Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
ROLUL STRUCTURA CHIMICĂ
LIPIDELE ROLUL STRUCTURA CHIMICĂ PROPRIETĂŢI

2
Funcţiile biologice ale lipidelor.
Clasificarea lipidelor( structurală, funcţională, după proprietăţile fizico-chimice ). Lipidele de rezervă – acilgliceridele, reprezentanţii, structura, proprietăţile fizico-chimice, rolul biologic. Lipidele protoplasmatice – fosfogliceridele, sfingolipidele, glicolipidele, colesterolul- structura, proprietăţilefizico-chimice, rolul biologic. Eicosanoizii- prostaglandinele, leucotrienele, tromboxanele- structura şi rolul biologic. Vitaminele liposolubile – A,D,E, K – structura şi rolul biologic. Membranele biologice: Funcţiile şi structura – modelul S.G.Sînger şi G.L.Nicolson; Proprietăţile fundamentale – fluiditatea, molilitatea, permeabilitatea selectivă, asimetria, autoasamblarea şi autorapararea; Diversitatea şi specificitatea structurilor şi funcţiilor diferitor membrane biologice.
. Lipidele de rezervă – acilgliceridele, reprezentanţii, structura, proprietăţile fizico-chimice, rolul biologic. Lipidele protoplasmatice – fosfogliceridele, sfingolipidele, glicolipidele, colesterolul- structura, proprietăţilefizico-chimice, rolul biologic. Eicosanoizii- prostaglandinele, leucotrienele, tromboxanele- structura şi rolul biologic. Vitaminele liposolubile – A,D,E, K – structura şi rolul biologic. Membranele biologice: Funcţiile şi structura – modelul S.G.Sînger şi G.L.Nicolson; Proprietăţile fundamentale – fluiditatea, molilitatea, permeabilitatea selectivă, asimetria, autoasamblarea şi autorapararea; Diversitatea şi specificitatea structurilor şi funcţiilor diferitor membrane biologice.")
3
LIPIDE Substanţe organice, greu solubile în apă dar solubile în solvenţi organici (eter, benzen, cloroform).
.")
4
Rolul principala formă de depozitare şi de transport a rezervelor energetice ale organusmului Constituienţi structurali ai membranelor celulare şi intracelulare rol în procesul de comunicare şi recunoaştere intercelulară Izolator: termo; mecano-; electroizolator Pot avea efecte biologice: sunt vitamine, hormoni, prostaglandine

5
Clasificarea Saponifiabile – prin hidroliză se descompun în substanţele componente (acilglicerolii, fosfogliceride, sfingolipide, cerurile, glicolipidele) Nesaponifiabile – nu se scindează hidrolitic în compuşi simpli (hidrocarburile, alcoolii, aldehidele, acizi cu schelete alifatice sau ciclice cu structură poliizoprenică: terpenele, steroizii, carotenoizii)
 Nesaponifiabile – nu se scindează hidrolitic în compuşi simpli (hidrocarburile, alcoolii, aldehidele, acizi cu schelete alifatice sau ciclice cu structură poliizoprenică: terpenele, steroizii, carotenoizii)")
6
Structurală: Lipide monocomponente (monomeri lipidici) – lipide nesaponifiabile Lipidele policomponente Simple: cerurile, gliceridele, steridele Conjugate (mixte): fosfolipidele, sfingolipidele, glicolipidele
: fosfolipidele, sfingolipidele, glicolipidele.")
7
După proprietăţile fizico-chimice:
Polare (FL, AG) Nepolare – Tg
 Nepolare – Tg.")
8
După importanţa fiziologică:
Lipide de rezervă - Tg Structurale – FL, Col, Sfingolipidele Lipidele sângelui: Tg, Col, LP Lipidele constituie 10-20% din masa corpului. 10-12 kg lipide (2-3 kg – lipide structurale; 98% - concentrate în ţesutul adipos)
")
9
AG – structura, proprietăţi
AG- derivaţii hidrocarburilor alifatice ce conţin druparea carboxil Clasificare: 1. După nr atomilor de C: - AG cu nr par - AG cu nr impar 2. După gradul de saturare: saturaţi nesaturaţi 3. După rolul fiziologic: esenţiali neesenţiali (linoleic şi linolenic)
")
10
Structura chimică CH3-COOH acetic CH3-(CH2)2-COOH butiric
CH3-(CH2)4-COOH caproic CH3-(CH2)6-COOH caprilic CH3-(CH2)8-COOH capric CH3-(CH2)10-COOH lauric CH3-(CH2)12-COOH miristic CH3-(CH2)14-COOH palmitic CH3-(CH2)16-COOH stearic
4-COOH caproic. CH3-(CH2)6-COOH caprilic. CH3-(CH2)8-COOH capric. CH3-(CH2)10-COOH lauric. CH3-(CH2)12-COOH miristic. CH3-(CH2)14-COOH palmitic. CH3-(CH2)16-COOH stearic.")
11
AG mononesaturaţi AG nesaturaţi conţin una sau mai multe legături duble etilenice care de regulă sunt între poziţia C9 şi C10 –cis a. palmitooleic (C16:1) CH3- (CH2)5- CH=CH-(CH2)7-COOH a. oleic (C18:1) CH3-(CH2)7-CH=CH-(CH2)7-COOH
 CH3- (CH2)5- CH=CH-(CH2)7-COOH. a. oleic (C18:1) CH3-(CH2)7-CH=CH-(CH2)7-COOH.")
12
AG polinesaturaţi a.linoleic (C18:2 –cis9, 12) CH3-(CH2)4-CH=CH-CH2-CH=CH-(CH2)7- COOH a. linolenic (C18:3 –cis9,12,15) CH3-CH2-CH=CH-CH2-CH=CH-CH2-CH=CH-(CH2)7-COOH a.arahidonic (C20:4, cis5,8,11,14) CH3-(CH2)4-CH=CH-CH2-CH=CH-CH2-CH=CH-CH2-CH=CH-(CH2)3-COOH
7-COOH. a.arahidonic (C20:4, cis5,8,11,14) CH3-(CH2)4-CH=CH-CH2-CH=CH-CH2-CH=CH-CH2-CH=CH-(CH2)3-COOH.")
13
Proprietăţile AG AG saturaţi până la C8 – sunt lichizi; mai sus de C8- solizi. AG nesaturaţi – sunt lichizi În cristale catenele hidrocarburilor saturate au configuraţia de zig-zag AG nesaturaţi prezintă izomerie cis-trans însă formele naturale ale acestor – sunt izomerii cis AG – puţin solubili în apă, solubilitatea scăzând odată cu lungimea catenei AG nu se află în stare liberă în celule şi ţesuturi ci sunt legaţi covalent AG nesaturaţi au t de topire mai joasă comparativ cu cei saturaţi--- lungimea şi prezenţa legăturilor duble amplifică fluiditatea

