Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
Variabilitatea genetica
Recombinarea genetica Variatia (diversitatea) genetica
 genetica.")
2
DEFINITIE: variabilitatea este
(1) tendinta caracteristicilor genetice individuale de a fi diferite la nivel populational (2) potentialul unui genotip de a se modifica, atunci cand este supus actiunii unor factori de mediu sau genetici
 tendinta caracteristicilor genetice individuale de a fi diferite la nivel populational. (2) potentialul unui genotip de a se modifica, atunci cand este supus actiunii unor factori de mediu sau genetici.")
3
Definitie Variabilitatea genetica ≠ variatia genetica (sau diversitatea genetica). Daca variabilitatea descrie masura in care un genotip sau fenotip (caracter) tinde sa fie diferit (sa varieze), variatia genetica masoara numarul efectiv de modificari la nivelul populatiilor. Desi deseori folosite ca sinonime cele doua notiuni difera.
 tinde sa fie diferit (sa varieze), variatia genetica masoara numarul efectiv de modificari la nivelul populatiilor. Desi deseori folosite ca sinonime cele doua notiuni difera.")
4
DEFINITIE Variatia genetica a unei populatii este masurata prin gradul de polimorfism, respectiv heterozigozitate Variatia se refera la actualele diferente prezente la indivizii unei populatii sau grup studiat si poate fi direct observata ca o colectie de trasaturi/caracteristice.

5
DEFINITIE Variabilitatea descrie potentialul sau capacitatea de a varia, de a fi diferit. Comparativ cu variatia, variabilitatea genetica este mai dificil de masurat. La nivel molecular, de ex. variabilitatea genetica poate fi estimata prin determinarea ratei mutatiilor.

6
Definitie Termenul VG poate fi utilizat pentru:
1. Observarea, descrierea, analiza diferentelor dintre genotipurile (Gt) individuale (si implicit a fenotipurilor/ caracterelor) dintr-o populatie si 2. Evidentierea ratei (gradului) la care un anumit genotip se poate schimba, ca raspuns la actiunea unor factori de mediu sau genetici. Simplificand, variabilitatea studiaza genotipurile la nivel de indivizi si populatii, iar variatia studiaza genotipurile din interiorul unei specii si comparativ intre specii. Definirea conceptului de variabilitate genetica (VG) presupune intelegerea urmatoarelor doua specificari:
 individuale (si implicit a fenotipurilor/ caracterelor) dintr-o populatie si. 2. Evidentierea ratei (gradului) la care un anumit genotip se poate schimba, ca raspuns la actiunea unor factori de mediu sau genetici. Simplificand, variabilitatea studiaza genotipurile la nivel de indivizi si populatii, iar variatia studiaza genotipurile din interiorul unei specii si comparativ intre specii. Definirea conceptului de variabilitate genetica (VG) presupune intelegerea urmatoarelor doua specificari:")
7
Semnificatia VG Cu cat VG este mai mare, cu atat este mai apta populatia sa se adapteze la modificarile mediului, deci sa reziste la boli, modificari ale climei, specii competitoare, etc.

8
Metodele de masurare a VG
3. Experimente de SELECTIE ARTIFICIALA (raspunsul Gt la modificarile artificiale ale mediului) 1. POLIALELISMUL sporeste VG 2. Determinarea RATEI MUTATIILOR indica in ce masura (cat de repede /incet) se adapteza un anumit genotip la stimulii genetici sau din mediu. O rata inalta a mutatiilor indica faptul ca gena ar trebui sa fie capabila sa raspunda rapid noilor conditii, ceea ce este specific unei VG mari.
 1. POLIALELISMUL sporeste VG. 2. Determinarea RATEI MUTATIILOR indica in ce masura (cat de repede /incet) se adapteza un anumit genotip la stimulii genetici sau din mediu. O rata inalta a mutatiilor indica faptul ca gena ar trebui sa fie capabila sa raspunda rapid noilor conditii, ceea ce este specific unei VG mari.")
9
Surse de variabilitate
Pentru ca selectia naturala sa poata actiona in cadrul unei populatii, este nevoie ca indivizii ce o alcatuiesc sa fie diferiti. Variabilitatea la eucariote are urmatoarele 5 mecanisme posibile: 1. Aparitia mutatiilor 2. Diploidia 3. Reproducerea sexuata 4. Incrucisarile in afara speciei 5. Polimorfismele fenotipice echilibrate in populatia studiata

10
Natura dovedeste ca legendele isi au radacinile in realitate.

11
Mutatiile asigura aparitia unor noi variante alelice (polialelism sau polimorfism genic).
Diploidia, adica prezenta celor doi cromozomi omologi in celule, contribuie la pastrarea in genofond (= totalitatea genelor unei populatii) a alelelor recesive. In stare de heterozigotie aceste alele nu sunt supuse selectiei naturale, raman “ascunse” si “depozitate” pentru generatiile viitoare, pentru descendenti.
 a alelelor recesive. In stare de heterozigotie aceste alele nu sunt supuse selectiei naturale, raman ascunse si depozitate pentru generatiile viitoare, pentru descendenti.")
12
Reproducerea sexuata creeaza indivizi cu noi combinatii alelice
Reproducerea sexuata creeaza indivizi cu noi combinatii alelice. Aceste rearanjamente (sau aceasta recombinare genetica) isi au originea in 3 evenimente, care au loc in timpul reproducerii sexuate: 1. crossing-overul 2. asortarea independenta a omologilor 3. cuplarea intamplatoare a gametilor
 isi au originea in 3 evenimente, care au loc in timpul reproducerii sexuate: 1. crossing-overul. 2. asortarea independenta a omologilor. 3. cuplarea intamplatoare a gametilor.")
13
Recombinarea intracromozomica
1. Crossing - overul sau schimbul de fragmente egale de ADN intre cromatidele nesurori ale cromozomilor omologi din pahitenul si diplotenul profazei meiozei reductionale/ heterotipice are drept urmare modificarea continutului informational al cromozomilor recombinanti. Acesti cromozomi nu vor mai aduce informatia de la un singur ascendent/predecesor.

