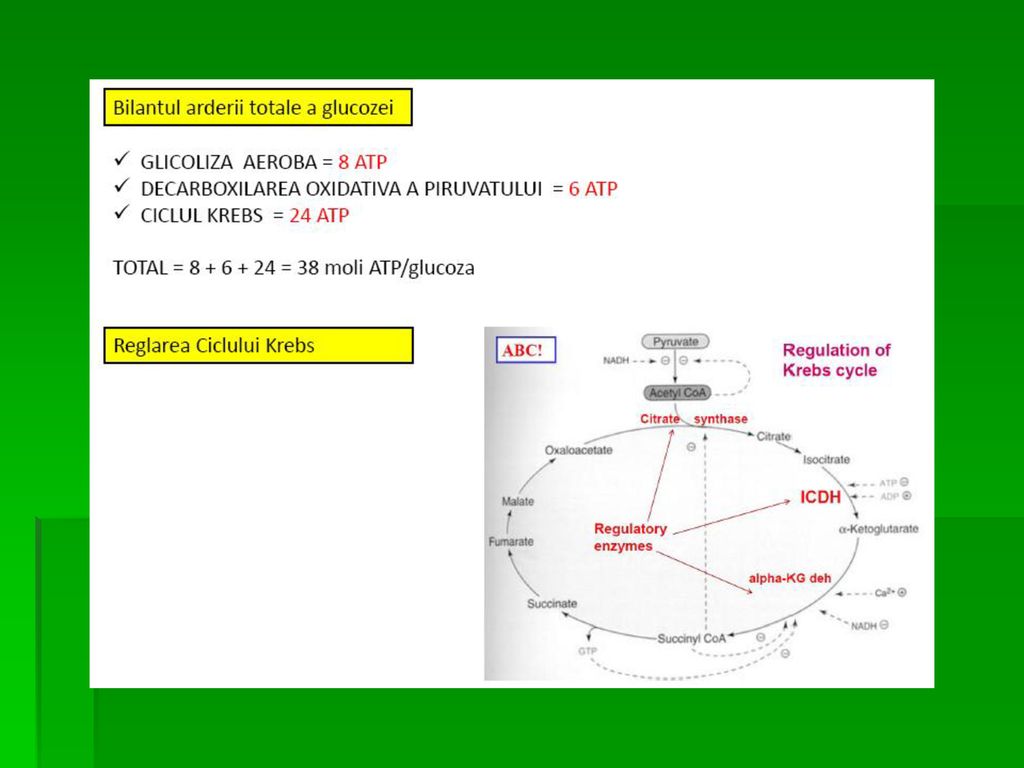
GLICOLIZA Definiţie. Degradarea glucozei cu producere de ATP ( 2 molecule ATP/ 1mol Glucoză ) şi metaboliţi intermediari ce pot fi utilizaţi în alte căi metabolice. Are loc în citosol. În centrul metabolismului CH În aerobioză (reoxidarea NADH este indispensabilă): Piruvatul este convertit la acetil- CoA,prin oxidarea la nivelul ciclului citric şi a lanţului respirator mitocondrial rezultând moli ATP/mol G 10 REACŢII
 şi metaboliţi intermediari ce pot fi utilizaţi în alte căi metabolice. Are loc în citosol. În centrul metabolismului CH. În aerobioză (reoxidarea NADH este indispensabilă): Piruvatul este convertit la acetil- CoA,prin oxidarea la nivelul ciclului citric şi a lanţului respirator mitocondrial rezultând moli ATP/mol G. 10 REACŢII.")
50
GLICOLIZA Alternativ, Py este redus la lactat (NADH este oxidată la NAD +. Glicoliza anaerobă permite sinteza de ATP în ţesuturile în care aportul de oxigen este insuficient: hematii ce nu conţin mitocondrii, leucocite, cornee, cristalin, celule în hipoxie-muşchi scheletic în contracţie rapidă