14
Proprietăţile chimice
AG saturaţi şi nesaturaţi pot forma: Esteri (gliceride, fosfogliceride) Săruri (săpunuri cu proprietăţi tensioactive) Amide (sfingolipide) AG nesaturaţi adiţionează la nivelul dublei legături – halogeni (Br2; Cl2), tiocianatul (SCN)2, gruparea hidroxil AG nesaturaţi suferă procesul de peroxidare, la care se alterează gustul şi mirosul (râncezire)
 Săruri (săpunuri cu proprietăţi tensioactive) Amide (sfingolipide) AG nesaturaţi adiţionează la nivelul dublei legături – halogeni (Br2; Cl2), tiocianatul (SCN)2, gruparea hidroxil. AG nesaturaţi suferă procesul de peroxidare, la care se alterează gustul şi mirosul (râncezire)")
15
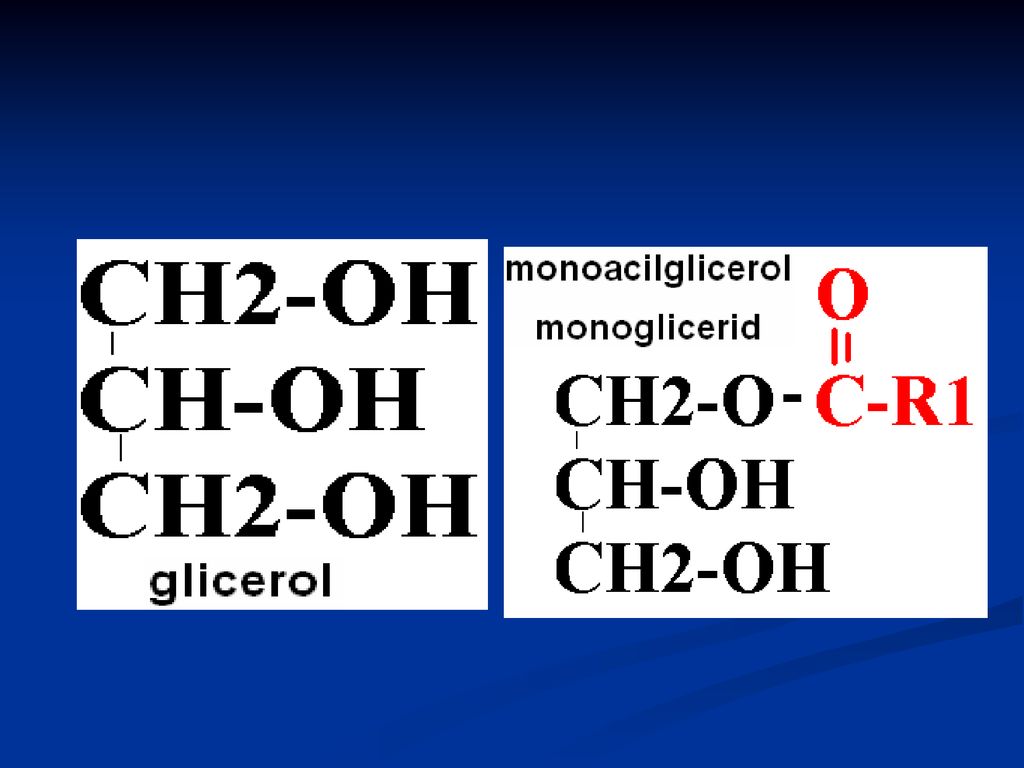
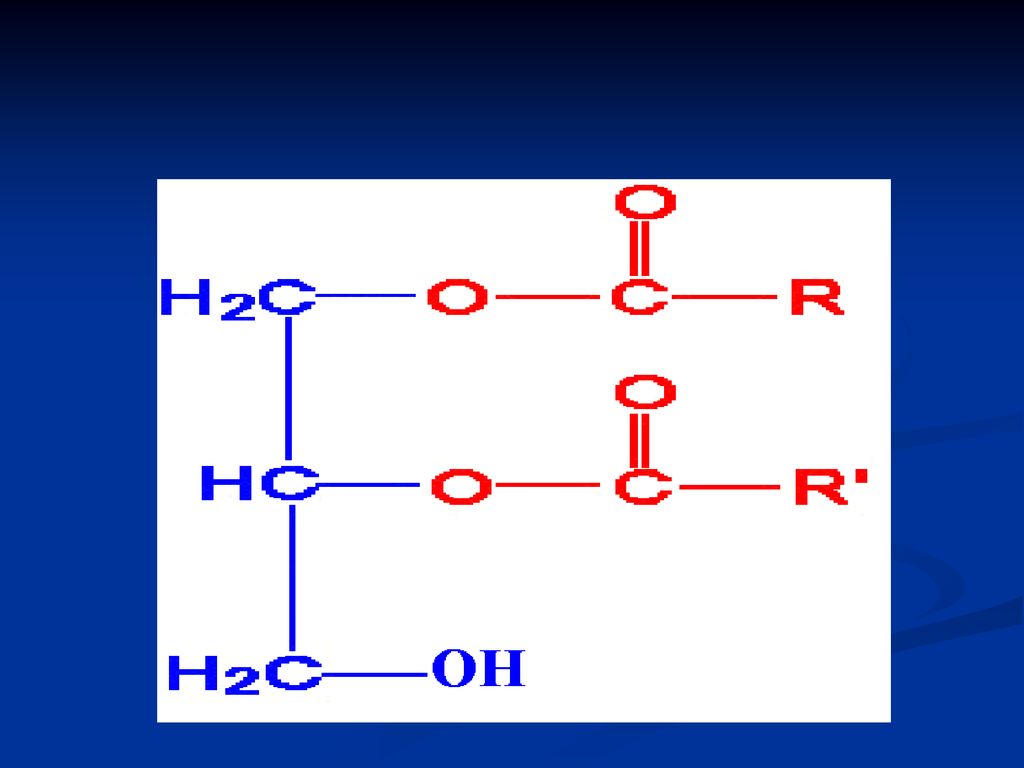
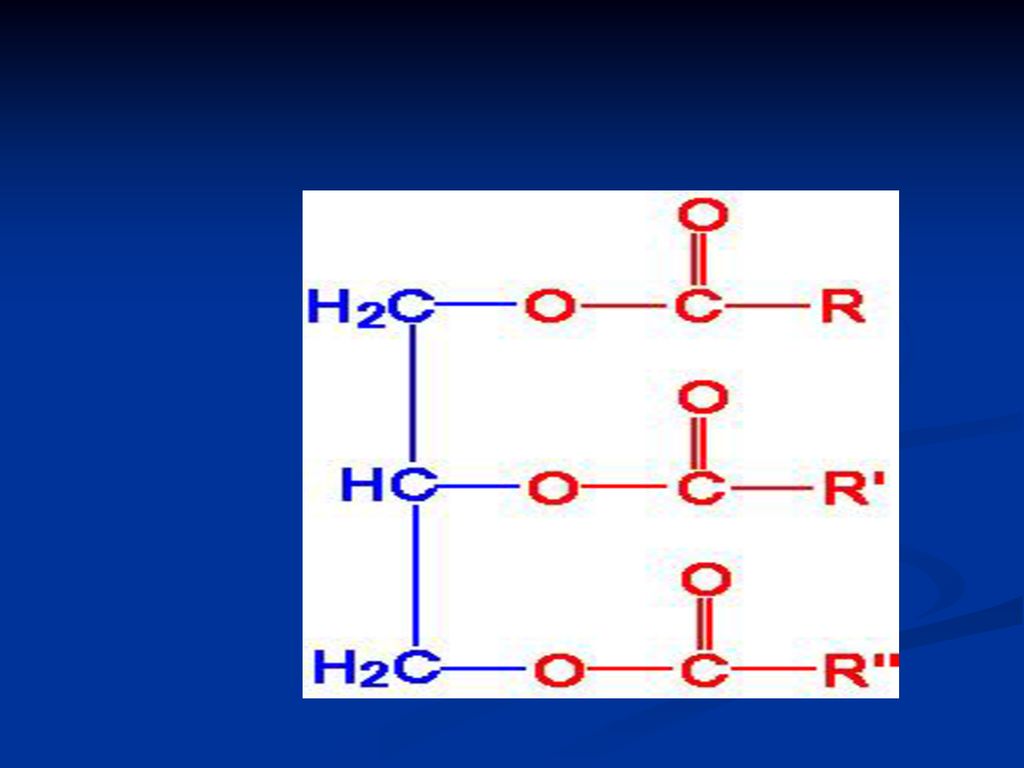
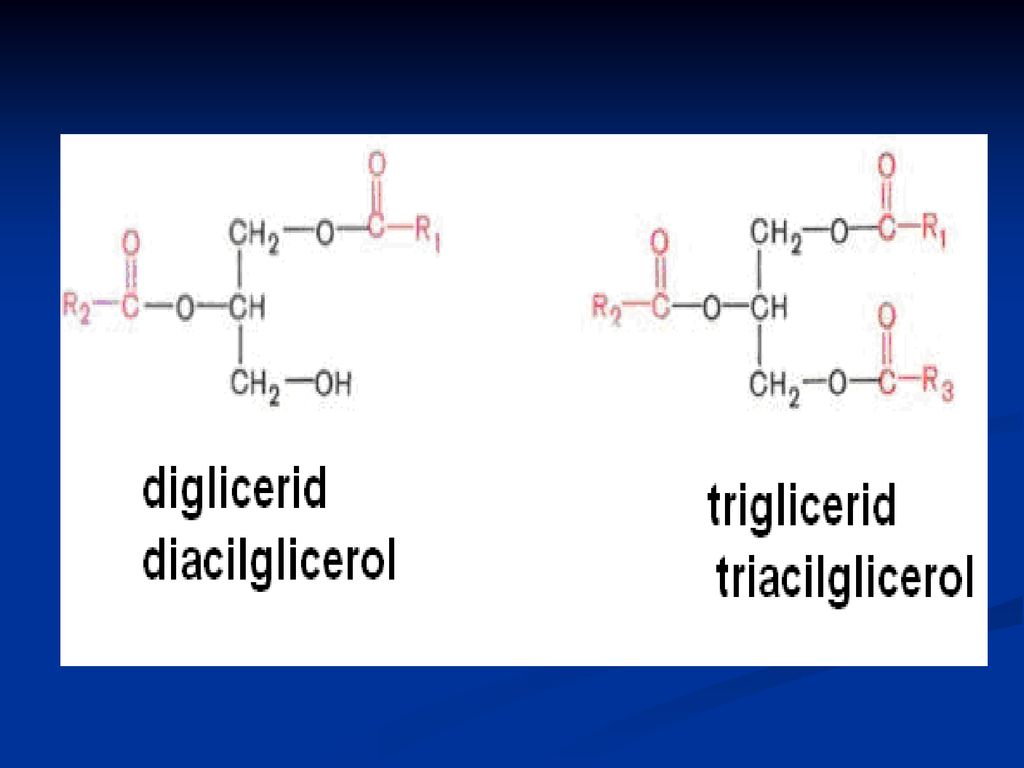
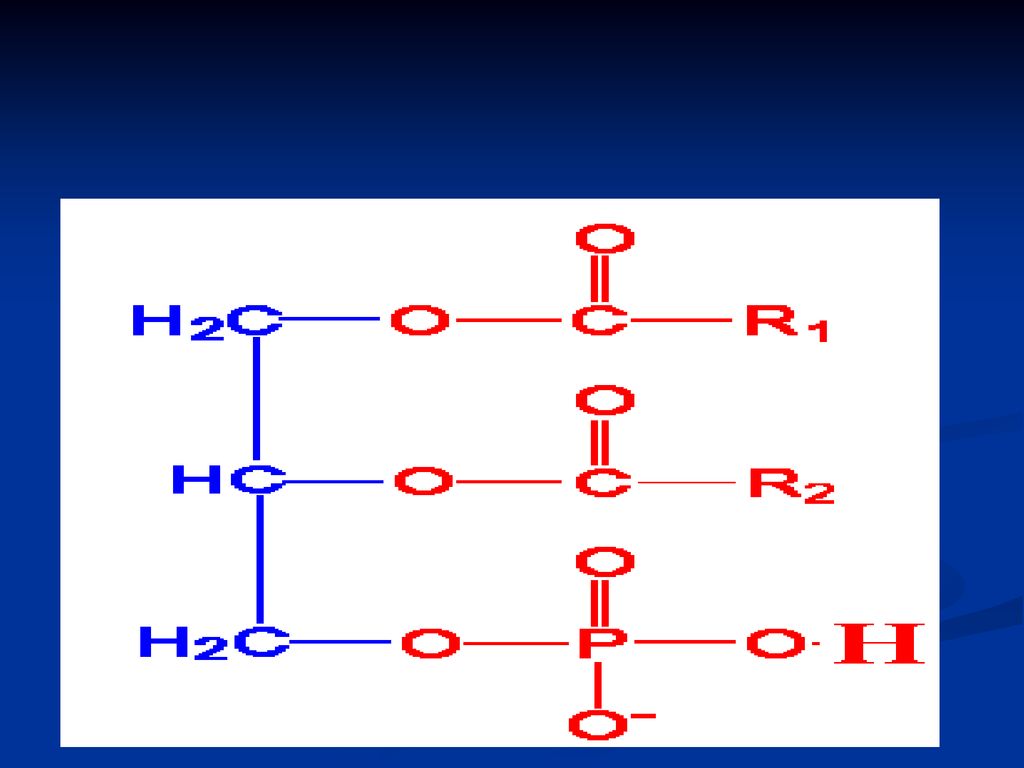
GLICERIDE ( acilgliceroli, grăsimi neutre)
Sunt esteri ai glicerolului cu AG După nr grupărilor alcoolice esterificate din glicerol deosebim: mono-; di- şi tri gliceride