14
Recombinarea intercromozomica
2. Asortarea independenta a omologilor in metafaza primara creeaza celule fiice avand combinatii intamplatoare de cromozomi materni si paterni Dispunerea cromozomilor in placa metafazica va influenta orientarea lor spre un pol sau celalalt al celulei in anafaza primara. Daca unii cromozomi vor fi de origine materna, altii vor fi de origine paterna in ovocitele de ordin II si spermatocitele de ordin II.

15
In concluzie fiecare individ are o structura genica unica.
Prin asortarea independenta a cromozomilor se pot forma 223 tipuri de gameti, adica peste 8,4 milioane de gameti diferiti, la fiecare dintre cele doua sexe. Dupa fecundare pot rezulta teoretic 246 combinatii, valoare ce depaseste de opt mii de ori populatia actuala a globului. Aceste cifre sunt insa calculate pentru situatia in care cromozomii se deosebesc prin alelele unei singure perechi de gene, ceea ce nu corepunde realitatii! In concluzie fiecare individ are o structura genica unica. Genetica Medicala – Covic + colab, pag.243

16
Recombinarea genomica
3. Combinarea intamplatoare a gametilor in cursul fertilizarii contribuie la diversificarea combinatiilor genice la nivelul zigotului. Desi se considera ca fiind un eveniment in mare masura supus intamplarii, nu toti spermatozoizii au aceeasi viteza de deplasare (legat se pare de forma lor).
.")
17
Importanta recombinarii
1. mareste rata starii de heterozigotie 2. determina dinamica evolutiei biologice, prin amplificarea diversitatii 3. determina o capacitate de adaptare (raspuns) diferentiata a indivizilor la modificarile factorilor de mediu (de ex. medicamente) 4. intervine in realizarea tipului genetic constitutional individual
 diferentiata a indivizilor la modificarile factorilor de mediu (de ex. medicamente) 4. intervine in realizarea tipului genetic constitutional individual.")
18
Surse de variabilitate
la eucariote : 1. Aparitia mutatiilor 2. Diploidia 3. Reproducerea sexuata 4. Incrucisarile in afara speciei 5. Polimorfismele fenotipice echilibrate in populatia studiata

19
Incrucisari in afara speciei?.....
doar Sci-fi!

20
Incrucisarile in afara speciei
Liger = hibrid intre un leu (Panthera leo) si o tigroaica (Panthera tigris), avand parinti apartinand aceluiasi gen dar unor specii diferite. Este diferit de tiglon. Este cea mai mare felina cunoscuta. Inoata precum tigrii si este sociabil precum leii. Exista doar in captivitate deoarece habitatele speciilor parentale nu se suprapun. De observat, tipic ei cresc atat cat sunt ambii parinti pusi la un loc.
 si o tigroaica (Panthera tigris), avand parinti apartinand aceluiasi gen dar unor specii diferite. Este diferit de tiglon. Este cea mai mare felina cunoscuta. Inoata precum tigrii si este sociabil precum leii. Exista doar in captivitate deoarece habitatele speciilor parentale nu se suprapun. De observat, tipic ei cresc atat cat sunt ambii parinti pusi la un loc.")
21
Polimorfismele fenotipice echilibrate in populatia studiata
Deseori un singur fenotip asigura cea mai buna adaptare, in timp ce altele sunt mai putin avantajoase In astfel de cazuri, frecventa alelelor pentru caracterul avantajos va creste in populatie in detrimentul frecventei celorlalte alele

22
Polimorfismele fenotipice
In general insa, in multe populatii se observa coexistenta a doua sau mai multe fenotipuri diferite Prezenta polimorfismelor in cadrul populatiilor se explica prin 3 mecanisme: - heterosis (hibrizi vigurosi) - avantajul minoritatii - avantajul heterozigotilor
 - avantajul minoritatii. - avantajul heterozigotilor.")
23
Polimorfismele fenotipice
Heterosisul consta din selectarea prin hibridare a unor loci avantajosi desi heterozigoti ; de ex. porumbul va fi mai rezistent la imbolnaviri, urmare a reducerii locilor cu alele recesive homozigote, ce il predispun la boala. Reproducerea sexuata prin recombinarea genomica asigura atat ereditatea (continuitatea, pastrarea genelor si respectiv caracterelor), cat si variabilitatea. Descendentii indivizilor neinruditi vor prezenta o vitalitate sporita, calitati noi, o adaptabilitate si o feritilitate crescute, datorita heterozigotismului. In cazul consanguinitatii, gradul de VG se reduce si apare o omogenizare genetica si respectiv fenotipica (se reduce sau dispare heterozigotismul si creste homozigotismul)
, cat si variabilitatea. Descendentii indivizilor neinruditi vor prezenta o vitalitate sporita, calitati noi, o adaptabilitate si o feritilitate crescute, datorita heterozigotismului. In cazul consanguinitatii, gradul de VG se reduce si apare o omogenizare genetica si respectiv fenotipica (se reduce sau dispare heterozigotismul si creste homozigotismul)")
24
Polimorfismele fenotipice
Avantajul minoritatii apare cand cel mai putin obisnuit fenotip reprezinta un avantaj selectiv; de ex., atunci cand o prada prezentand caracteristici rareori intalnite este evitata de pradator, acesta avand o “imagine de cautare” sau reprezentare standard a vanatului. La om, de ex. in infectia cu HIV (virusul imunodeficientei umane) sunt prezente o serie de specii virale mutante, care se dezvolta independent de restul populatiei virale si care eludeaza tratamentul antiviral; initial minoritare acestea pot deveni insa dominante servind drept un rezervor de variabilitate si accelerand instalarea rezistentei la terapie.
 sunt prezente o serie de specii virale mutante, care se dezvolta independent de restul populatiei virale si care eludeaza tratamentul antiviral; initial minoritare acestea pot deveni insa dominante servind drept un rezervor de variabilitate si accelerand instalarea rezistentei la terapie.")
25
Polimorfismele fenotipice
Avantajul heterozigotilor apare atunci cand starea de heterozigot acorda un avantaj selectiv superior starii de homozigot, a.i toate cele 3 genotipuri si implicit fenotipurile pe care le determina, se vor pastra in cadrul populatiei. In cursul evolutiei de obicei la un moment dat au predominat heterozigotii, adaptati mai bine conditiilor de mediu. Un exemplu la om este anemia falciforma.