51
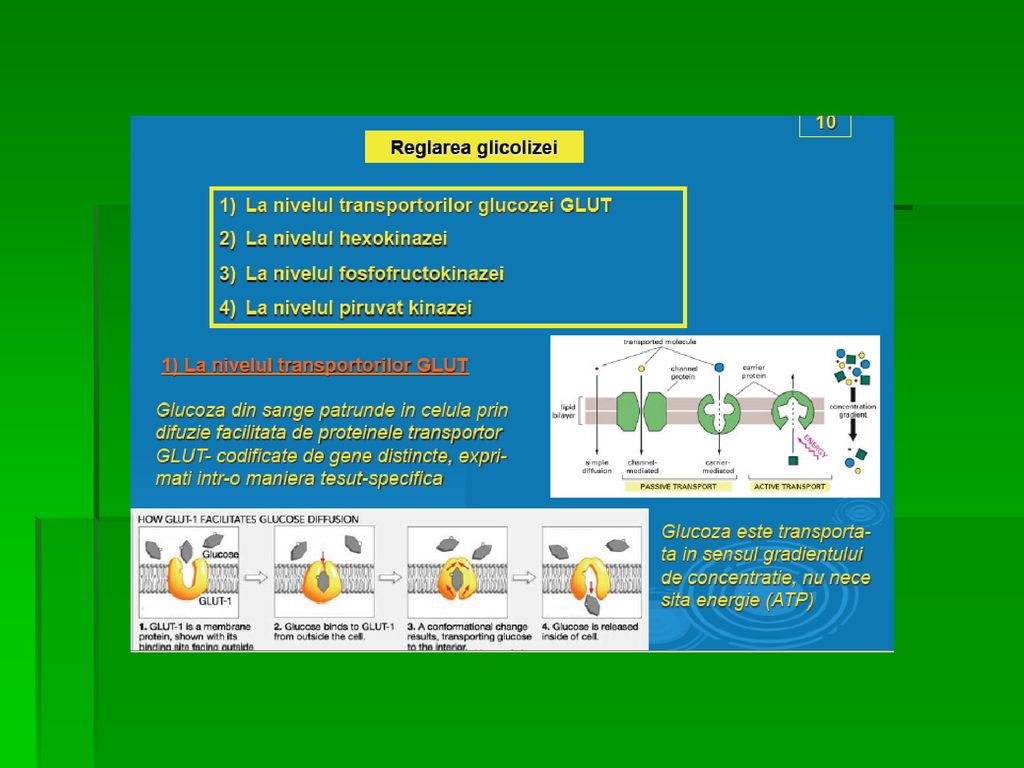
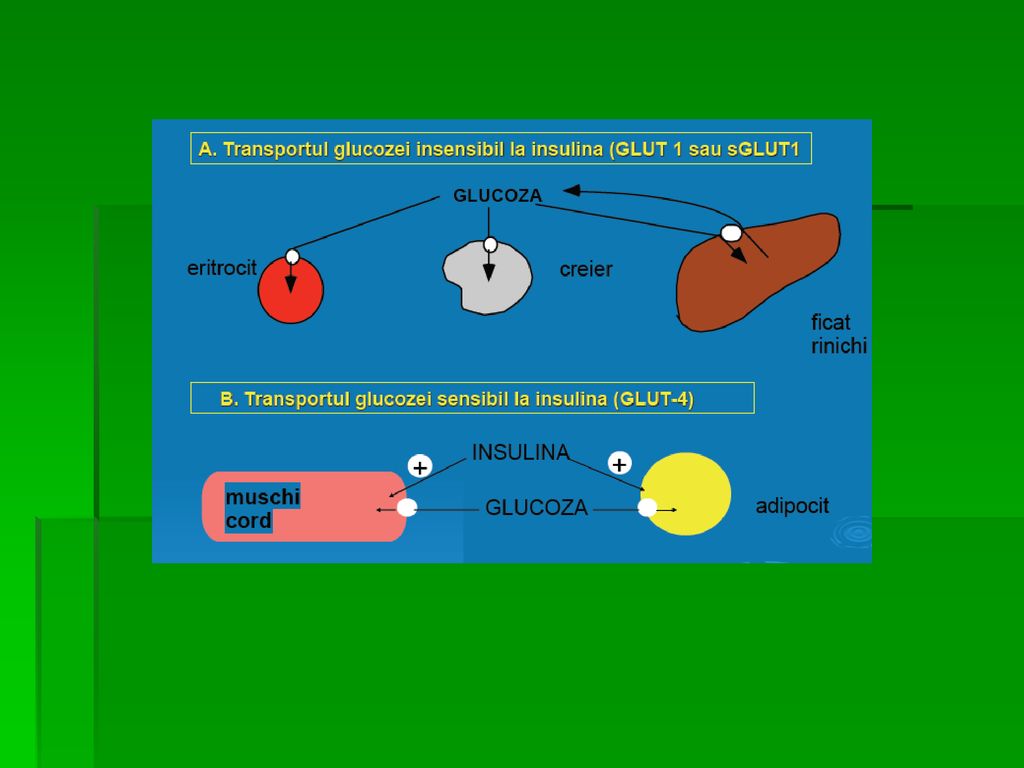
TRANSPORTUL GLUCOZEI ÎN CELULE
Glucoza nu poate pătrunde în celule prin difuziune simplă 2 mecanisme: transport facilitat şi cotransport Difuziunea facilitată (Na +- independentă) realizată cu 14 proteine transportoare membranare (izoforme ale aceluiaşi transportor): GLUT 1-14 ce prezintă secvenţe comune şi specifice membranei rezidente G extracelulară se leagă la transportor care suferă o modificare conformaţională ce permite transportul G prin MC GLUT- Glucose Transporter
 realizată cu 14 proteine transportoare membranare (izoforme ale aceluiaşi transportor): GLUT 1-14 ce prezintă secvenţe comune şi specifice membranei rezidente. G extracelulară se leagă la. transportor care suferă o modificare. conformaţională ce permite. transportul G prin MC. GLUT- Glucose Transporter.")
52
TRANSPORTUL GLUCOZEI ÎN CELULE
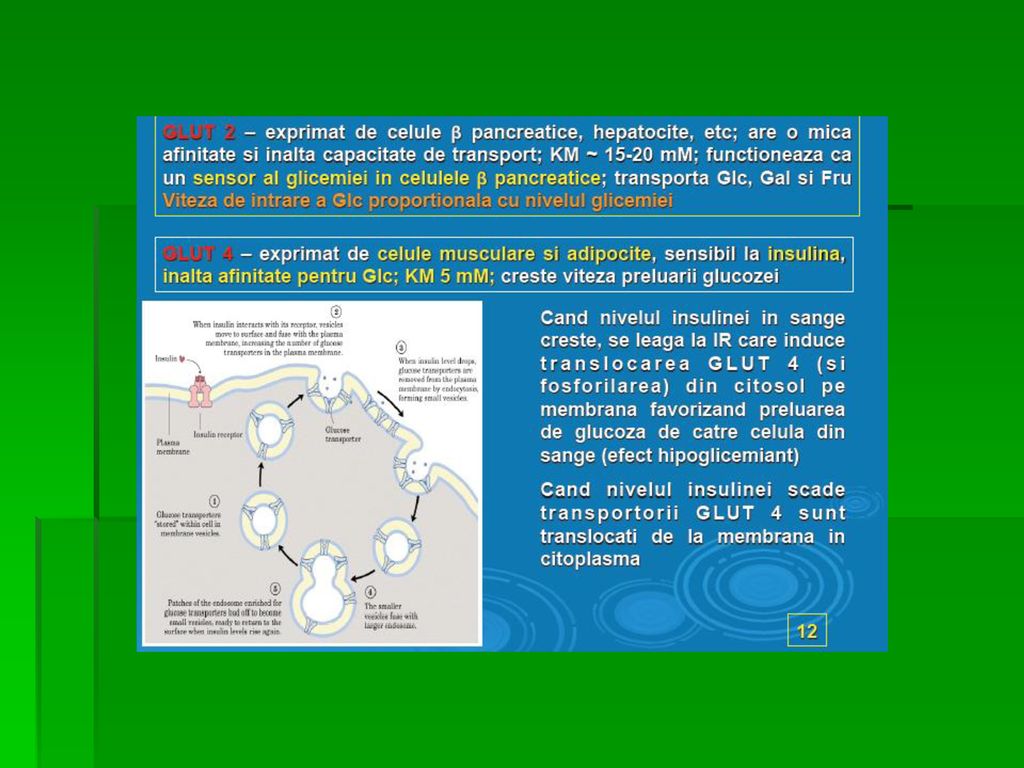
Specificitatea tisulară a expresiei genei GLUT: GLUT-3 principalul transportor la nivel neuronal GLUT-1 în eritrocite şi ţesutul cerebral GLUT-4 în adipocite şi celulele musculare striate, nr şi activitatea lor fiind crescute de insulină Funcţiile specializate ale izoformelor GLUT: În difuziunea facilitată G este transportată din mediul cu [G] mai mare spre cel cu [G] mai mică GLUT-1, GLUT-3 şi GLUT-4 implicate în preluarea G din circulaţie