20
Rolul Tg Constituie grăsimea de rezervă din ţesuturi Rol energetic
Se găsesc în LP plasmatice Izolator La oxidarea 1g se elimină 9kcal Tg din ţesutul uman cuprind următorii AG: Oleic-45%, palmitic-25%, linoleic – 8%, palmitooleic – 7%, stearic- 7% şi alţii -7%

21
Proprietăţile Tg Sunt determinate de natura şi numărul de AG constituienţi Insolubile în apă, solubile în solvenţi organici Tg ce conţin AG saturaţi – solide(grăsimi, unt)- de origine animală Tg ce conţin AG nesaturaţi – consistenţă lichidă (uleiurile vegetale) Prin hidroliza enzimatică (se produce în intestin sub acţiunea lipazei) se scindează în glicerol şi 3AG Prin hidroliza alcalină (KOH, NaOH) –glicerol+săpunuri Prezenţa AG nesaturaţi – proprietatea de a adiţiona halogenii la dublele legături Sub acţiunea luminii, căldurii – se autooxidează
- de origine animală. Tg ce conţin AG nesaturaţi – consistenţă lichidă (uleiurile vegetale) Prin hidroliza enzimatică (se produce în intestin sub acţiunea lipazei) se scindează în glicerol şi 3AG. Prin