26
In alcatuirea unei molecule de hemoglobina intra 4 lanturi de globina

27
Genele care codifica globina, in functie de perioada ontogenetica.
Lanturile de globina sunt codificate de un sistem poligenic: unele gene sunt pe cromozomul 11, altele pe cromozomul 16 Hemoglobina A este alcatuita din lanturi alpha si doua lanturi beta de globina

28
Bose frontale; febra Paloare (anemie) Hepato- si splenomegalie (hemoliza) Substitutia punctiforma* (A T) din codonul 6 al β–globinei determina inlocuirea acidului glutamic cu valina. Dureri osoase Dureri Edeme palmare si plantare; Frotiu sanguin: hematii in secera * transversie: purina inlocuita cu pirimidina Nu toti copii au tabloul clinic complet
 din codonul 6 al β–globinei determina inlocuirea acidului glutamic cu valina. Dureri osoase. Dureri. Edeme palmare si plantare; Frotiu sanguin: hematii in secera. * transversie: purina inlocuita cu pirimidina. Nu toti copii au tabloul clinic complet.")
29
Electroforeza hemoglobinei unui heterozigot, a unei persoane cu anemie falciforma si a uneia normale. Petele de culoare arata pozitiile in care au migrat hemoglobinele in gelul de agaroza fata de punctul de origine Care este oare tipul de transmitere a acestei boli?

30
MALARIA este cauzata de un parazit (Plasmodium falciparum) unicelular, care se transmite cu ajutorul unui vector (femela tantarului Anopheles) de la o persoana infectata la una sanatoasa.
 unicelular, care se transmite cu ajutorul unui vector (femela tantarului Anopheles) de la o persoana infectata la una sanatoasa.")
31
Hematii normale si una in forma de secera. (dg.= drepanocitoza)
Parazitului Plasmodium falciparum nu ‘ii plac’ nici eritrocitele homozigotului/ persoanei bolnave (SS) si nici cele ale heterozigotului/ persoanei purtatoare (AS). In conditii speciale, de presiune scazuta a O2 celulele normale se deformeaza dovedind starea de heterozigot!
 si nici cele ale heterozigotului/ persoanei purtatoare (AS). In conditii speciale, de presiune scazuta a O2 celulele normale se deformeaza dovedind starea de heterozigot!")
32
deces selectati

33
In imaginea anterioara s-a putut observa cum in urma cu sute de ani mutatia producatoare de HbS s-a pastrat in cadrul populatiei umane. In acel moment, starea de heterozigot era avantajoasa, deoarece nu exista un tratament pentru malarie. Astazi, populatiile cu risc crescut sunt consiliate, sa efectueze analize prenuptiale in vederea cunoasterii posibilei stari de heterozigozitate. Daca sotii sunt heterozigoti se recomanda diagnosticul prenatal. Daca produsul de conceptie se dovedeste a fi homozigot pentru mutatia cauzatoare de anemie falciforma se discuta optiunile reproductive (terminarea sau continuarea sarcinii) cu cei doi soti.
 cu cei doi soti.")
34
Cele 3 boli eritrocitare s-au raspandit in populatia umana datorita malariei.

35
Variabilitatea Transferul ADNului poate sa se faca si prin alte mecanisme, in afara de reproducere sexuata, recombinare si anume prin: - conjugarea bacteriana - transductia dintre bacteriofagi si bacterii - transformarea bacteriilor prin plasmide - transfectia in culturi de celule eucariote

36
Recombinarea la bacterii
In 1946, Lederberg si Tatum au aratat ca informatia genetica poate fi schimbata/ transmisa de la o bacterie la alta sau de la o tulpina de bacterii la alta. Aceasta ar corespunde unei asa numite “sexualitati” si ar conduce la recombinarea materialului genetic la procariote

37
Recombinarea la procariote
Bacteriile sunt avantajos de studiat caci: - sunt organisme haploide - au un timp de viata redus - se multiplica rapid (timp redus per generatie) - organismele mutante pot fi usor identificate Prin urmare, un numar aproape nelimitat poate fi testat in scurt timp, ceea ce le face utile in studiul mutatiilor.
 - organismele mutante pot fi usor identificate. Prin urmare, un numar aproape nelimitat poate fi testat in scurt timp, ceea ce le face utile in studiul mutatiilor.")
38
1. 2. In 1952 Joshua si Esther Lederberg au folosit o ‘stampila’ (replica) acoperita cu catifea pentru a muta coloniile din culturile de origine pe: mediu normal si respectiv mediu cu antibiotic. In al doilea caz au crescut doar coloniile rezistente la antibiotic.