53
TRANSPORTUL GLUCOZEI ÎN CELULE
Funcţiile specializate ale izoformelor GLUT-2 este localizată în ficat şi rinichi şi transportă G în interiorul hepatocitelor şi al celulelor renale când glicemia este crescută sau invers (din aceste celule în sânge) la glicemie mică ( à jeun); este prezent şi în pancreas GLUT-5: în trasnportul primar al Fr (intestin subţire, testicule) GLUT-7: în ficat şi alte ţesuturi gluconeogenetice mediază fluxul G prin membrana RE
 la glicemie mică ( à jeun); este prezent şi în pancreas. GLUT-5: în trasnportul primar al Fr (intestin subţire, testicule) GLUT-7: în ficat şi alte ţesuturi gluconeogenetice mediază fluxul G prin membrana RE.")
54
TRANSPORTUL GLUCOZEI ÎN CELULE
Sistemul de co-transport Na + - glucoză necesită consum de energie şi asigură transportul G împotriva gradientului de concentraţie (extracelular [G] este < decât intracelular). La nivelul transportorului sunt ataşaţi G şi Na + . Ionii de sodiu sunt transportaţi în celulă simultan cu G, proces favorizat de energia conţinută în gradientul de Na. Transportorul este sodiu-dependent sau SGLT În celulele epiteliului intestinal, tubilor renali şi ale plexului coroid
. La nivelul transportorului sunt ataşaţi G şi Na + . Ionii de sodiu sunt transportaţi în celulă simultan cu G, proces favorizat de energia conţinută în gradientul de Na. Transportorul este sodiu-dependent sau SGLT. În celulele epiteliului intestinal, tubilor renali şi ale plexului coroid.")
55
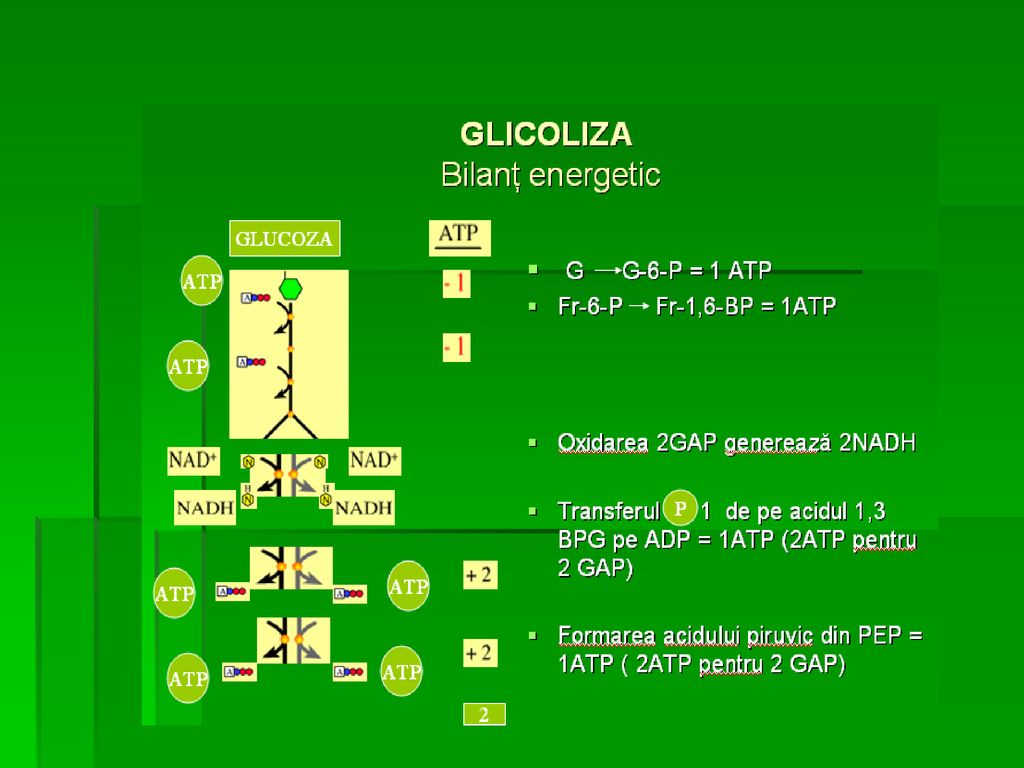
REACŢIILE GLICOLIZEI Conversia G la Py decurge în 2 etape:
Prima, consumatoare de 2ATP/mol de G, hexozele fosforilate fiind transformate într-un produs comun: gliceraldehid-3-fosfat (GAP) A II-a, de oxidare a GAP, generatoare de 4 ATP/mol de glucoză, 2Py şi 2 NADH, H+, ca urmare a oxidării GAP Cele două molecule de NADH sunt oxidate la NAD+ pe măsură ce Py este redus la lactat (glicoliză anaerobă)
 A II-a, de oxidare a GAP, generatoare de 4 ATP/mol de glucoză, 2Py şi 2 NADH, H+, ca urmare a oxidării GAP. Cele două molecule de NADH sunt oxidate la NAD+ pe măsură ce Py este redus la lactat (glicoliză anaerobă)")
56
REACŢIILE GLICOLIZEI Toţi intermediarii glicolizei (de la G la Py) sunt fosforilaţi Gruparea fosfat a intermediarilor glicolizei are 3 roluri principale: 1. de a-i menţine în celulă (cu un grup polar negativ sunt incapabili să traverseze membrana celulară la pH=7, prin simpla difuziune) 2. păstrarea moleculelor energetice în celulă (compuşi hidrofili) 3. interacţiunea enzimă-substrat mai bună
 2. păstrarea moleculelor energetice în celulă (compuşi hidrofili) 3. interacţiunea enzimă-substrat mai bună.")
57
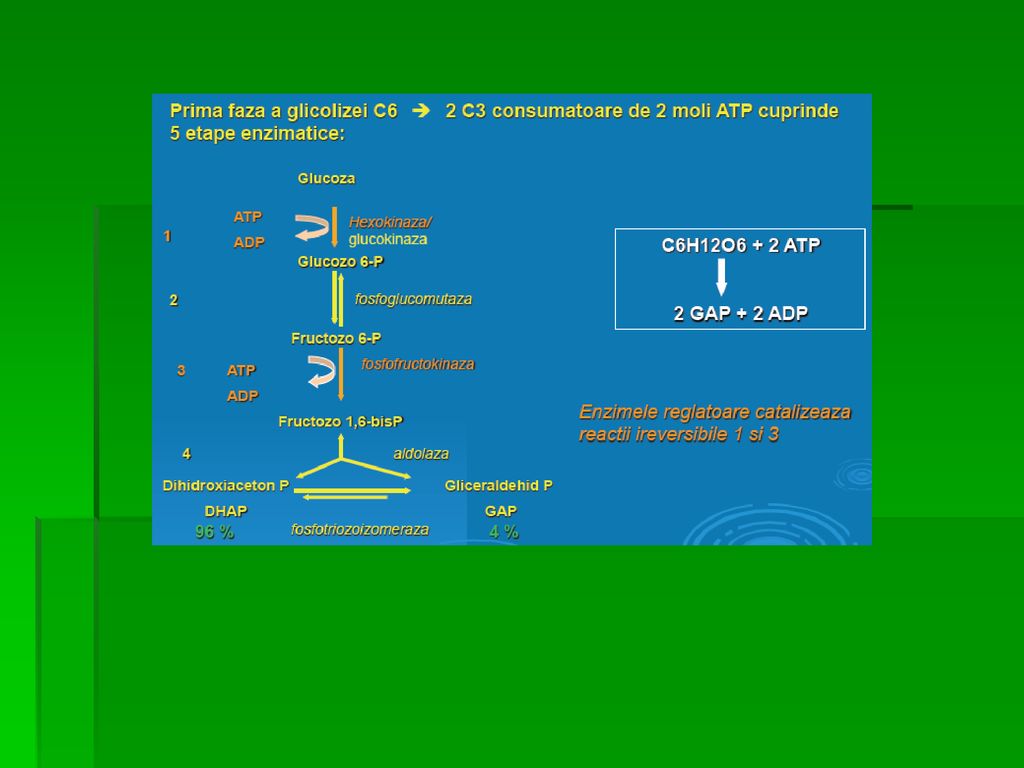
REACŢIILE GLICOLIZEI Primele 5 reacţii: A. Fosforilarea glucozei
B. Izomerizarea G-6-P la Fr-6-P (fructozo-6-fosfat) C. Fosforilarea Fr-6-P la Fr-1,6- bisfosfat D. Scindarea Fr-1,6- difosfatului E. Izomerizarea dihidroxiaceton-fosfatului (Interconversiunea trioz-fosfaţilor)
 C. Fosforilarea Fr-6-P la Fr-1,6- bisfosfat. D. Scindarea Fr-1,6- difosfatului. E. Izomerizarea dihidroxiaceton-fosfatului (Interconversiunea trioz-fosfaţilor)")
58
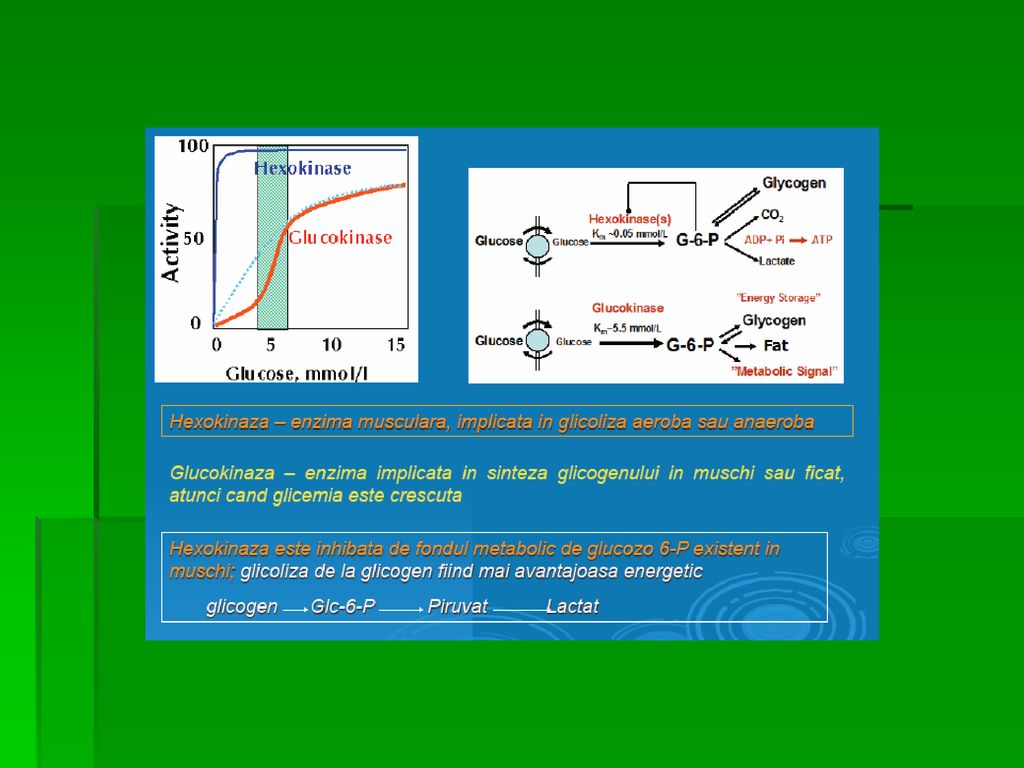
REACŢIILE GLICOLIZEI A. Fosforilarea glucozei este catalizată de izoenzimele hexokinazei ( una dintre cele 3 enzime reglatoare ale glicolizei alături de fosfofructokinază şi piruvat-kinază) Hexokinaza: ○ Sp largă pt S fosforilând mai multe hexoze înafara G ○ inhibată de produsul final de reacţie G-6-fosfatul care se acumulează în organism când rata de metabolizare scade ○ Km scăzut pt G ○ Vmax redus pt G ( nu poate reţine hexozele fosforilate în interiorul celulei şi nici nu poate fosforila o cantitate de glucide mai mare decât necesarul
 Hexokinaza: ○ Sp largă pt S fosforilând mai multe hexoze înafara G. ○ inhibată de produsul final de reacţie G-6-fosfatul care se acumulează în organism când rata de metabolizare scade. ○ Km scăzut pt G. ○ Vmax redus pt G ( nu poate reţine hexozele fosforilate în interiorul celulei şi nici nu poate fosforila o cantitate de glucide mai mare decât necesarul.")
59
REACŢIILE GLICOLIZEI Reacţia de fosforilare este ireversibilă şi necesită 1mol ATP Mg este indispensabil în toate fosforilările Glucokinaza sau hexokinaza D sau tipul IV ○ principala enzimă care asigură fosforilarea G ○ în celulele β pancreatice, detector de G, determinând pragul secreţiei de insulină ○ la nivel hepatic facilitează fosforilarea G în episoadele hiperglicemice ○ Sp de S similară izoenzimelor hexokinazei