hidroliza alcalină (KOH, NaOH) –glicerol+săpunuri. Prezenţa AG nesaturaţi – proprietatea de a adiţiona halogenii la dublele legături. Sub acţiunea luminii, căldurii – se autooxidează.")
22
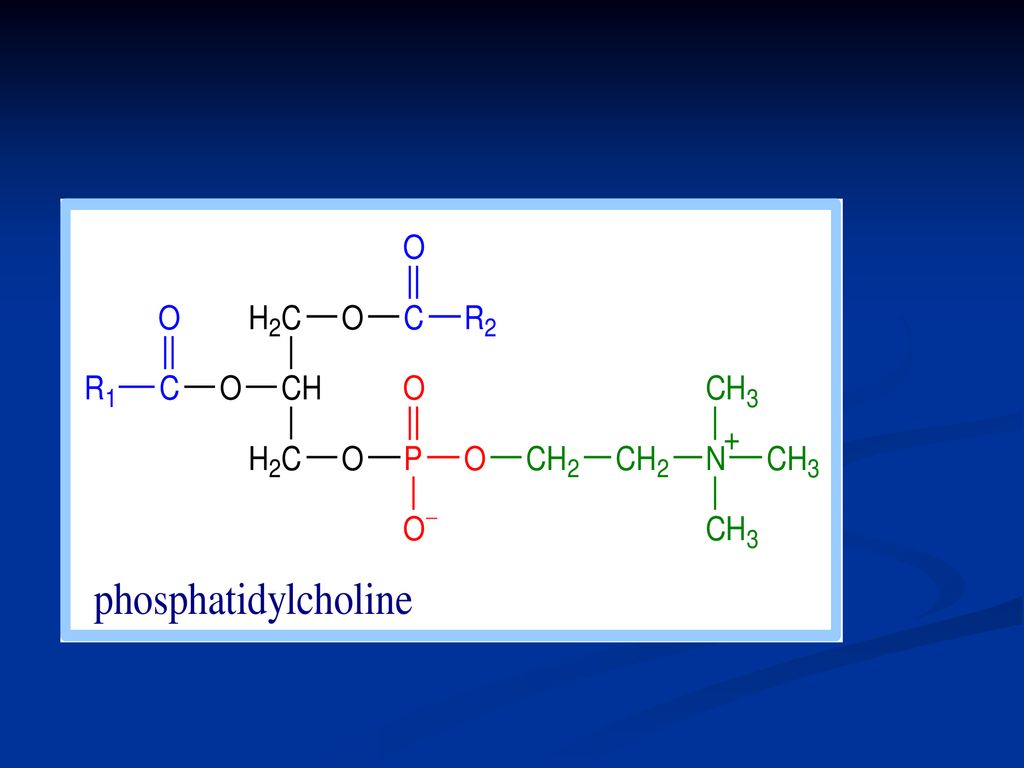
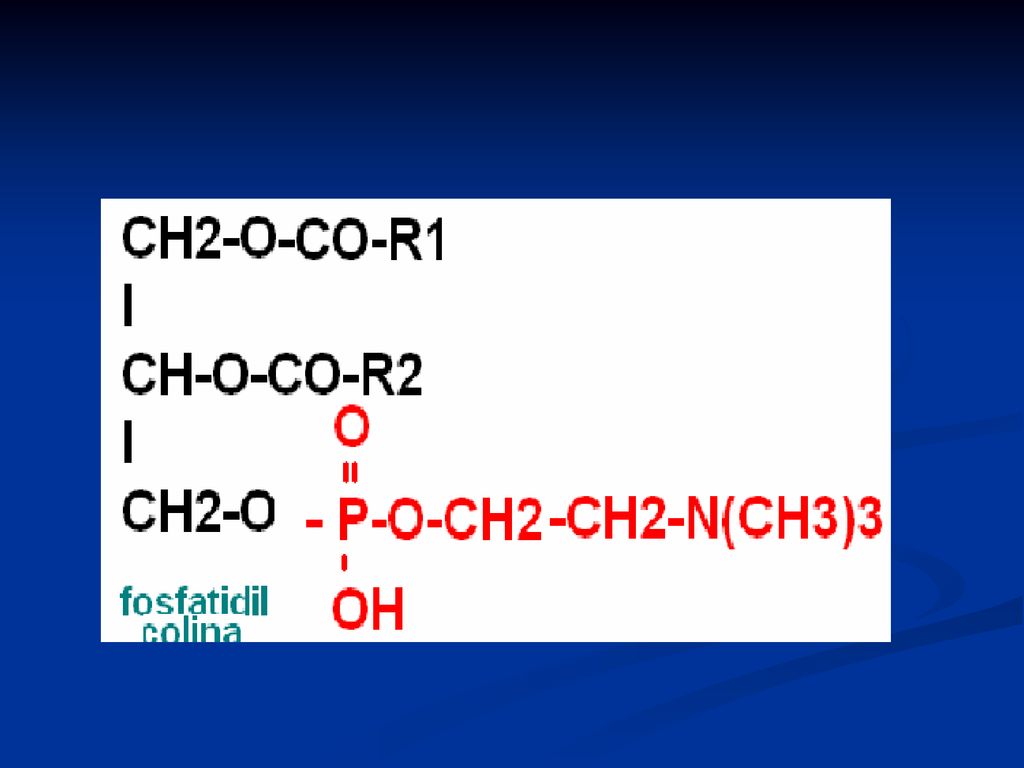
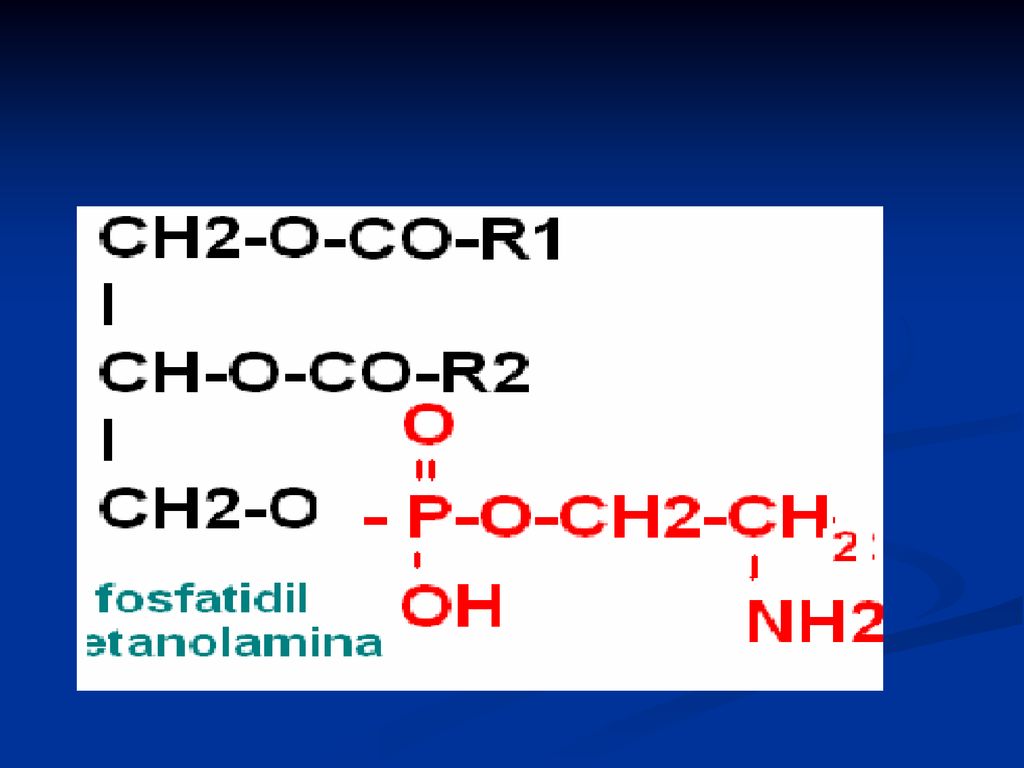
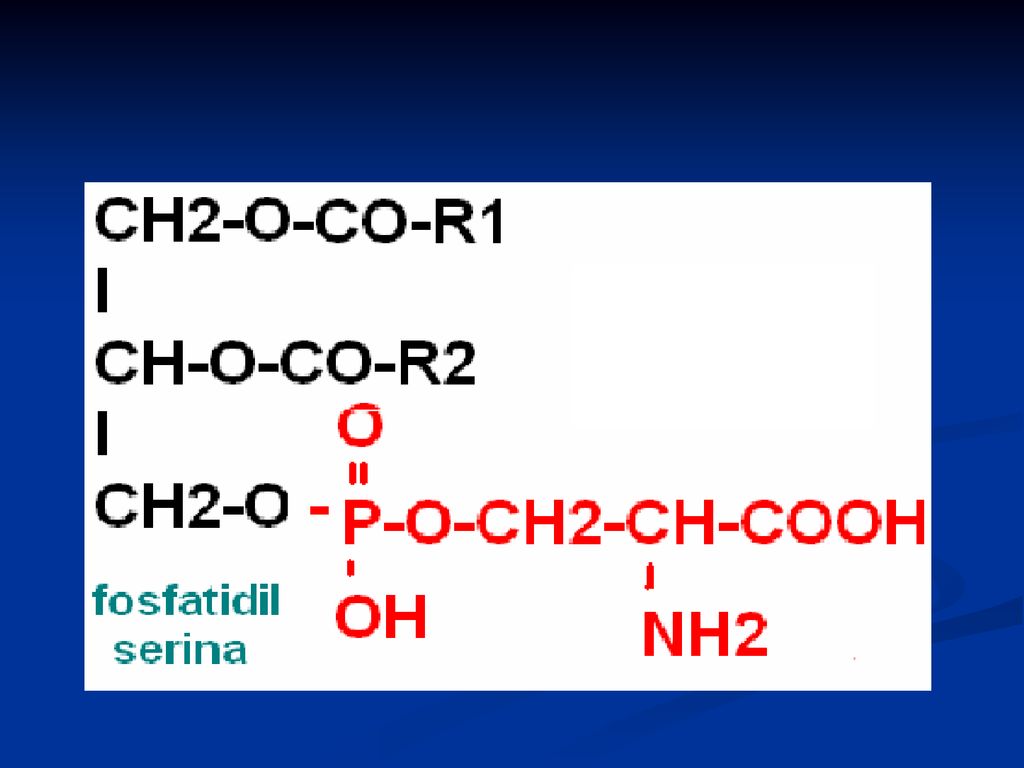
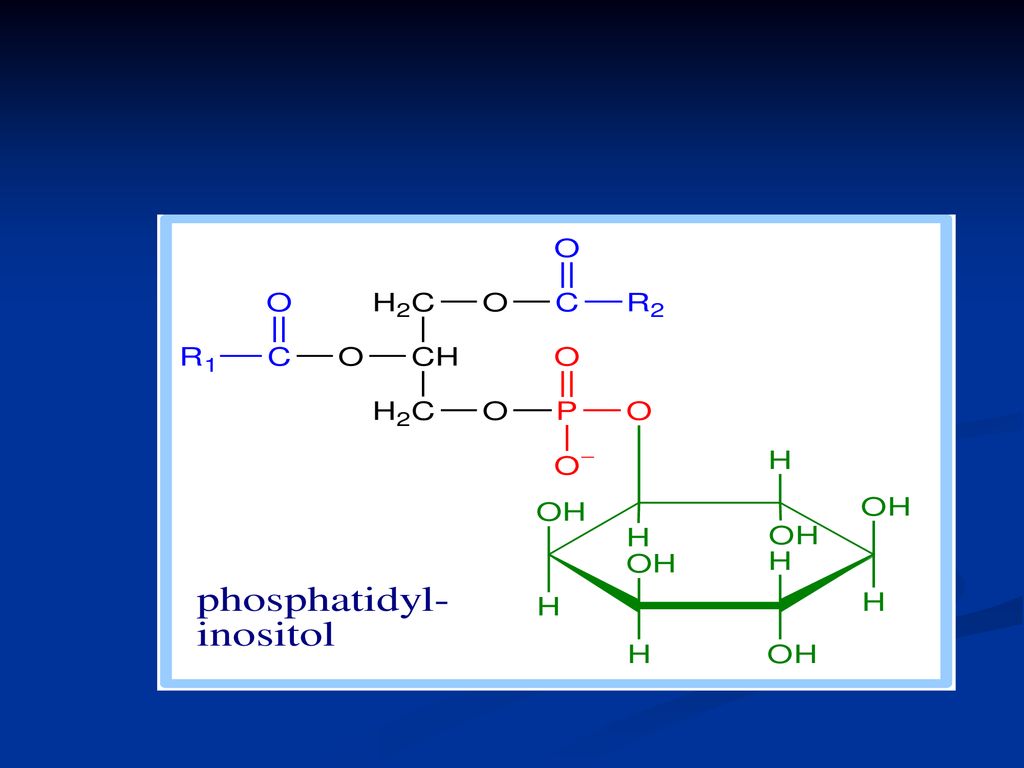
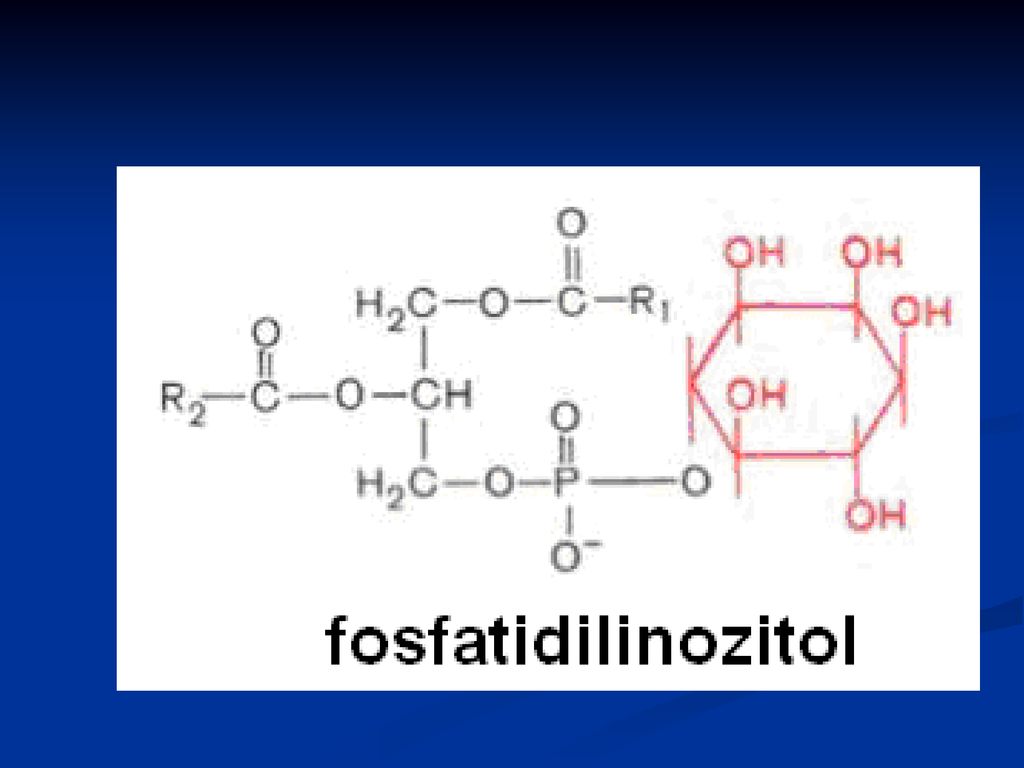
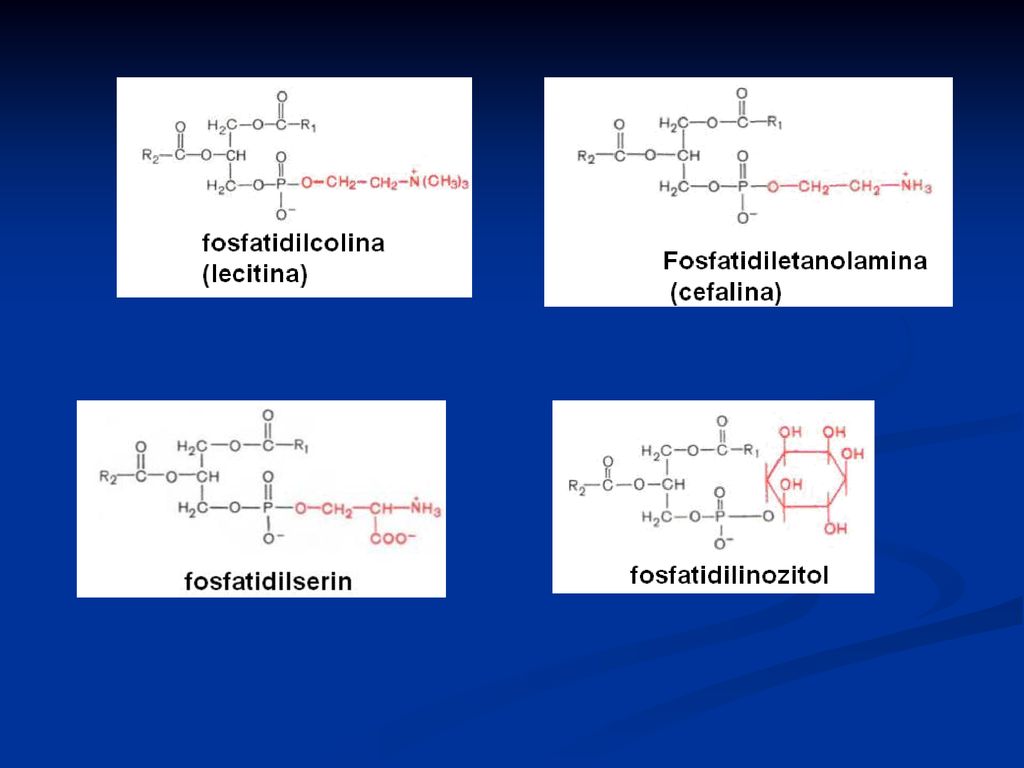
Fosfatidele Derivaţi ai acidului fosfatidic Reprezentanţii:
fosfatidilcolina (lecitina) Fosfatidiletanolamina (cefalina) Fosfatidilserina Fosfatidilinozitol
 Fosfatidiletanolamina (cefalina) Fosfatidilserina. Fosfatidilinozitol.")
30
Rolul: Structural Fosfatidilcolina cu 2 resturi de palmitil este componentul principal al surfactantului pulmonar, ce acoperă alveolele şi împedică colapsul la expiraţie Fosfatidiletanolamina – sunt abundente în ţesutul nervos Fosfatidilinozitolul- rol în procesul de transmitere a semnalelor extracelulare