39
Experimentul identifica bacteriile mutante
Primul vas Petri contine colonia martor/normala In al doilea este cultura cu mediu mai putin nutritiv (= minimal), iar in urmatoarele doua este mediu minimal imbogatit diferentiat. Bacteriile care lipsesc in al doilea vas sunt fie deficitare in Thr, fie in Arg, fapt dovedit de prezenta unora in al treilea si a altora in al patrulea vas Petri, unde s-a adaugat cate unul din acesti aminoacizi.
, iar in urmatoarele doua este mediu minimal imbogatit diferentiat. Bacteriile care lipsesc in al doilea vas sunt fie deficitare in Thr, fie in Arg, fapt dovedit de prezenta unora in al treilea si a altora in al patrulea vas Petri, unde s-a adaugat cate unul din acesti aminoacizi.")
40
Recombinarea la bacterii
In experimentul lor devenit clasic, Lederberg si Tatum au folosit 2 tulpini bacteriene diferite, ambele auxotrofe (mutante, nu pot creste fara anumite substante din mediul de cultura) Ce au observat? Cand se amesteca cele 2 tulpini, fara a adauga aminoacizi in mediul de cultura, vor creste colonii. Neasteptatul fenomen apare destul de rar (aprox.1 din 107 celule cultivate in Petri), dar a fost evident datorita numarului mare de bacterii puse pe placa.
 Ce au observat Cand se amesteca cele 2 tulpini, fara a adauga aminoacizi in mediul de cultura, vor creste colonii. Neasteptatul fenomen apare destul de rar (aprox.1 din 107 celule cultivate in Petri), dar a fost evident datorita numarului mare de bacterii puse pe placa.")
41
Tulpina A necesita metionina si biotina, iar cea B treonina si leucina.
Apar insa colonii pe un mediu minimal, fara a.a., cand se cultiva impreuna cele 2 tulpini. Interpretare: a avut loc o recombinare intre tulpinile A si B. Proprietatile genetice ale celulelor din tulpinile parentale s-au completat reciproc (“complementare genetica”)
")
42
Recombinarea la bacterii
Schimbul de material genetic intre bacterii (CONJUGAREA) a fost evidentiat cu microscopul optic. Transferul de ADN bacterian decurge numai intr-un singur sens: materialul cromozomial “masculin” este introdus intr-o celula “feminina” Bacteriile Escherichia coli difera in functie de prezenta/absenta factorului de fertilitate F Cand sunt mixate celule F+ si F-, se formeaza niste perechi ‘conjugale’: un pil(us) sexual al bacteriei masculine (F+) se ataseaza la suprafata celulei F-.
 a fost evidentiat cu microscopul optic. Transferul de ADN bacterian decurge numai intr-un singur sens: materialul cromozomial masculin este introdus intr-o celula feminina Bacteriile Escherichia coli difera in functie de prezenta/absenta factorului de fertilitate F. Cand sunt mixate celule F+ si F-, se formeaza niste perechi ‘conjugale’: un pil(us) sexual al bacteriei masculine (F+) se ataseaza la suprafata celulei F-.")
43
Recombinarea la bacterii
Factorul F (de fertilitate) poate fi integrat (1) in cz. bacterian printr-un crossing-over specific. Cz. nou format se numeste Hfr (“high frequency of recombination”), deoarece are o frecventa mare de recombinare cu genele altor bacterii in urma procesului de conjugare.
 poate fi integrat (1) in cz. bacterian printr-un crossing-over specific. Cz. nou format se numeste Hfr ( high frequency of recombination ), deoarece are o frecventa mare de recombinare cu genele altor bacterii in urma procesului de conjugare.")
44
Recombinarea bacteriana Bacteriile pot contine factorul F de fertilitate sub forma unui cz. mic aditional (2), respectiv o molecula circulara de ADN de aprox pb, plasmidul F Acesta reprezinta cam 1/40 din totalul informatiei genetice continute in cz. bacterian. Intr-o celula apare un singur plasmid si el poate fi transferat altei bacterii.
, respectiv o molecula circulara de ADN de aprox pb, plasmidul F. Acesta reprezinta cam 1/40 din totalul informatiei genetice continute in cz. bacterian. Intr-o celula apare un singur plasmid si el poate fi transferat altei bacterii.")
45
Aprox. 1/3 din ADNul factorului F (plasmidului) consta din gene de transfer, inclusiv gene pentru formarea pililor sexuali. Transferul factorului F incepe dupa ce se deschide dubla catena. O monocatena este transferata celulei acceptoare, se replica si devine dubla. Si monocatena ramasa in celula donoare se replica redevenind dubla. In final ambele celule sunt de tip F+

46
Recombinarea bacteriana
Uneori cromozomul se rupe in timpul transferului. Astfel, bacteriile F- receptoare mostenesc o copie incompleta a elementului F. Celulele F- , care au primit doar o parte din cz. donor se numesc zigoti partiali sau hemizigoti si raman de tip F- Daca transferul este complet, bacteria receptoare va mosteni un element F complet si proprietatea Hfr; descendentii sai vor fi tot celule Hfr, respectiv F+ Cand cromozomii bacterieni contin diferiti markeri genetici, celulele au suferit frecvente recombinari.

47
Bacteriofag = Virus bacterian
Desi initial (1941) s-a crezut ca s-a identifi-cat o modalitate de combatere a infectii-lor bacteriene, de fapt acestia au fost utilizati pt a analiza bacteriile. Reproducerea bacteriofagilor nu are loc totdeauna imediat dupa infectarea celulei. Ocazional, ADNul fagic este integrat in cz. bacterian si replicat odata cu acesta (ciclul lizogenic)
 s-a crezut ca s-a identifi-cat o modalitate de combatere a infectii-lor bacteriene, de fapt acestia au fost utilizati pt a analiza bacteriile. Reproducerea bacteriofagilor nu are loc totdeauna imediat dupa infectarea celulei. Ocazional, ADNul fagic este integrat in cz. bacterian si replicat odata cu acesta (ciclul lizogenic)")
48
In acest caz bacteriofagul Lambda s-a inserat prin crossing-over in ADNul bacteriei E.coli, dupa formarea unui inel. Atasarea se face la o secventa cu omologie din cz.bacterian. Fagul induce ciclul litic si apoi se elibereaza prin mecanismul invers.