60
REACŢIILE GLICOLIZEI Km pt glucokinază >>> Km pt hexokinaza
Glucokinaza funcţionează la [G] hepatocitare (postprandial) Vmax pt glucokinaza >>> ○ permite Fi să elimine eficient fluxul masiv de G adus prin sângele portal ○ previne supraâncărcarea cu G a circulaţiei sistemice ○ reduce Hglicemia din per. de absorţie (GLUT-2)
 Vmax pt glucokinaza >>> ○ permite Fi să elimine eficient fluxul masiv de G adus prin sângele portal. ○ previne supraâncărcarea cu G a circulaţiei sistemice. ○ reduce Hglicemia din per. de absorţie (GLUT-2)")
61
REACŢIILE GLICOLIZEI Activitatea glucokinazei: specific
○ nu este inhibată alosteric de G-6-P (ca pt alte hexokinaze); ○ este inhibată indirect de Fr-6-fosfat (în echilibru cu G-6-P) ○ stimulată indirect de G printr-un mecanism specific
; ○ este inhibată indirect de Fr-6-fosfat (în echilibru cu G-6-P) ○ stimulată indirect de G printr-un mecanism. specific.")
62
REACŢIILE GLICOLIZEI Nucleul hepatocitelor conţine o proteină reglatoare a glucokinazei În prezenţa Fr-6-P, GK este translocată în nucleu şi se leagă strâns de proteina reglatoare (GRP), inactivând enzima când glicemia este crescută (GLUT-2 contribuie la creşterea [G] hepatocitare, G determină separarea GK de GRP cu pătrunderea enzimei în citosol, având loc fosforilarea G la G-6-P Când [G] libere scade, Fr-6-P determină translocarea GK înapoi în nucleul hepatocitar şi legarea acesteia la GRP cu inhibarea activităţii enzimatice; GK=senzor de G; creşte secreţia de insulină metabolizarea G
, inactivând enzima. când glicemia este crescută (GLUT-2 contribuie la creşterea [G] hepatocitare, G determină separarea GK de GRP cu pătrunderea enzimei în citosol, având loc fosforilarea G la G-6-P. Când [G] libere scade, Fr-6-P determină translocarea GK înapoi în nucleul hepatocitar şi legarea acesteia la GRP cu inhibarea activităţii enzimatice; GK=senzor de G; creşte secreţia de insulină metabolizarea G.")
63
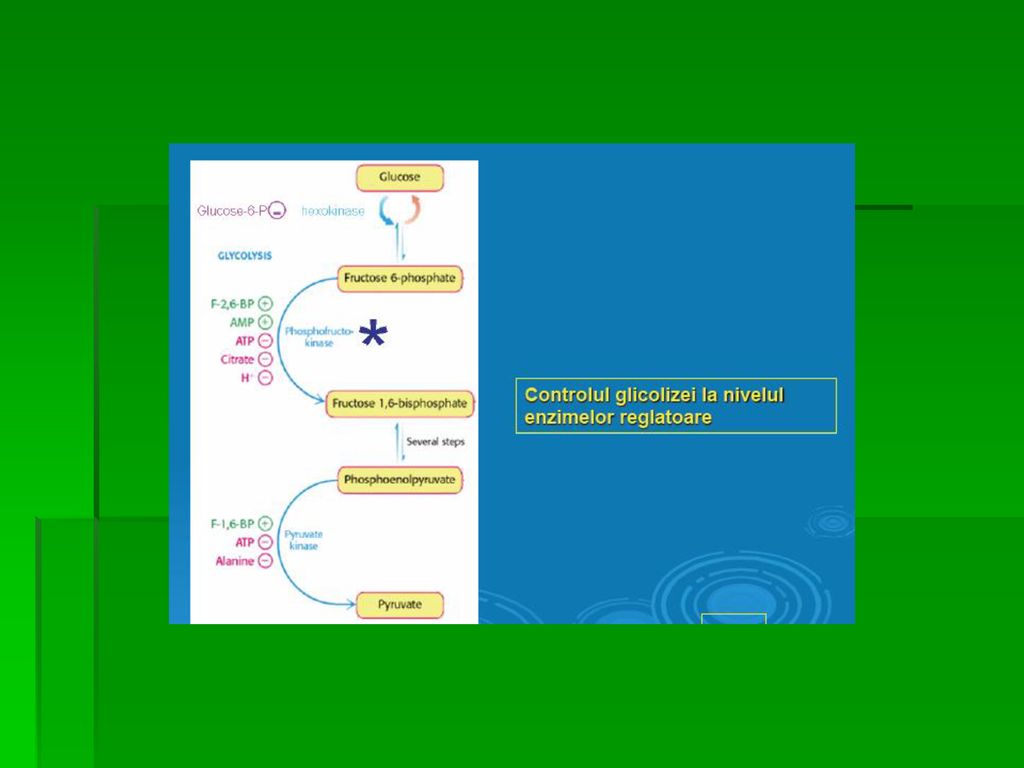
REACŢIILE GLICOLIZEI B. Izomerizarea G-6-P la Fr-6-P (fructozo-6-fosfat) Reacţia este complet reversibilă Nu constituie o etapă limitantă de viteză Are loc în prezenţa fosfoglucoizomerazei