32
Proprietăţile Caracter amfipatic (lecitinile şi cefalinele au structură bipolară: prezenţa resturilor acil – hidrofobe şi a sarcinilor electrice: sarcina negativă – restul a fosforic; sarcină pozitivă – gr alcool)
")
33
Proprietăţile FL 2. Proprietăţi tensioactive puternice
3. În apă se dizolvă formând agregate micelii, alcătuite din 2 straturi bilipidice 4. Sunt neutre la electricitate 5. Fosfatidilserina şi fosfatidilinozitolul au sarcină negativă

34
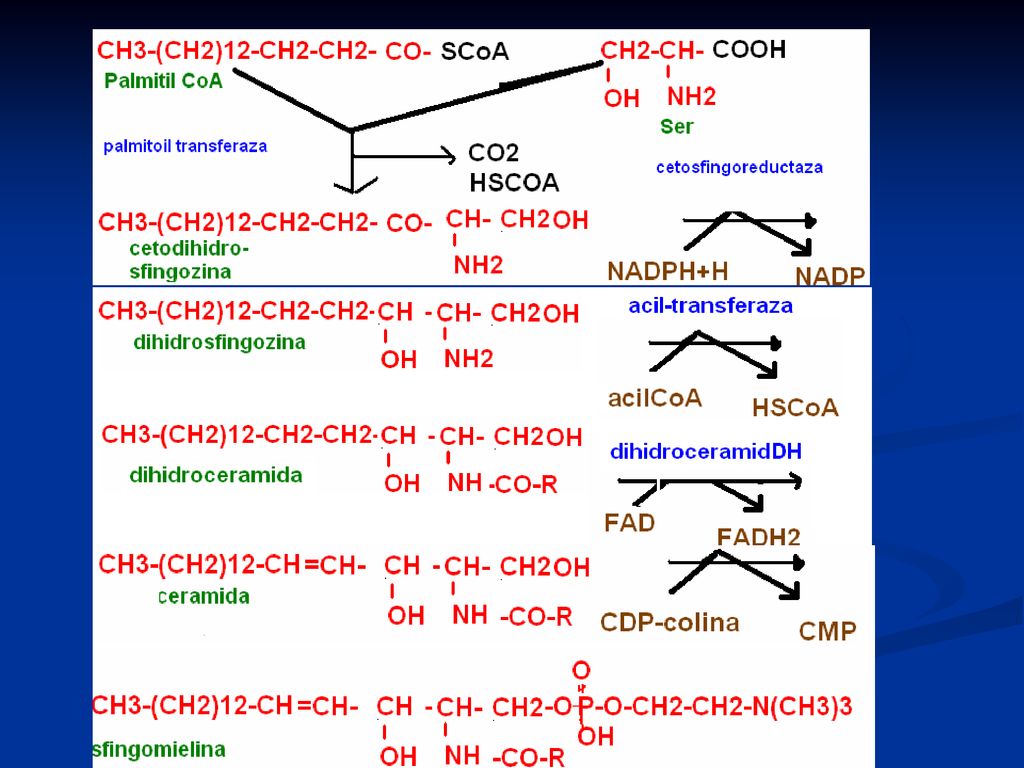
Sfingolipidele Derivaţi ai sfingozinei
Se găsesc cu predelecţie în ţesutul cerebral, splină şi eritrocite Rolul: Sunt surse energetice (datorită AG) Componente ale membranei celulare (nervoasă) Participă la coagularea sângelui Sunt transportori de e şi componente ale mitocondriilor, unde au loc procesele de O/R Rol în fiziologia SNC Proprietăţi; amfioni, au caracter amfipatic 2 categorii: sfingomielina şi glicosfingolipidele
 Componente ale membranei celulare (nervoasă) Participă la coagularea sângelui. Sunt transportori de e şi componente ale mitocondriilor, unde au loc procesele de O/R. Rol în fiziologia SNC. Proprietăţi; amfioni, au caracter amfipatic. 2 categorii: sfingomielina şi glicosfingolipidele.")
35
Sfingozina Sfingozina +AG= ceramidă