49
In 1952 Zinder si Lederberg au descris un nou tip de recombina-re intre 2 tulpini de bacterii.
Fagi care au infectat anterior bacterii, care produceau lactoza, au transferat aceasta proprietate noilor gazde. Transductia = transferul unui mic fragment de ADN dintr-un cz. bacterian prin intermediul unui bacteriofag in alta celula bacteriana.

50
Transductia poate fi de 2 feluri:
T. Generalizata: fagul se insera oriunde in genomul gazdei T. Specializata: fagul se insera numai la nivelul unui segment de ADN cz. particular

51
Transformarea prin plasmide
Celulele modificate genetic prin incorporarea de ADN sunt considerate “transformate”. Termenul se refera de obicei la rezultat si nu la mecanism. Transformarea prin plasmide Plasmidele = molecule mici, circulare de ADN, care se replica autonom si care nu intra in componenta cz. din celula bacteriana in care se gasesc Deseori contin gene de rezistenta la antibiotice (de ex. la ampicilina) Incorporarea plasmidului transforma celula bacteriana initial sensibila la ampicilina in una rezistenta.
 Incorporarea plasmidului transforma celula bacteriana initial sensibila la ampicilina in una rezistenta.")
52
Plasmidele sunt utilizate ca vectori in transferul de ADN.
Pentru aceasta se utilizeaza un mediu selectiv a.i vor creste pe el doar bacteriile care au incorporat plasmidul recombinant continand ADNul de investigat.

53
Transfectia = transferul de ADN intre celule eucariote in cultura
- Poate fi folosita in analiza transmiterii unor caracteristici genetice In cele 2 cazuri alaturate fibroblastele normale pierd “contactul de inhibitie” si se transforma asemanator celulelor cu care vin in contact. Astfel de studii au fost initial utilizate pentru a identifica oncogene (gene cauzatoare de cancere)
")
54
Un alt nivel al variabilitatii genetice
Contrar celor crezute anterior, copia ARN nu este totdeauna fidela ADN-ului. Atat la copil, cat si la adult s-au evidentiat niste modificari, ce intial au parut erori de transcriptie. Aceste modificari au fost detectate in numar semnificativ in ARN-ul leucocitelor, celulelor din piele si din creier. Modificarile fiind mereu aceleasi se presupune, ca au un rol. De exemplu, in limfocitele B (la un grup de 27 persoane) peste de nucleotide au fost gasite modificate, ceea ce a insemnat una (sau mai multe) ‘erori’ in fiecare gena transcrisa, respectiv in 40% din genele translatate (traduse).
 peste de nucleotide au fost gasite modificate, ceea ce a insemnat una (sau mai multe) ‘erori’ in fiecare gena transcrisa, respectiv in 40% din genele translatate (traduse).")
55
Polimorfismele - genetice sunt responsabile pentru cea mai mare parte din diversitatea si individualitatea observate in cadrul speciei umane - proteice au fost studiate primele (ABH, Rh, etc). Cel mai polimorfic sistem genetic uman este considerat sistemul HLA
. Cel mai polimorfic sistem genetic uman este considerat sistemul HLA.")
56
Polimorfisme proteice - exemple
1. Antigene eritrocitare: ABH, Rh, MNS, Xg, Lutheran, Lewis, etc 2. Proteine serice: Hp, Trf, imunoglobuline, alfa-1-antitripsina (A1AT), etc 3. Enzime eritrocitare: PAE, G6PD, etc 4. Enzime plasmatice: pseudocolinesteraza, etc 5. Enzime tisulare: alcool-dehidrogenaza, etc Variatia individuala in populatia generala a polimorfismelor proteice poate atinge 20% din totalul proteinelor.
, etc. 3. Enzime eritrocitare: PAE, G6PD, etc. 4. Enzime plasmatice: pseudocolinesteraza, etc. 5. Enzime tisulare: alcool-dehidrogenaza, etc. Variatia individuala in populatia generala a polimorfismelor proteice poate atinge 20% din totalul proteinelor.")
57
Polimorfisme proteice
Cu cat 2 persoane sunt mai indepartate dpdv genetic, adica apartin unor grupuri etnice diferite, cu atat gradul de diversitate este mai accentuat. Datorita numarului mare de sisteme polimorfice (peste 30) si de variante in fiecare sistem, se poate realiza un numar imens de combinatii Fiecare individ poseda o combinatie specifica a acestor variante, ceea ce il face unic dpdv biologic.
 si de variante in fiecare sistem, se poate realiza un numar imens de combinatii. Fiecare individ poseda o combinatie specifica a acestor variante, ceea ce il face unic dpdv biologic.")
58