64
REACŢIILE GLICOLIZEI C. Fosforilarea Fr-6-P la Fr-1,6- bisfosfat are loc în prezenţa Fosfofructokinazei I (PFK-1) sau a fructozo-6- fosfat kinazei Cea mai importantă etapă limitantă de viteză a glicolizei PFK-1 este o enzimă alosterică, “cheie”, inhibată de prezenţa unor cantităţi mari de ATP (semnal energogen) şi de Fr-6-P modularea activităţii PFK-1 de alte substanţe reglatoare
 şi de Fr-6-P. modularea activităţii PFK-1 de alte substanţe reglatoare.")
65
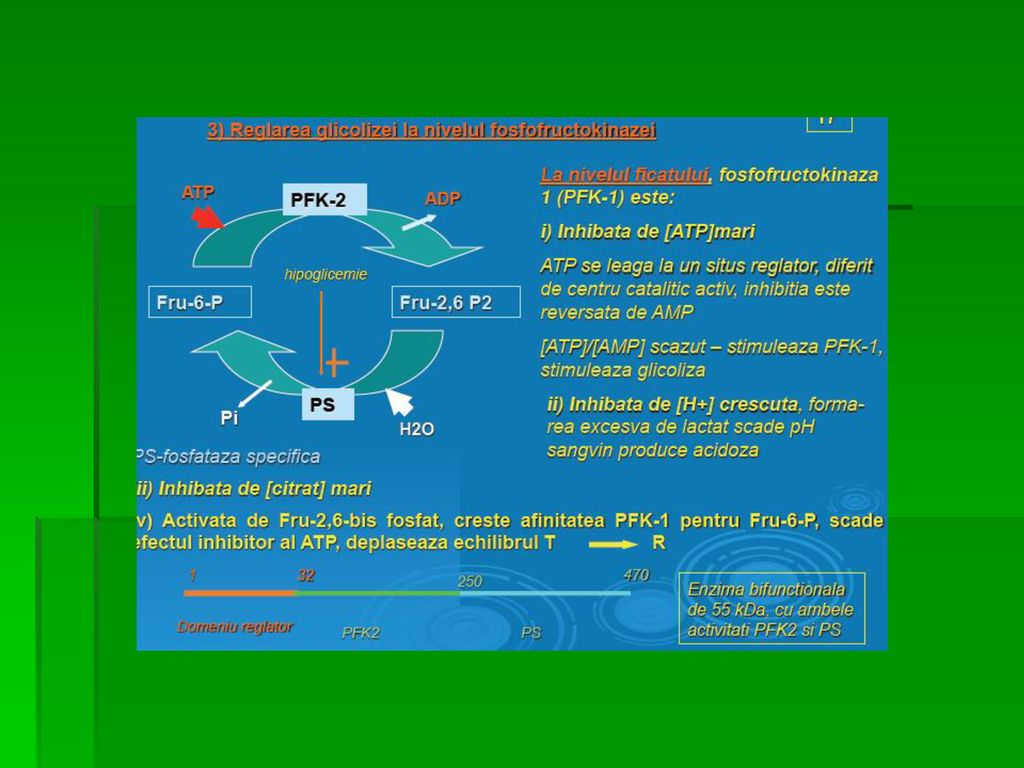
REACŢIILE GLICOLIZEI Reglarea realizată de rezervele energetice icel.
PFK-1 este inhibată de: ○ [ATP] crescută ○ [citrat] crescută PFK-1 este inhibată de [AMP] crescută, indicând epuizarea rezervelor energetice celulare Reglarea exercitată de [Fr-2,6-difosfat] Fr-2,6-difosfat= cel mai puternic activator al PFK-1 activând-o şi în prezenţa unor cantităţi crescute de ATP

66
REACŢIILE GLICOLIZEI Reglarea exercitată de [Fr-2,6-difosfat]
○ Fr-2,6-difosfat se formează sub acţiunea PFK-2, diferită de PFK-1 cu dublă activitate: kinazică şi fosfatazică ce asigură retroconversia Fr-2,6-difosfat la Fr-6-P ○ În ficat, domeniul kinazic este activ când este defosforilat ○ Fr-2,6-difosfat inhibă Fr-1,6-disfosfataza, enzima GNG
![REACŢIILE GLICOLIZEI Reglarea exercitată de [Fr-2,6-difosfat]](http://slideplayer.gr/slide/14451298/90/images/66/REAC%C5%A2IILE+GLICOLIZEI+Reglarea+exercitat%C4%83+de+%5BFr-2%2C6-difosfat%5D.jpg "○ Fr-2,6-difosfat se formează sub acţiunea PFK-2, diferită de PFK-1 cu dublă activitate: kinazică şi fosfatazică ce asigură retroconversia Fr-2,6-difosfat la Fr-6-P. ○ În ficat, domeniul kinazic este activ când este defosforilat. ○ Fr-2,6-difosfat inhibă Fr-1,6-disfosfataza, enzima GNG.")
67
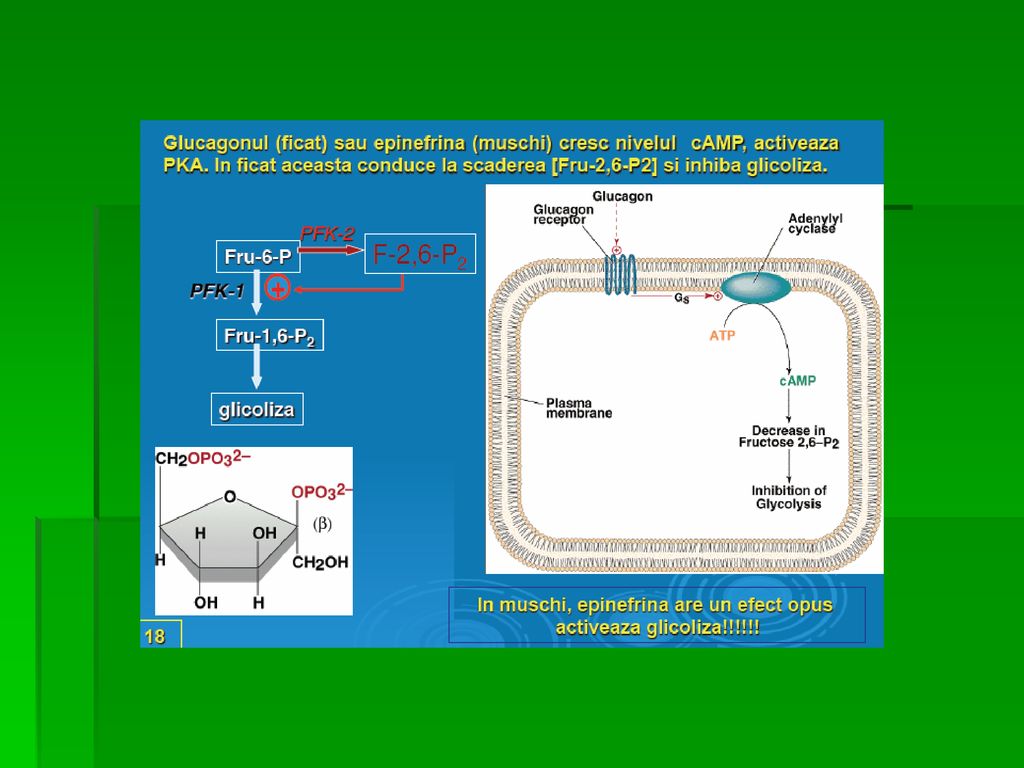
REACŢIILE GLICOLIZEI Acţiunea reciprocă a Fr-2,6-bisfosfat de activare a glicolizei şi de inhibare a GNG (nefiind simultane cele două căi) este împiedicată crearea unui ciclu inutil prin care G ar fi transformată în Py în paralel cu resinteza G din Py postprandial, [Glucagon] şi [insulină] stimulează activitatea Fr-2,6-bisfosfat, cu activarea glicolizei Fr-2,6-bisfosfat = semnal icel ce indică prezenţa unei [G] à jeun, [Glucagon] şi [insulină] reduce [Fr-2,6-bisfosfat] intrahepatocitar cu scăderea ratei globale a glicolizei
 este împiedicată crearea unui ciclu inutil prin care G ar fi transformată în Py în paralel cu resinteza G din Py. postprandial, [Glucagon] şi [insulină] stimulează activitatea Fr-2,6-bisfosfat, cu activarea glicolizei. Fr-2,6-bisfosfat = semnal icel ce indică prezenţa unei [G] à jeun, [Glucagon] şi [insulină] reduce [Fr-2,6-bisfosfat] intrahepatocitar cu scăderea ratei globale a glicolizei.")
68
REACŢIILE GLICOLIZEI D. Scindarea Fr-1,6- difosfatului în prezenţa aldolazei (Fr-1,6- bisfosfat-aldolaza) reacţie reversibilă şi necontrolată În ficat şi rinichi, aldolaza B clivează Fr-1,6- difosfatul, intervenind în metabolizarea Fr alimentare În urma clivării Fr-1,6- bisfosfat-ului se obţine DHAP (dihidroxiacetonfosfat) şi GAP (gliceraldehid-3-fosfat)
 şi GAP. (gliceraldehid-3-fosfat)")
69
REACŢIILE GLICOLIZEI E. Izomerizarea dihidroxiaceton-fosfatului (Interconversiunea trioz-fosfaţilor) Numai GAP este degradat în faza a II-a a glicolizei DHAP este utilizat după conversia la GAP Reacţia este reversibilă şi catalizată de triozo-fosfat-izomeraza avâd ca rezultat net producerea a 2 molecule de gliceraldehid-3-fosfat din produşii de clivare ai Fr-1,6-difosfatului Se încheie prima etapă a glicolizei