36
Sfingomielina: ceramida+ rest de fosforilcolină

37
Sfingomielina Sfingozina +AG = ceramidă Ceramida+ rest de
fosforilcolină = sfingomielina

38
Glicolipide (Glicosfingolipidele)
- includ ceramida legată glicozidic de monozaharide sau oligozaharide (nu cuprind fosfor) Cerebrozide – se află abudent în substanţa albă a creierului; în nervi. Proprietăţi:nu au sarcină electrică; au caracter amfipatic Structură: ceramida (AG-24C)+βgGal (mai rarGl)
 Cerebrozide – se află abudent în substanţa albă a creierului; în nervi. Proprietăţi:nu au sarcină electrică; au caracter amfipatic. Structură: ceramida (AG-24C)+βgGal (mai rarGl)")
39
Proprietăţi: la PH=7 au sarcină negativă
2. Gangliozide – sunt prezente în toate ţesuturile în cantităţi mici, mai abudent în substanţa albă a creierului; în nervi Structură: ceramida (AG-stearic)+oligozaharid (Gal+Gl+Nacetilglucozamină)+ 1,2sau3 resturi de a sialic – acetilneuraminic mono-; di-; tri- sialogangliozid Proprietăţi: la PH=7 au sarcină negativă
+oligozaharid (Gal+Gl+Nacetilglucozamină)+ 1,2sau3 resturi de a sialic – acetilneuraminic mono-; di-; tri- sialogangliozid. Proprietăţi: la PH=7 au sarcină negativă.")
40
Steroizii Sunt derivaţi a ciclului pentanperhidrofenantren (steranului –C17)
")
41
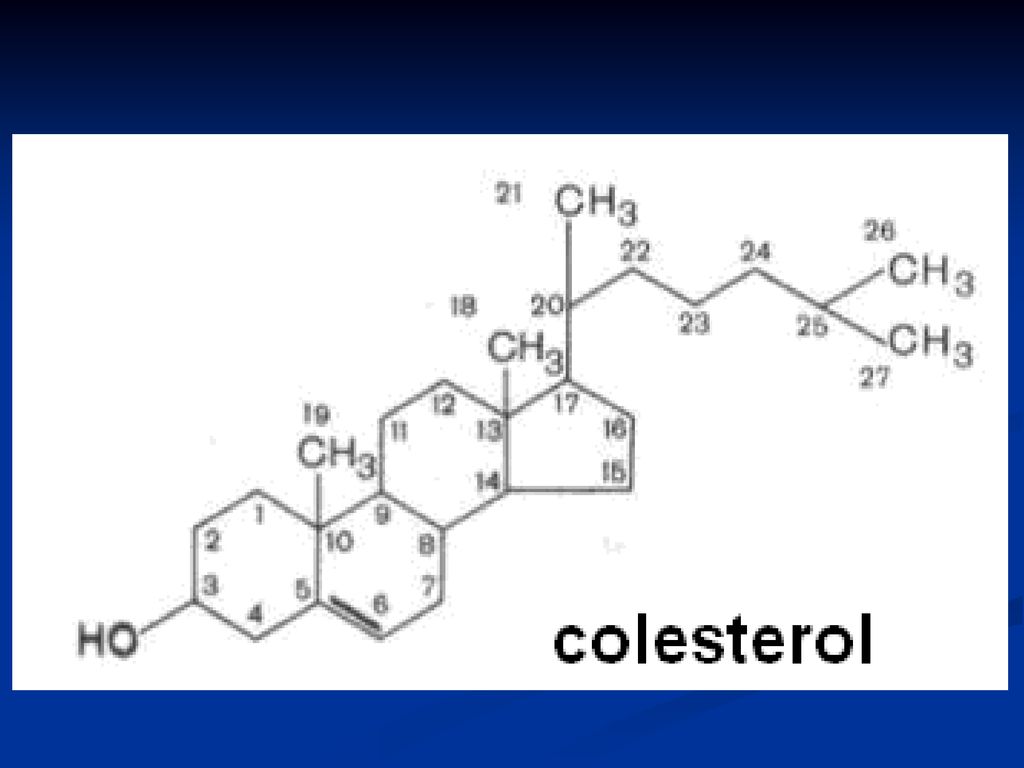
Colesterolul C27 Origine animală (lipseşte în plante)
Solid, alb, insolubil în apă Rol structural Precursorul tuturor compuşilor steroidici (provitaminei D, acizilor biliari, hormonilor steroizi) 2 forme: esterificat şi neesterificat
 2 forme: esterificat şi neesterificat.")
43
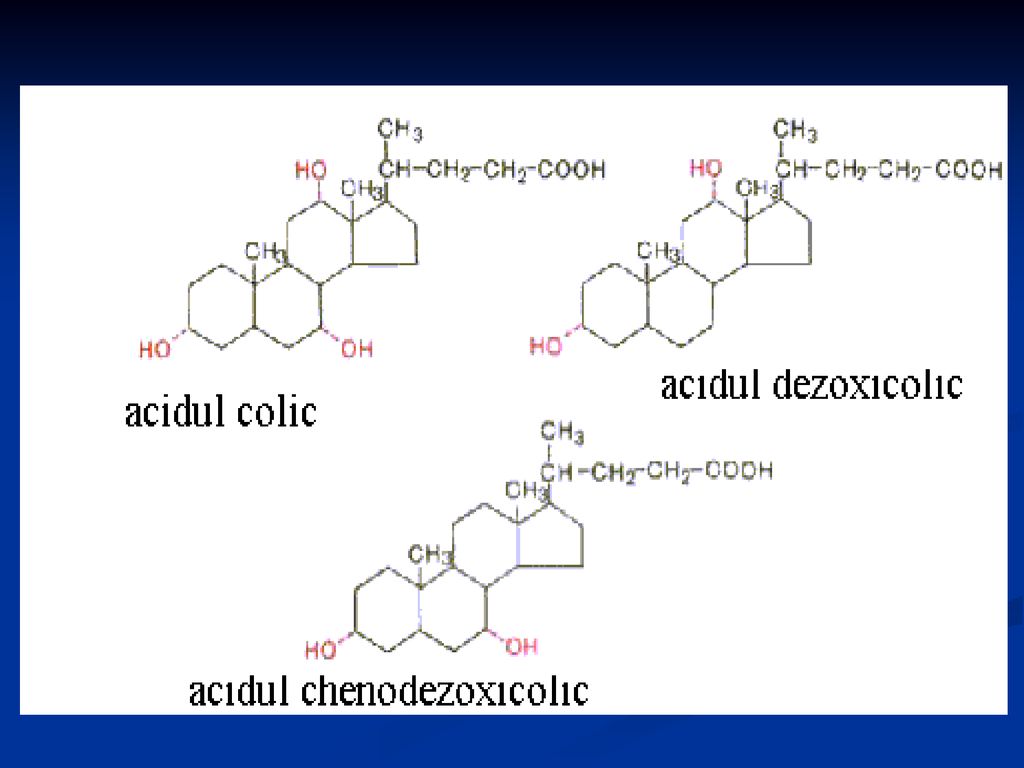
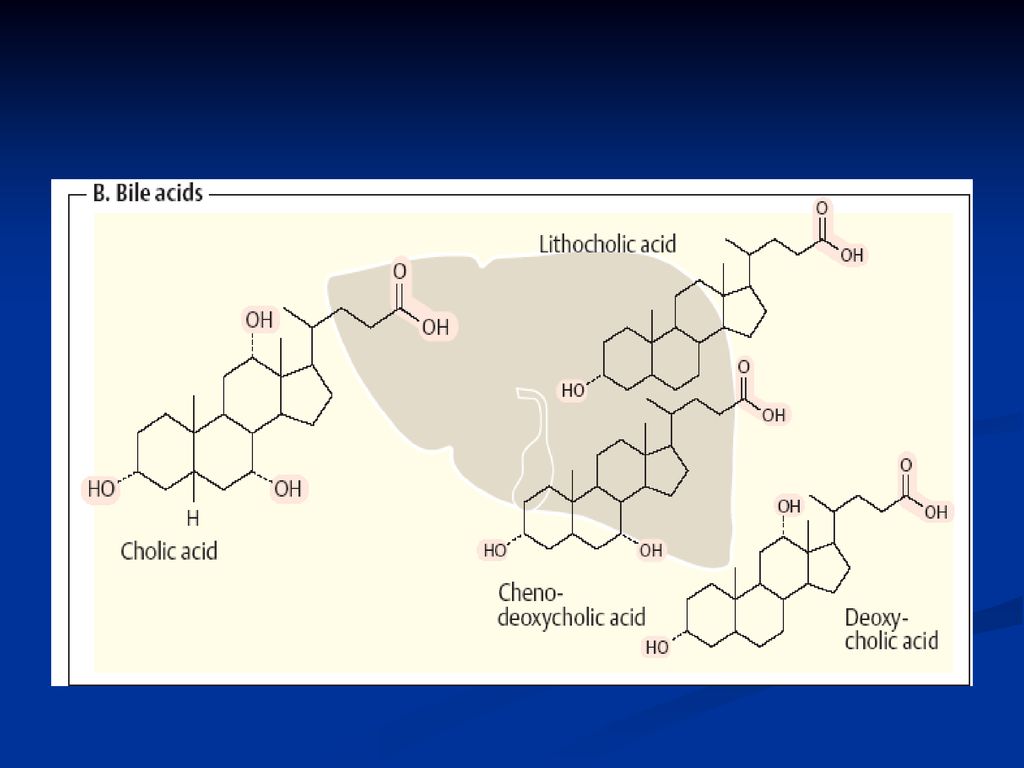
Acizii biliari Sunt componenţi ai bilei
Structură: steroizi C24: colic, chenodezoxicolic, dezoxicolic Rolul: proprietăţi de solubilitate (micele în apă şi au capacitatea de a solubiliza alte lipide) Transport (AG) Emulgatori Activează lipaza pancreatică La PH alcaln se găsesc sub formă de săruri biliare (conjugaţi cu glicina sau taurina
 Transport (AG) Emulgatori. Activează lipaza pancreatică. La PH alcaln se găsesc sub formă de săruri biliare (conjugaţi cu glicina sau taurina.")
46
Membranele biologice Structuri superorganizate, posedînd constituienţii de bază - proteinele şi lipidele. despart celula de mediul estern şi separă mediul ei intern în compartimente reprezintă o barieră de permeabilitate selectivă Conţin pompe moleculare specifice şi canale (sisteme de transport) Sunt flexibile, labile, permanent se reînnoiesc sunt elastice şi dure la deformare
 Sunt flexibile, labile, permanent se reînnoiesc. sunt elastice şi dure la deformare.")
47
Funcţiile biologice ale membranelor:
de separare (de barieră) de integrare sau asociere – asociază procesele biochimice izolate în structuri integre de transport (pasiv, activ); Osmotică – de concentrarea a substanţelor între spaţiile intra- şi extracelulare Electrică – repartizarea neuniformă a sarcinilor pe ambele părţi ce duce la apariţia diferenţei de potenţial electric;
 de integrare sau asociere – asociază procesele biochimice izolate în structuri integre. de transport (pasiv, activ); Osmotică – de concentrarea a substanţelor între spaţiile intra- şi extracelulare. Electrică – repartizarea neuniformă a sarcinilor pe ambele părţi ce duce la apariţia diferenţei de potenţial electric;")
48
de transformare a energiei – asigură transformarea energiei electrice, osmotice în energia chimică -ATP; de recepţie – prin intermediul receptorilor ele reglează schimbul de informaţie între celule şi mediul extern; reglatoare; metabolică – participă la formarea AMPc; GMPc; enzimele membranare – în diverse transformări metabolice; Antigenică – glicoproteinele membranare determină capacitatea de formare a anticorpilor;