Polimorfismul A1AT A1AT produsa in ficat protejeaza plamanul de actiunea elastazei eliberata de neutrofile (in inflamatii). A1AT este codificata de gena SERPINA 1 de la nivel 14q32.1. Gena are peste 100 alele mutante (ex. S si Z). Alela normala este M Gt: MS, MZ, SS, SZ, si ZZ (forma cea mai grava); alelele sunt codominante A1AT = inhibitor de proteaza (PI) Pi ZZ este fenotipul cel mai grav, al deficitului de A1AT, cand sunt manifeste afectarea pulmonara si hepatica. Activitate enzimatica redusa => elastaza poate actiona nestingherita (afectare pulmonara) Agregarea moleculelor de A1AT la nivelul RE al hepatocitelor determina afectarea ficatului (hepatita, ciroza) Alte fenotipuri nu prezinta afectare hepatica. Se estimeaza, ca 95% din cazuri nu sunt dg.
. Alela normala este M. Gt: MS, MZ, SS, SZ, si ZZ (forma cea mai grava); alelele sunt codominante. A1AT = inhibitor de proteaza (PI) Pi ZZ este fenotipul cel mai grav, al deficitului de A1AT, cand sunt manifeste afectarea pulmonara si hepatica. Activitate enzimatica redusa => elastaza poate actiona nestingherita (afectare pulmonara) Agregarea moleculelor de A1AT la nivelul RE al hepatocitelor determina afectarea ficatului (hepatita, ciroza) Alte fenotipuri nu prezinta afectare hepatica. Se estimeaza, ca 95% din cazuri nu sunt dg. gene=SERPINA1.")
59
Secventa ADN-ului nuclear este identica in proportie de 99,9% la doua persoane alese la intamplare.
Genomul uman fiind compus din 3 miliarde de perechi de baze, inseamna ca 0,1% reprezinta aproape 3 milioane de baze azotate, ceea ce determina diferentele fenotipice (uneori dramatice) dintre doua persoane!
 dintre doua persoane!")
60
DIVERSITATEA GENETICA INDIVIDUALA SI POPULATIONALA
Din cele 0,1% secvente de ADN unele nu au efecte sau au efecte fenotipice minore, pe cand altele sunt direct responsabile de producerea unor boli (mutatii punctiforme). Intre aceste doua extreme se gasesc variatiile genetice care au drept urmare variabilitatea fenotipica, precum: - caracterele anatomice - particularitatile fiziologice - intolerantele alimentare - raspunsul imun si/ sau susceptibilitatea la infectii - reactiile adverse si/sau raspunsul la o anumita terapie - predispozitia pentru dezvoltarea anumitor boli (tumori) - caracteristicile temperamentale si psihologice - talentul artistic - aptitudinile sportive, etc
. Intre aceste doua extreme se gasesc variatiile genetice care au drept urmare variabilitatea fenotipica, precum: - caracterele anatomice. - particularitatile fiziologice. - intolerantele alimentare. - raspunsul imun si/ sau susceptibilitatea la infectii. - reactiile adverse si/sau raspunsul la o anumita terapie. - predispozitia pentru dezvoltarea anumitor boli (tumori) - caracteristicile temperamentale si psihologice. - talentul artistic. - aptitudinile sportive, etc.")
61
Un concept important al geneticii umane si medicale
Boala genetica este doar cea mai evidenta si deseori cea mai extrema manifestare a diferentelor genetice, extremitatea unui continuum format din variatii ce se intind de la variante rare ce produc boala, trecand prin variantele mai obisnuite care pot spori susceptibilitatea pentru a dezvolta maladia, pana la cea mai comuna (obisnuita) varianta din cadrul populatiei, care nu are nici o relevanta cunoscuta in patologia umana. Thompson, pag. 175
 varianta din cadrul populatiei, care nu are nici o relevanta cunoscuta in patologia umana. Thompson, pag")
62
Majoritatea PG sunt silentioase fenotipic
O varianta genetica (alela) obisnuita, adica cu o frecventa peste (sau egala cu) 1% in populatia generala, este considerata un POLIMORFISM GENETIC (PG). Majoritatea PG sunt silentioase fenotipic Alelele cu o frecventa < 1% sunt numite variante rare. Nu exista o corelatie directa intre frecventa cu care apare o alela mutanta si efectul sau in patologia umana.
 obisnuita, adica cu o frecventa peste (sau egala cu) 1% in populatia generala, este considerata un POLIMORFISM GENETIC (PG). Majoritatea PG sunt silentioase fenotipic. Alelele cu o frecventa < 1% sunt numite variante rare. Nu exista o corelatie directa intre frecventa cu care apare o alela mutanta si efectul sau in patologia umana.")
63
Motoo Kimura postula teoria neutralista a evolutiei la sfarsitul anilor 1960: nivelul de evolutie moleculara explica evolutia speciei prin mutatiile succesive pastrate in genom si produse cu precadere printr-o deriva aleatorie; diversitatea specifica fiind in mare parte rezultatul acumularii substitutiilor ‘neutre’, care nu modifica adaptabilitatea organismului la mediul sau de viata. Diversitatea genetica este unul din mijloacele prin care populatiile se adapteaza la mediul mereu schimbator. Diversitatea este cu atat mai mare cu cat: - numarul de alele ale unei gene este mai mare - frecventele alelice sunt apropiate, adica nu exista una mai frecventa - frecventele indivizilor heterozigoti si homozigoti sunt echilibrate