70
REACŢIILE GLICOLIZEI Etapa producătoare de energie- 5 etape:
F. Oxidarea gliceraldehid-3-fosfat (GAP) la 1,3 bisfosfoglicerat G. Sinteza 3-fosfoglicerat (Transferul grupării fosfat pe ADP) cu sinteza de ATP H. Translocarea fosfatului de pe C 3 pe C 2 (Izomerizarea 3- fosfoglicerat la 2- fosfoglicerat) I. Deshidratarea 2- fosfoglicerat cu formarea fosfoenolpiruvat (PEP) J. Sinteza Py (Transferul fosfatului de pe PEP pe ADP)
 la 1,3 bisfosfoglicerat. G. Sinteza 3-fosfoglicerat (Transferul grupării fosfat pe ADP) cu sinteza de ATP. H. Translocarea fosfatului de pe C 3 pe C 2 (Izomerizarea 3- fosfoglicerat la 2- fosfoglicerat) I. Deshidratarea 2- fosfoglicerat cu formarea fosfoenolpiruvat (PEP) J. Sinteza Py (Transferul fosfatului de pe PEP pe ADP)")
71
REACŢIILE GLICOLIZEI F. Oxidarea gliceraldehid-3-fosfat (GAP) la 1,3 bisfosfoglicerat în prezenţa gliceraldehid-3-fosfat-dehidrogenaza (GAPDH), este prima reacţie de oxidoreducere a glicolizei ce va conduce la formarea de NADH, H+: Gruparea carboxil rezultată din oxidarea funcţiei aldehidice este legată printr-o legătură macroergică de fosfat, rezultând un ester carboxilfosfat
 la 1,3 bisfosfoglicerat în prezenţa gliceraldehid-3-fosfat-dehidrogenaza (GAPDH), este prima reacţie de oxidoreducere a glicolizei ce va conduce la formarea de NADH, H+: Gruparea carboxil rezultată din oxidarea funcţiei aldehidice este legată printr-o legătură macroergică de fosfat, rezultând un ester carboxilfosfat.")
72
REACŢIILE GLICOLIZEI F. Oxidarea gliceraldehid-3-fosfat (GAP) la 1,3 bisfosfoglicerat Cele 2 molecule de GAP obţinute în prima fază vor fi metabolizate la Py, secvenţa enzimatică fiind descrisă pornind de la o moleculă GAP Enzima care catalizează reacţia este gliceraldehid-3-fosfat dehidrogenaza, GAPDH, în prezenţa fosfatului anorganic. Produsul obţinut este 1,3-bisfosfoglicerat. Electronii eliberaţi sunt preluaţi de NAD+. Reacţia este reversibilă

73
REACŢIILE GLICOLIZEI Pt ca glicoliza să poată continua, NADH rezultat trebuie reoxidat la NAD + prin 2 mecanisme: ○ conversia Py la Lactat (reacţie anaerobă) ○ oxidarea NADH în lanţul respirator mitocondrial Energia fosfatului macroergic facilitează sinteza ATP-ului în următoarea reacţie a glicolizei
 ○ oxidarea NADH în lanţul respirator mitocondrial. Energia fosfatului macroergic facilitează sinteza ATP-ului în următoarea reacţie a glicolizei.")
74
REACŢIILE GLICOLIZEI G. Sinteza 3-fosfoglicerat (Transferul grupării fosfat pe ADP) cu sinteza de ATP: Reacţia este reversibilă în condiţii fiziologice, obţinându-se 3- fosfoglicerat şi ATP= fosforilare la nivel de substrat (energia necesară formării unui compus macroergic provine din clivajul unui S, nu din lanţul transportor de electroni) 1moleculă G generează 2 molecule 1,3BPG reacţia înlocuieşte cele 2 molecule de ATP consumate
 1moleculă G generează. 2 molecule 1,3BPG. reacţia înlocuieşte cele. 2 molecule de ATP consumate.")
75
REACŢIILE GLICOLIZEI Sinteza 2,3-bifosfogliceratului în eritocite
2,3 DPG prezent în celule în cantităţi infime, în eritrocite există în concentraţii crescute, glicoliza fiind modificată prin aceste reacţii de şuntare Fosfoglicerat- kinaza 1,3 BPG Mutaza 3-Fosfoglicerat Fosfataza 2,3 DPG

77
REACŢIILE GLICOLIZEI Mecanismul intoxicaţiei cu arsenic
Toxicitatea arsenicului inhibă: piruvat- dehidrogenaza (cofactor- acidul lipoic) Arsenicul pentavalent împiedică formarea de ATP si NADH în calea glicolizei, fără a inhiba direct această cale: Arsenicul intră în competiţie directă cu fosfatul anorganic ca substrat al GAPDH (gliceraldehid-3-fosfat-dehidrogenazei), formând un complex care hidrolizează spontan cu formare de 3-fosfoglicerat Prin scurtcircuitarea sintezei şi a defosforilării moleculei de 1,3- DPG, celula este privată de energia produsă în cursul glicolizei
 Arsenicul pentavalent împiedică formarea de ATP si NADH în calea glicolizei, fără a inhiba direct această cale: Arsenicul intră în competiţie directă cu fosfatul anorganic ca substrat al GAPDH (gliceraldehid-3-fosfat-dehidrogenazei), formând un complex care hidrolizează spontan cu formare de 3-fosfoglicerat. Prin scurtcircuitarea sintezei şi a defosforilării moleculei de 1,3- DPG, celula este privată de energia produsă în cursul glicolizei.")
78
REACŢIILE GLICOLIZEI H. Translocarea fosfatului de pe C 3 pe C 2 (Izomerizarea 3- fosfoglicerat la 2- fosfoglicerat) este catalizată de fosfoglicerat- mutază, fiind o reacţie reversibilă spontan I
 este catalizată de fosfoglicerat- mutază, fiind o reacţie reversibilă spontan. I.")
79
REACŢIILE GLICOLIZEI I. Deshidratarea 2- fosfoglicerat cu formarea fosfoenolpiruvat (PEP) are loc sub acţiunea enolazei inducând redistribuirea energiei în cadrul moleculei cu formarea fosfoenolpiruvatului (PEP) ce conţine o grupare enol-fosfat macroergică. Reacţia este reversibilă în pofida caracterului macroergic al produsului
 ce conţine o grupare enol-fosfat macroergică. Reacţia este reversibilă în pofida caracterului macroergic al produsului.")
80
REACŢIILE GLICOLIZEI J. Sinteza Py (Transferul fosfatului de pe PEP pe ADP) Transferul fosfatului de pe PEP pe ADP este catalizat de piruvat-kinază (fosfotransferaza)= a II-a fosforilare de substrat ;a 3-a reacţie ireversibilă a glicolizei
= a II-a fosforilare de substrat ;a 3-a reacţie ireversibilă a glicolizei.")
81
REACŢIILE GLICOLIZEI 1. Reglarea prin conexiune directă (feed- forward) a piruvat-kinazei: La nivel hepatic, piruvat-kinaza este activată de Fr-1,6-difosfat, produsul reacţiei catalizate de fosfofructokinază Reglarea feed- forward este diferită de mecanismul feedback, determinând cuplarea celor două kinaze: ○ creşterea activităţii FFrK conduce la creşterea [Fr-1,6- difosfat], care activează piruvat-kinaza