49
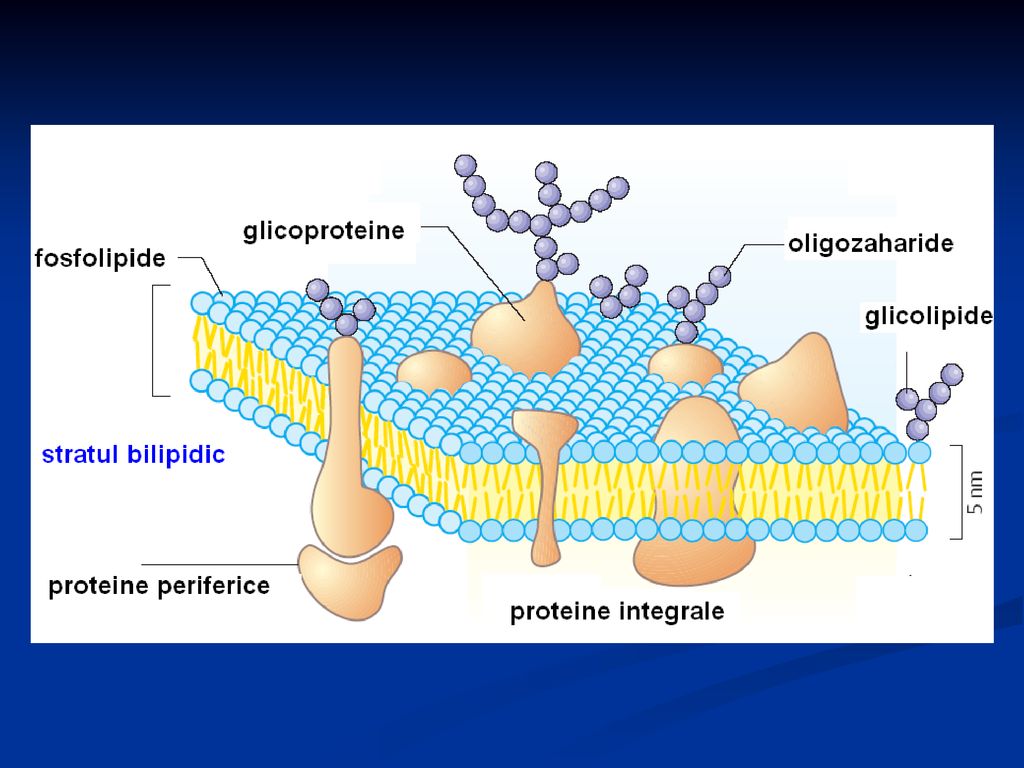
Structura MB Componenţi de bază: lipidele şi proteinele (1:4 pînă la 4:1) Glucidele Lipidele: conţin grupe hidrofobe (nepolară) şi hidrofile (polară) – în soluţii apoase formează stratul bilipidic Fosfolipidele şi glicolipidele – constituienţii cheie ai membranelor – îndeplinesc funcţia de barieră a permeabilităţii şi asigură crearea compartimentelor unice
 şi hidrofile (polară) – în soluţii apoase formează stratul bilipidic. Fosfolipidele şi glicolipidele – constituienţii cheie ai membranelor – îndeplinesc funcţia de barieră a permeabilităţii şi asigură crearea compartimentelor unice.")
50
Capătul polar al fosfolipidelor este situate la suprafateţele membranei, iar capetele nepolare sînt îndreptate în interior. Moleculele colesterolului se alătură cu grupele sale hidrofile de grupele polare ale fosfolipidelor, iar ciclul steran pătrunde în grosimea stratului bilipidic între catenele hidrocarburilor fosfolipidelor.

51
Proteinele – asigură transportul, transmiterea informaţiei, transformările de energie.
Deosebim: proteine periferice şi integrale Proteinele periferice sunt legate de membrane prin forţe electrostatice şi de hidrogen Proteinele integrale - amfipatice, conţin AA nepolari – formează cu lipidele membranei legături hidrofobe

52
Se cunosc următoarele variante de aranjare a proteinelor.
superficial parţial cufundate în stratul lipidic cufundate definitiv în stratul bilipidic străbat de la un capăt la altul membrana

53
Glucidele – sub formă de glicolipide şi glicoproteine
Sunt situate pe suprafaţa membranei, dar nu în faza hidrocarbonată Favorizează menţinerea asimetriei membranelor biologice Au funcţia de identificare intercelulară, de detectare a celulelor străine sistemului imun Determină transportul proteinelor membranare în locusurile necesare

55
Proprietăţile membranelor biologice.
fluiditatea. Starea în care se află lipidele membranare se numeşte fluidcristalină, deoarece pătura lipidică este lichidă, însă în ea se conţin sectoare compacte. Reglatorul fluidităţii – AG saturaţi, nesaturaţi şi Col Situîndu-se între catenele acil, Col: evită cristalizarea şi fuziunea lor, blochează mobilitatea catenelor acil, micşorînd fluiditatea MC Col menţine fluiditatea la un nivel mediu

56
Proprietăţile membranelor biologice.
Asimetria: 1. repartizarea neuniformă a lipidelor şi proteinelor pe suprafaţa internă şi externă a MB Ex.: membrana eritrocitului: Partea internă- fosfatidiletanolamină şi fosfatidilserină Partea externă – fosfatidilcolină şi sfingomielină Asigură orientarea proteinelor membranare în bistrat 2. La suprafaţa externă este prezentă componenta glucidică (glicocalixul), pe cînd pe partea internă ele practic lipsesc. 3. Sistemele de transport din membrană funcţionează într-o singură direcţie (Na-K-ATP-aza)
, pe cînd pe partea internă ele practic lipsesc. 3. Sistemele de transport din membrană funcţionează într-o singură direcţie (Na-K-ATP-aza)")
57
Proprietăţile membranelor biologice.
Mobilitatea Moleculele lipidelor membranei se află în mişcare continuă. Sunt posibile cîteva tipuri de mişcare a moleculelor: difuzie de rotaţie , transversală şi difuzie laterală. Proteinele membranare tot posedă mobilitate. Ele parcă plutesc în pătura lipidică, deplasîndu–se în procesul difuziunii laterale.

58
Proprietăţile membranelor biologice.
permeabilitatea selectivă. Membrana biologică este permeabilă pentru substanţele lipofile (steroizi ş.a.) şi moleculele nepolare mici. Substanţele polare (ionii, aminoacizii, glucoza etc.) nu pot trece membrana prin difuziune simplă, ele necesită anumite sisteme de transport.
 şi moleculele nepolare mici. Substanţele polare (ionii, aminoacizii, glucoza etc.) nu pot trece membrana prin difuziune simplă, ele necesită anumite sisteme de transport.")
59
Digestia şi absorbţia lipidelor Metabolismul lipoproteinelor

60
Obiectivele Importanţa lipidelor în alimentaţie.
Digestia şi absorbţia lipidelor în tractul gastrointestinal. Acizii biliari – clasificarea, structura, funcţiile lor. Metabolismul acizilor biliari ( noţiuni generale). Resintaza lipidelor în enterocite. Soarta lipidelor resintetizate. Metabolismul LPP.
. Resintaza lipidelor în enterocite. Soarta lipidelor resintetizate. Metabolismul LPP.")
61
Importanţa lipidelor în alimentaţie.
Aportul alimentar de lipide necesar zilnic este în mediu de 80g, incluzând grăsimi atât de origine animală cât şi de origine vegetală. Principalele lipide ale raţiei alimentare sunt: Tg; FL; Col liber şi esterificat. Lipidele alimentare sunt sursa AG indespensabili – linolic şi linolenic. Funcţionarea normală a organismului necesită un consum minim obligatoriu de vitamine liposolubile ( A,D,E,K )
")
62
Digestia grăsimilor în TGI
Digestia şi absorbţia lipidelor alimentare necesită: 1. prezenţa AB (compuşii majori ai bilei) care conduc la: a. emulsionarea lipidelor alimentare, b. activarea enzimelor lipolitice; c. absorbţia produselor finale ale digestiei. 2. pH alcalin -bicarbonaţii sucului pancreatic crează pH-ul optim pentru enzimele lipolitice. 3. Prezenţa enzimelor lipolitice: lipaza, fosfolipazele, colesterolesteraza, sfingomielinaza şi ceramidaza.
 care conduc la: a. emulsionarea lipidelor alimentare, b. activarea enzimelor lipolitice; c. absorbţia produselor finale ale digestiei. 2. pH alcalin -bicarbonaţii sucului pancreatic crează pH-ul optim pentru enzimele lipolitice. 3. Prezenţa enzimelor lipolitice: lipaza, fosfolipazele, colesterolesteraza, sfingomielinaza şi ceramidaza.")
63
În cavitatea bucală – Tg nu se supun modificărilor deoarece saliva nu conţine E digestiei lor
Excepţie: sugari, lipaza lingvală – digestează Tg în poziţia 3 – 1,2 diglicerid În stomac – digestia are loc doar la copiii sugari (PH sucului gastric = 5) – sub acţiunea lipazei gastrice se digestează Tg din lapte
 – sub acţiunea lipazei gastrice se digestează Tg din lapte.")
64
Digestia grăsimilor în TGI
la adulţi are loc în intestinul subţire. Prezenţa E lipolitice Condiţii pentru emulsionarea lipidelor PH optim pentru acţiunea E (neutru sau slab alcalin)
")
65
În intestin - chimul din stomac este neutralizat de bicarbonaţii sucului pancreatic şi intestinal.
Grăsimile se supun emulsionării sub acţiunea sărurilor AB şi E lipolitice. AB se amplasează la suprafaţa grăsime-apă împedică separarea acestor 2 faze. Peristaltismul intestinului ajută la fărîmiţarea picăturilor mici de grăsime, iar sărurile AB le menţin în stare suspendată, împedicând contopirea picăturilor mici de grăsime

66
Digestia TG lipaza pancreatică: Este o glicoproteidă
Se activează sub acţiunea colipazei şi AB PH=8-9 are specificitate pentru legăturile 1,3.