64
Polimorfismele pot fi localizate: intre gene in introni in exoni
Se estimeaza ca aprox. 1 din 2500 perechi de baze azotate sunt heterozigote in regiunile ce codifica proteine din genom. Polimorfismele pot fi localizate: intre gene in introni in exoni in regiunile reglatoare

65
Variom-ul = totalitatea modificarilor genetice gasite in populatiile speciilor care au avut o schimbare evolutiva relativ scurta De exemplu, la om, aprox. 1 din 1200 nucleotide difera. Specia umana a aparut acum doar de ani, rata variatiei fiind comparativ mica. In practica, variomul poate fi suma SNPs ( single nucleotide polymorphisms) de la nivelul populatiei.
 de la nivelul populatiei.")
66
Polimorfismele ADN 1. Polimorfismele unui singur nucleotid (SNPs)RFLP
2. Polimorfismele fragmentelor mai mici de 1000 perechi de baze (minisateliti, microsateliti,VNTR, indel) 3. Polimorfismele submicroscopice 4. Polimorfismele microscopice (heteromorfismele heterocromatinei si ale eucromatinei, situsurile fragile)
 3. Polimorfismele submicroscopice. 4. Polimorfismele microscopice (heteromorfismele heterocromatinei si ale eucromatinei, situsurile fragile)")
67
Polimorfismele unui singur nucleotid: SNP
SNPs reprezinta forma cea mai frecventa a polimorfismelor ADN. Prin secventierea genomului uman s-au identificat aprox. 18 milioane de SNPs (1% din genom), 1 SNP la circa de nucleotide, dar frecventa poate fi mai inalta in anumite regiuni, precum insulele CpG s.a 1 la 100 sau mai redusa in interiorul unor gene s.a 1 la pb.
, 1 SNP la circa de nucleotide, dar frecventa poate fi mai inalta in anumite regiuni, precum insulele CpG s.a 1 la 100 sau mai redusa in interiorul unor gene s.a 1 la pb.")
68
SNPs neutre sunt folosite ca markeri genetici
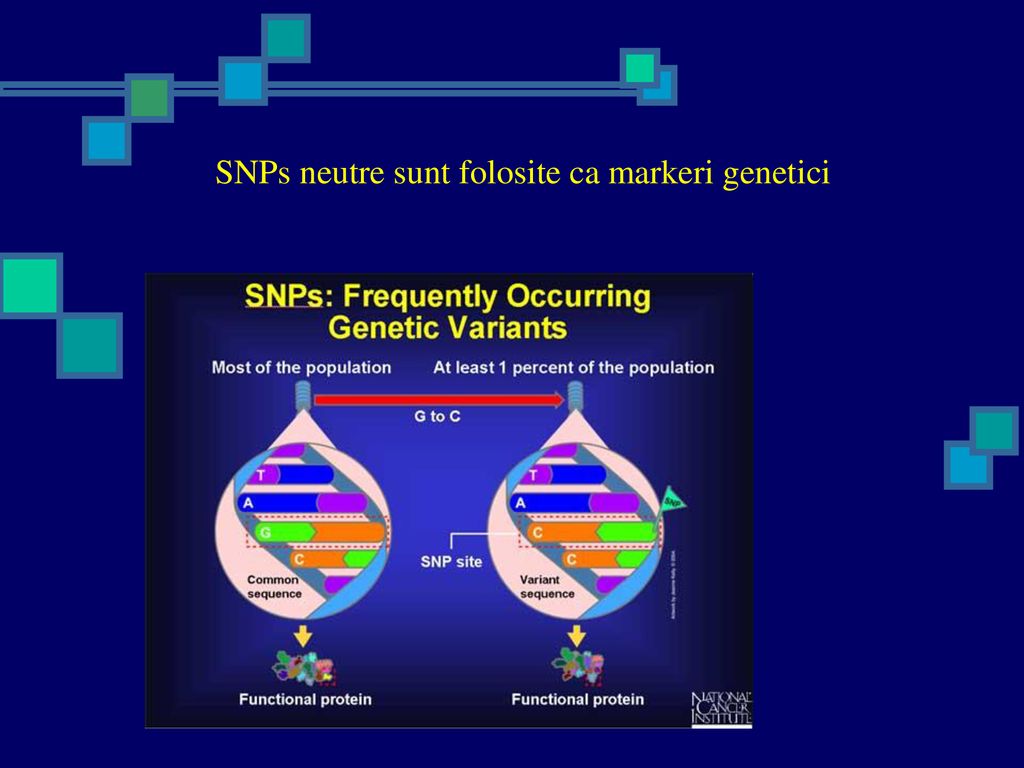
69
SNP inlantuit cu gena Y

70
In prezent SNPs utilizate pentru:
Localizarea genelor de susceptibilitate in cazul bolilor multifactoriale (diabet) folosindu-se studii de dezechilibru al inlantuirii si de asociere a haplotipurilor Studii de farmacogenetica Intelegerea originii si evolutiei speciei umane(migratia populatiei)
 folosindu-se studii de dezechilibru al inlantuirii si de asociere a haplotipurilor. Studii de farmacogenetica. Intelegerea originii si evolutiei speciei umane(migratia populatiei)")
71
SNPs au fost identificate prin digestia ADN-ului cu endonucleaze de restrictie; aceste enzime bacteriene recunosc si sectioneaza ADN-ul la nivelul unei secvente specifice. In acest caz CCTGAGG. Urmare a mutatiei punctiforme (SNP) dispare un situs de restrictie, iar fragmentele de ADN vor fi diferite in cazul lantului normal si respectiv anormal de globina. RFLP (restriction fragment length polymorphism) a fost folosit in dg. genotipic indirect. Fragmentele de restrictie cu lungimi variate sunt evidentiate prin tehnica Southern blot dupa hibridarea cu o proba complementara Vezi