82
REACŢIILE GLICOLIZEI 2. Modularea covalentă a piruvat-kinazei:
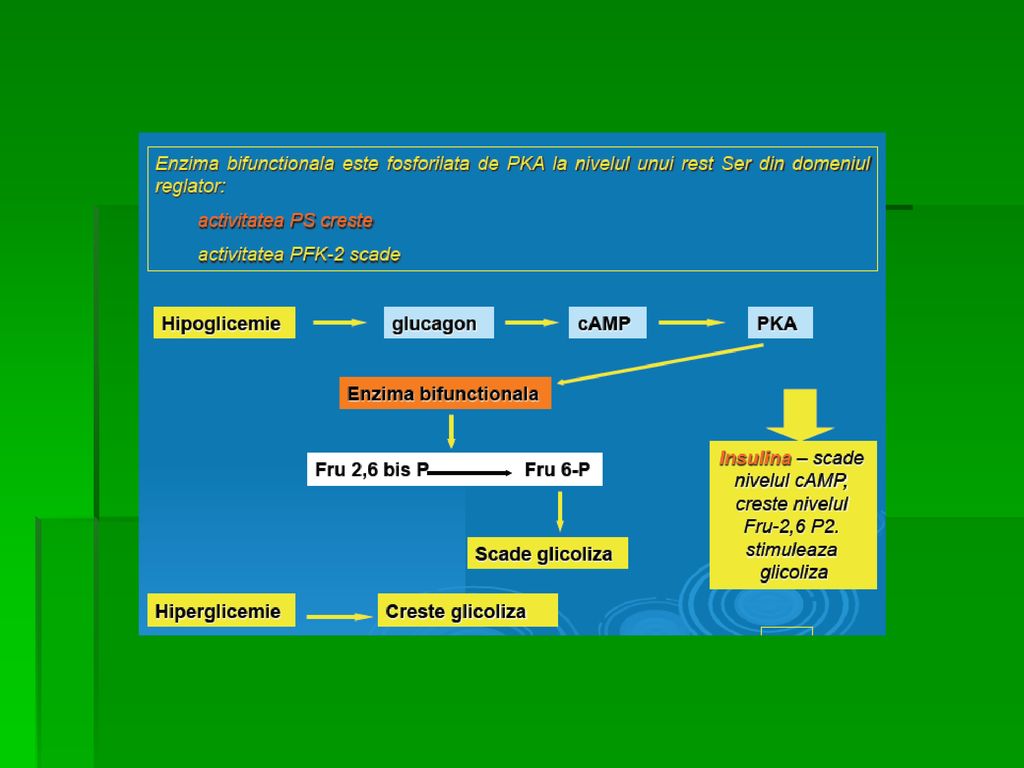
Fosforilarea unei protein-kinaze AMPc- dependente determină inactivarea piruvat-kinazei hepatice, mecanism declanşat la glicemie scăzută (glucagonul stimulează creşterea [cAMP]) PEP nu mai participă la recţiile glicolizei şi pătrunde în circuitul GNG; glucagonul inhibă glicoliza hepatică şi stimulează GNG Defosforilarea piruvat kinazei sub acţiunea unei fosfoprotein- fosfataze determină reactivarea enzimei
 PEP nu mai participă la recţiile glicolizei şi pătrunde în circuitul GNG; glucagonul inhibă glicoliza hepatică şi stimulează GNG. Defosforilarea piruvat kinazei sub acţiunea unei fosfoprotein- fosfataze determină reactivarea enzimei.")
83
REACŢIILE GLICOLIZEI 2. Modularea covalentă a piruvat-kinazei:

84
REGLAREA GLICOLIZEI

85
REACŢIILE GLICOLIZEI 3. Deficitul de piruvat-kinază ca frecvenţă, reprezintă a doua cauză de anemie hemolitică prin deficit enzimatic (după deficitul de glucozo-6-fosfat dehidrogenaza) Eritrocitul matur, normal, nu dispune de mitocondrii, pt producerea de ATP fiind complet dependent de glicoliză ATP: necesităţi metabolice eritrocitare; combustibil biologic al pompelor ionice ce asigură menţinerea formei biconcave flexibile a hematiei
 Eritrocitul matur, normal, nu dispune de mitocondrii, pt producerea de ATP fiind complet dependent de glicoliză. ATP: necesităţi metabolice eritrocitare; combustibil biologic al pompelor ionice ce asigură menţinerea formei biconcave flexibile a hematiei.")
86
REACŢIILE GLICOLIZEI Anemia hemolitică din deficitul de piruvat-kinază: consecinţă a modificării membranei eritrocitare cu alterarea formei hematiei fagocitarea hematiilor de către SRE (macrofagele splenice) 95% dintre defectele genetice ale enzimelor glicolitice 4% deficit de fosfogluco- izomerază formă cronică de gravitate medie sau severă (transfuzii repetate)
 95% dintre defectele genetice ale enzimelor glicolitice. 4% deficit de fosfogluco- izomerază. formă cronică de gravitate medie sau severă (transfuzii repetate)")
87
REACŢIILE GLICOLIZEI Reducerea Py la lactat are loc la nivelul structurilor slab vascularizate/ lipsite de mitocondrii Sinteza musculară de lactat este favorizată de creşterea raportului NADH/ NAD + când sinteza NADH în prezenţa GAPDH şi a celor 3 dehidrogenaze NAD + - dependente din ciclul citric depăşeşte capacitatea oxidativă a lanţului respirator

88
REACŢIILE GLICOLIZEI Ciclul lui Cori Sinteza musculară de lactat
În efort fizic intens, lactatul se acumulează în muşchi şi determină scăderea pH celular, crampe musculare Cea mai mare parte difuzează în sângele circulant fiind utilizat la nivel hepatic pt sinteza glucozei Ciclul lui Cori

89
REACŢIILE GLICOLIZEI Utilizarea lactatului
Sensul reacţiei catalizate de LDH depinde de : ○ [lactat], [piruvat] icel. ○ NADH/ NAD + La nivel hepatic şi cardiac, NADH/ NAD + este < faţă de muşchiul în efort, lactatul fiind oxidat la Py În ficat, Py este convertit în G prin GNG sau oxidat în ciclul citric Muşchiul cardiac este singurul capabil să oxideze lactatul în CO2 şi apă pe calea ciclului citric

90
Ciclul Cori sau cooperarea metabolică muşchi striat-ficat

91
REACŢIILE GLICOLIZEI Acidoza lactică = creşterea [lactat] ce însoţeşte colapsul circ. din ○ infarctul miocardic ○ embolia pulmonară ○ hemoragiile necontrolate ○ şoc Excesul de oxigen necesar recuperării după o perioadă de în care aportul de oxigen a fost inadecvat = "datorie de oxigen" Lactacidemia permite depistarea rapidă a datoriei de oxigen, permite monitorizarea recuperării pacientului cu şoc
![REACŢIILE GLICOLIZEI Acidoza lactică = creşterea [lactat] ce însoţeşte colapsul circ. din. ○ infarctul miocardic.](http://slideplayer.gr/slide/14451298/90/images/91/REAC%C5%A2IILE+GLICOLIZEI+Acidoza+lactic%C4%83+%3D+cre%C5%9Fterea+%5Blactat%5D+ce+%C3%AEnso%C5%A3e%C5%9Fte+colapsul+circ.+din.+%E2%97%8B+infarctul+miocardic..jpg "○ embolia pulmonară. ○ hemoragiile necontrolate. ○ şoc. Excesul de oxigen necesar recuperării după o perioadă de în care aportul de oxigen a fost inadecvat = datorie de oxigen Lactacidemia permite depistarea rapidă a datoriei de oxigen, permite monitorizarea recuperării pacientului cu şoc.")
92
APORTUL ENERGETIC AL GLICOLIZEI
Glicoliza anaerobă 2 molecule ATP/ moleculă de G fără consum/sinteză NADH Glicoliza aerobă 2 molecule ATP/ moleculă de G; 1 moleculă NADH în l. resp. mitoc.= 3 ATP; pt a ajunge în MMI se folosesc navete

94
REGLAREA HORMONALĂ A GLICOLIZEI
Reglarea alosterică a glicolizei sau prin fosforilarea/ defosforilarea E limitante de viteza este de scurtă durată Reglarea hormonală determină creşterea de ori a activităţii enzimatice (ore, zile) Mesele bogate în CH sau insulina activează transcripţia genică precum şi creşterea [glucokinază], [FFrK] şi [piruvat-kinaza] la nivel hepatic cu activarea glicolizei Transcripţia genică şi sinteza celor 3 E sunt reduse à jeun, în DZ, glucagon plasmatic şi insulina
 Mesele bogate în CH sau insulina activează transcripţia genică precum şi creşterea [glucokinază], [FFrK] şi [piruvat-kinaza] la nivel hepatic cu activarea glicolizei. Transcripţia genică şi sinteza celor 3 E sunt reduse à jeun, în DZ, glucagon plasmatic şi insulina.")
95
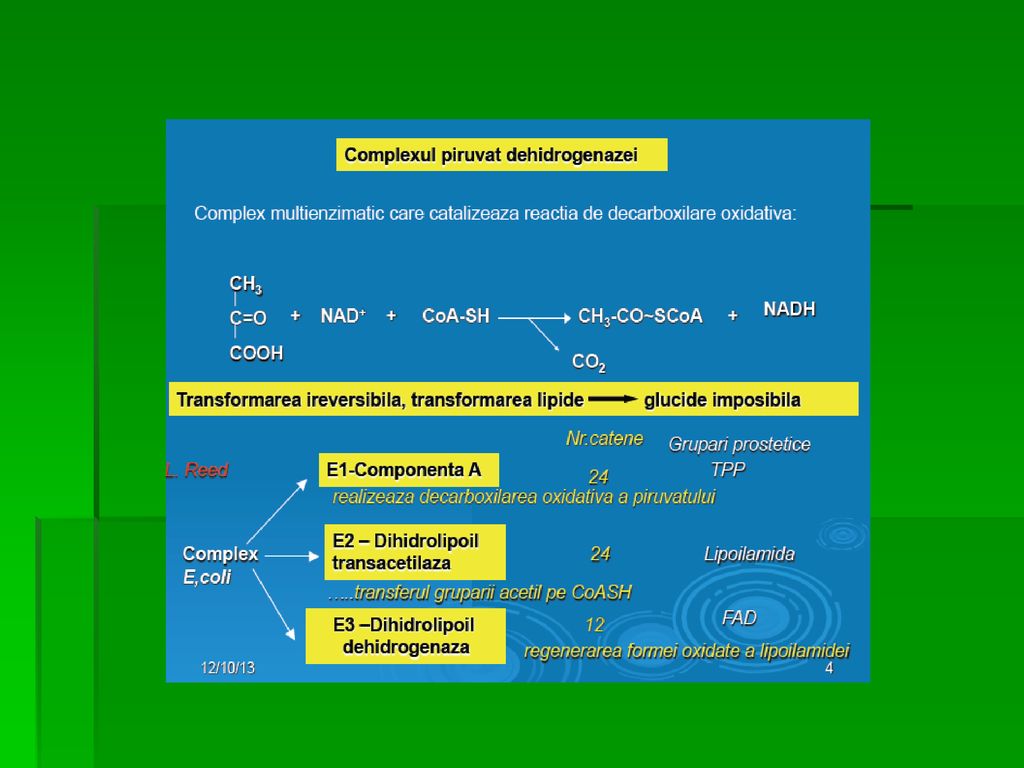
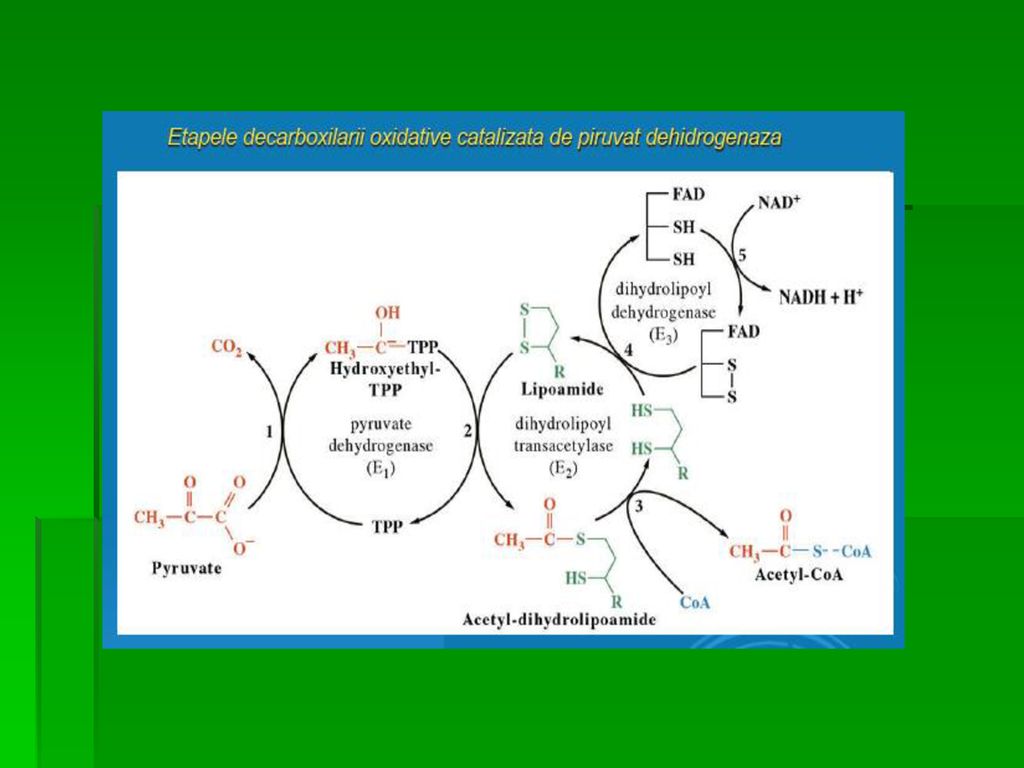
GLICOLIZA Căile de transformare a piruvatului
A. Decarboxilarea oxidativă a Py este importantă în ţesuturile cu capacitate oxidativă crescută (miocard) Complexul piruvat- dehidrogenazei asigură conversia ireversibilă a Py în acetil- CoA, principalul combustibil al ciclului citric şi elementul esenţial al sintezei AG inhibă PDH
 Complexul piruvat- dehidrogenazei asigură conversia ireversibilă a Py în acetil- CoA, principalul combustibil al ciclului citric şi elementul esenţial al sintezei AG. inhibă PDH.")
98
GLICOLIZA B. Carboxilarea Py la OAA sub acţiunea piruvat-carboxilazei biotin-dependentă sursă de intermediari ai ciclului citric, substrat al GNG Piruvat-carboxilaza este activată de acetil- CoA şi catalizează o reacţie ireversibilă

99
GLICOLIZA C. Reducerea Py la etanol sau fermentaţia alcoolică (metabolismul microorganismelor)
")
Παρόμοιες παρουσιάσεις