72
“Variatia cea mai mare la nivelul unui locus se evidentiaza in interiorul unei populatii si nu intre populatii”…..adevarat/fals? 85% este variatia genetica in interiorul raselor continentale si numai 15% intre ele. Concret, pentru un locus dat X in populatia 1 frecventa alelei A poate fi 40%, iar in populatia 2 poate fi 45%. Adica mica diferenta nu este informativa. Dar exista loci informativi: o mare parte a variatiei culorii tegumentului la doua populatii poate fi anticipata analizand gena SLC24A5, care codifica o proteina intracelulara membranara (Na, K, Ca) SNP, o substitutie, are un efect fenotipic major. O varianta este aproape fixata in populatia Europei si alta aproape fixata in cea din Africa. Altfel spus variatia genetica in cazul acesta este de aproape 100%, (si nu 15%) INTRE POPULATII interviewL. L. Cavalli-Sforza
 SNP, o substitutie, are un efect fenotipic major. O varianta este aproape fixata in populatia Europei si alta aproape fixata in cea din Africa. Altfel spus variatia genetica in cazul acesta este de aproape 100%, (si nu 15%) INTRE POPULATII interviewL. L. Cavalli-Sforza.")
73
Variatiile identificate in genomul uman
Variatii de secventa- prin modificarea unui singur nucleotid: prin substitutie SNV (single nucleotid variation)< 1% in pop. SNP >1% prin insertie/deletie = DIP (deletion-insertion polymorphism of one base pair) Variante structurale – fragmente de ADN modificate: Variatiile fragmentelor avand intre 2 – nucleotide Variatiile submicroscopice ( 1kb – cateva Mb) Variatiile microscopice cromozomiale sau genomice (heteromorfisme cz., inversii, situsuri fragile, unii cz. markeri) Covic pag 289
< 1% in pop. SNP >1% prin insertie/deletie = DIP (deletion-insertion polymorphism of one base pair) Variante structurale – fragmente de ADN modificate: Variatiile fragmentelor avand intre 2 – nucleotide. Variatiile submicroscopice ( 1kb – cateva Mb) Variatiile microscopice cromozomiale sau genomice (heteromorfisme cz., inversii, situsuri fragile, unii cz. markeri) Covic pag 289.")
74
Variatiile fragmentelor ADN cu lungime intre 2 – 1.000 pb:
1. SSR (short sequence repeats), numite mai intai VNTR (variable number tandem repeats) sunt de 2 feluri: Microsateliti (sau STR): repetitii foarte scurte (1-15pb), frecvent dinucleotidice (CA/TA) sau trinucleotidice, ce formeaza fragmente de 100pb foarte numeroase, polimorfe (cu circa 10 alele/locus)si cu o distributie uniforma: un microsatelit la 1.000pb Minisateliti: repetitii de secvente scurte (15-500pb), ce formeaza fragmente de 1-20 kb, dispersate in structura autozomilor si ocupand situsuri specifice (peste de localizari) Ambele tipuri utile in medicina legala pentru realizarea profi-lului genetic individual, specific (identificare sau paternitate)
, numite mai intai VNTR (variable number tandem repeats) sunt de 2 feluri: Microsateliti (sau STR): repetitii foarte scurte (1-15pb), frecvent dinucleotidice (CA/TA) sau trinucleotidice, ce formeaza fragmente de 100pb foarte numeroase, polimorfe (cu circa 10 alele/locus)si cu o distributie uniforma: un microsatelit la 1.000pb. Minisateliti: repetitii de secvente scurte (15-500pb), ce formeaza fragmente de 1-20 kb, dispersate in structura autozomilor si ocupand situsuri specifice (peste de localizari) Ambele tipuri utile in medicina legala pentru realizarea profi-lului genetic individual, specific (identificare sau paternitate)")
75
Cele 2 persoane se diferentiaza prin ambii loci.
Locusul A este o repetitie in tandem GC: 4 alele posibile, cu cate 2,3,4 sau 5 repetitii (A2, A3, A4, and A5). Locusul B cu motivul AGCT : 2 alele, cu 2 sau 3 repetitii (B2 and B3). AMPRENTA GENETICA include simultan ambii loci. Cele 2 persoane se diferentiaza prin ambii loci. AMPRENTELE GENETICE includ de obicei peste 10 loci polimorfi vezi homework Individ 1: Heterozigot A si homozigot B Individ 2: heterozigot A si B
. Locusul B cu motivul AGCT : 2 alele, cu 2 sau 3 repetitii (B2 and B3). AMPRENTA GENETICA include simultan ambii loci. Cele 2 persoane se diferentiaza prin ambii loci. AMPRENTELE GENETICE includ de obicei peste 10 loci polimorfi. vezi homework. Individ 1: Heterozigot A si homozigot B Individ 2: heterozigot A si B.")
76
S-au identificat 5 VNTR (2 Scurte si 3 Lungi) in 10 combinatii alelice
Genotipuri hTERT MNS16 A MNS16A minisateliti ai genei umane pentru telomeraza : un factor de risc pentru cancerul colorectal Genotype patterns of hTERT MNS16A. Five identified VNTRs were detected in 10 allele combinations named MNS16A genotype patterns 1–10. For SL classification, VNTR-243 and VNTR-274 were considered as short (S) and VNTR-302, VNTR-333 and VNTR-364 as long (L) variant alleles. S-au identificat 5 VNTR (2 Scurte si 3 Lungi) in 10 combinatii alelice Hofer P et al. Carcinogenesis 2011;32: © The Author Published by Oxford University Press. All rights reserved. For Permissions, please
 and VNTR-302, VNTR-333 and VNTR-364 as long (L) variant alleles. S-au identificat 5 VNTR (2 Scurte si 3 Lungi) in 10 combinatii alelice. Hofer P et al. Carcinogenesis 2011;32: © The Author Published by Oxford University Press. All rights reserved. For Permissions, please")
77
heterocromatina

78
Variatiile fragmentelor ADN cu lungime intre 2 – 1.000 pb:
1. SSR (short sequence repeats) = VNTR 2. INDELS = insertion + deletion - secventele nucleotidice sunt scurte - 1/3 din ele sunt intre 200 – 400 pb si reflecta polimorfismul secventelor Alu din genomul uman - utilizate in studiul evolutiei genomului uman si pentru intelegerea structurii sale dinamice
 = VNTR. 2. INDELS = insertion + deletion. - secventele nucleotidice sunt scurte. - 1/3 din ele sunt intre 200 – 400 pb si reflecta polimorfismul secventelor Alu din genomul uman. - utilizate in studiul evolutiei genomului uman si pentru intelegerea structurii sale dinamice.")
79
Polimorfismele submiscroscopice (1kb – cateva Mb)
1. Variatiile numarului de copii (CNV) reprezinta principala sursa de variatie genetica intre indivizi CNP = polimorfismul numarului de copii, cand CNV au o frecventa peste 1% in populatie 33% din genomul uman 2. Polimorfismul inversiilor sau translocatiilor 3. Polimorfismele genelor ARN necodant
 reprezinta principala sursa de variatie genetica intre indivizi. CNP = polimorfismul numarului de copii, cand CNV au o frecventa peste 1% in populatie. 33% din genomul uman. 2. Polimorfismul inversiilor sau translocatiilor. 3. Polimorfismele genelor ARN necodant.")
80
Orice clasificare este relativa, prin ea incercam doar sa intelegem putin mai mult lumea

81
Incercam sa intuim viitorul presupunand ca intelegem trecutul
Παρόμοιες παρουσιάσεις



