67
sub acţiunea izomerazei (transferă restul acil din poziţia β2 în α1) - 1/3 din 2 monoglicerid trece în 1 monoglicerid 1 monogliceridul sub acţiunea lipazei se scindează la AG şi glicerol

68
Digestia fosfogliceridelor
fosfolipazele pancreatice (A1, A2, C, D) pînă la glicerol, acizi graşi, acid fosforic şi compuşi azotaţi
 pînă la glicerol, acizi graşi, acid fosforic şi compuşi azotaţi.")
69
Digestia fosfogliceridelor
Cea mai activă este fosfolipaza A2 (se activează de tripsină, Ca; AB) fosfatidilcolina lizolecitina (proprietăţi detergente puternice; participă la solubilizarea lipidelor în intestin) Sub acţiunea lizofosfolipazei - lizolecitina se scindează în glicerolfosfocolină şi AG
 fosfatidilcolina lizolecitina (proprietăţi detergente puternice; participă la solubilizarea lipidelor în intestin) Sub acţiunea lizofosfolipazei - lizolecitina se scindează în glicerolfosfocolină şi AG.")
70
Digestia sfingomielinelor
Sfingomielinaza: singozină+AG+fosforilcolină Ceramidaza – degradează legătura N acilică

71
Digestia Col Colesterolesteraza pancreatică (activată de AB) scindează Col esterificat în Col şi AG
 scindează Col esterificat în Col şi AG")
72
Absorbţia lipidelor Produsele finale ale digestiei lipidelor sunt: 2 monogliceridul; glicerolul; AG; sfingozina; a. fosforic, colina, etanolamina; Col. sunt absorbite la nivelul intestinului subţire prin difuzie simplă sau pinocitoză micelară (sub formă de micele (picături lipidice mici), la formarea cărora participă AB). Prin difuzie simplă sunt transportaţi: AG cu catena scurtă; glicerolul, colina, etanolamina Acidul fosforic – sub formă de sare de Na sau K AG cu catena lungă, monoacilglicerolul, Col se absorb cu ajutorul AB
, la formarea cărora participă AB). Prin difuzie simplă sunt transportaţi: AG cu catena scurtă; glicerolul, colina, etanolamina. Acidul fosforic – sub formă de sare de Na sau K. AG cu catena lungă, monoacilglicerolul, Col se absorb cu ajutorul AB.")
73
Ciclul entero-hepatic
AG +AB formează o soluţie micelară şi pătrund în spaţiile intervilozitare de la nivelul jejunului proximal unde AG se absorb. Sărurile AB rămîn în lumen, participînd la solubilizarea şi transportul altor lipide. Abia în porţiunea distală a ileonului sărurile AB se absorb printr-un mecanism activ. Prin sistemul portal trec în ficat (se reînnoiesc)– bila --- intestin - circulaţia entero-hepatică a acizilor biliari.
– bila --- intestin - circulaţia entero-hepatică a acizilor biliari.")
74
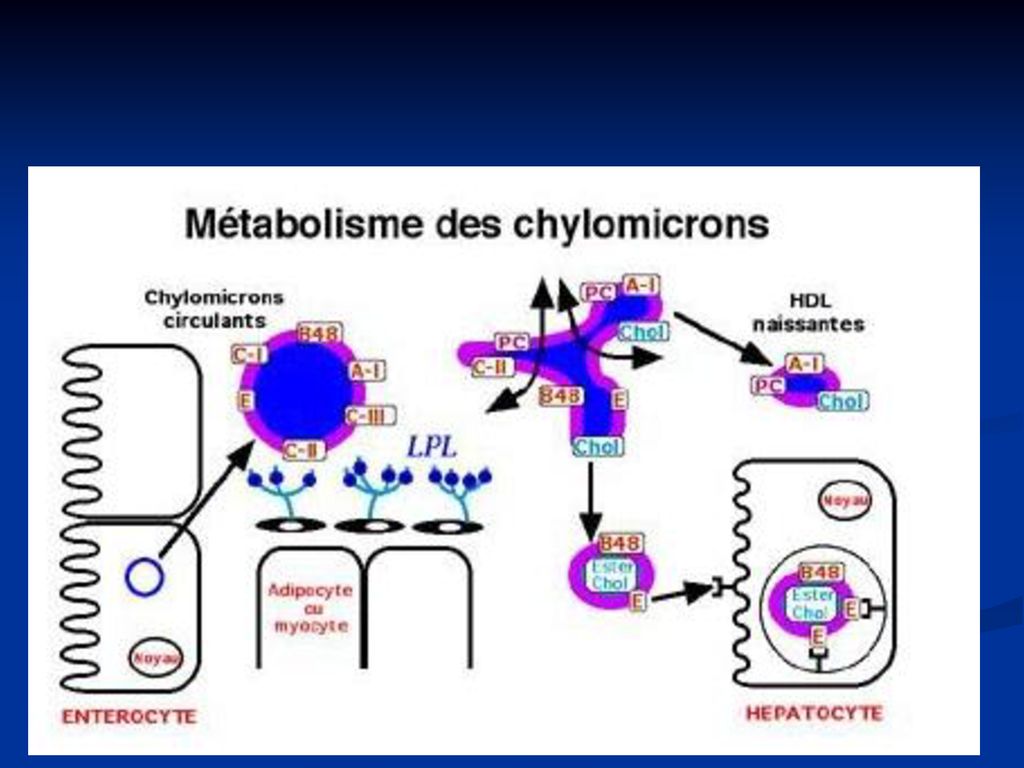
Resinteza lipidelor Din substanţele ce se absorb din lumenul intestinului în eterocite – are loc resinteza lipidelor specifice organismului uman: Tg, FL, Col esterificat Rolul: lipidele digestive se deosebesc după structură, proprietăţile fizico-chimice de lipidele specifice. În enterocite se formează lipide specifice organismului uman. Moleculele lipidelor reconstituite împreună cu cantităţi mici de proteină sunt încorporate în chilomicroni (CM) CM sunt secretaţi în vasele limfatice ce drenează intestinul şi la nivelul canalului toracic trec în plasmă.
 CM sunt secretaţi în vasele limfatice ce drenează intestinul şi la nivelul canalului toracic trec în plasmă.")
75
LIPOPROTEINELE (LP) LP- sînt complexe lipoproteice, alcătuite din componente lipidice şi proteice. Componentele lipidice: Tg, FL, Col, colesteride Componentele proteice sînt denumite apolipoproteine (Apo). Rolul LP Transportul lipidelor exogene şi endogene Participă la păstrarea compoziţiei lipidice a membranelor Reglează procese metabolice celulare Rolul Apo: componente amfipatice a LP Oferă situsuri de recunoaştere pentru R de pe suprafaţa celulelor Sunt activatori sau inhibitori ai E ce participă la metabolismul lor
. Rolul LP. Transportul lipidelor exogene şi endogene. Participă la păstrarea compoziţiei lipidice a membranelor. Reglează procese metabolice celulare. Rolul Apo: componente amfipatice a LP. Oferă situsuri de recunoaştere pentru R de pe suprafaţa celulelor. Sunt activatori sau inhibitori ai E ce participă la metabolismul lor.")
76
Structura LP LP au o structură comună, lipidele nepolare (TG şi esterii Col) formează un miez hidrofob, iar lipidele amfipatice (FL, Col) şi Apo alcătuesc învelişul hidrofil. LP cuprind şi cantităţi mici de glucide (sub formă de glicoproteine).
 formează un miez hidrofob, iar lipidele amfipatice (FL, Col) şi Apo alcătuesc învelişul hidrofil. LP cuprind şi cantităţi mici de glucide (sub formă de glicoproteine).")
77
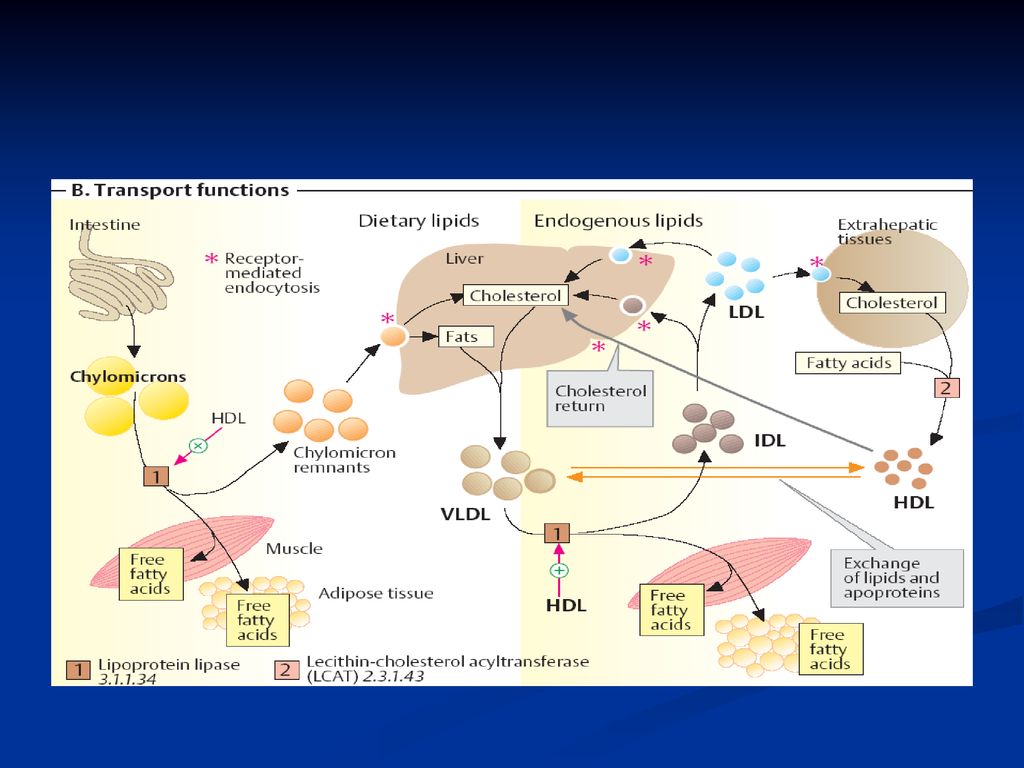
Metodele de separare LP plasmatice pot fi separate prin ultracentrifugare şi prin electroforeză. Prin centrifugare au fost obţinute 4 fracţiuni chilomicronii LP cu densitate foarte mică (VLDL, very low density lipoproteins) LP cu densitate mică (LDL, low density lipoproteins) LP cu densitate mare (HDL, high density lipoproteins)
 LP cu densitate mică (LDL, low density lipoproteins) LP cu densitate mare (HDL, high density lipoproteins)")
78
prin electroforeză: separarea se efectuează la pH alcalin (8,6) pe geluri de agaroză, de poliacrilamidă. Prin această tehnică sînt separate 4 fracţiuni: chilomicronii care nu migrează pre - - LP (corespund VLDL) - LP (corespund LDL) - LP (corespund HDL)
 - LP (corespund LDL) - LP (corespund HDL)")
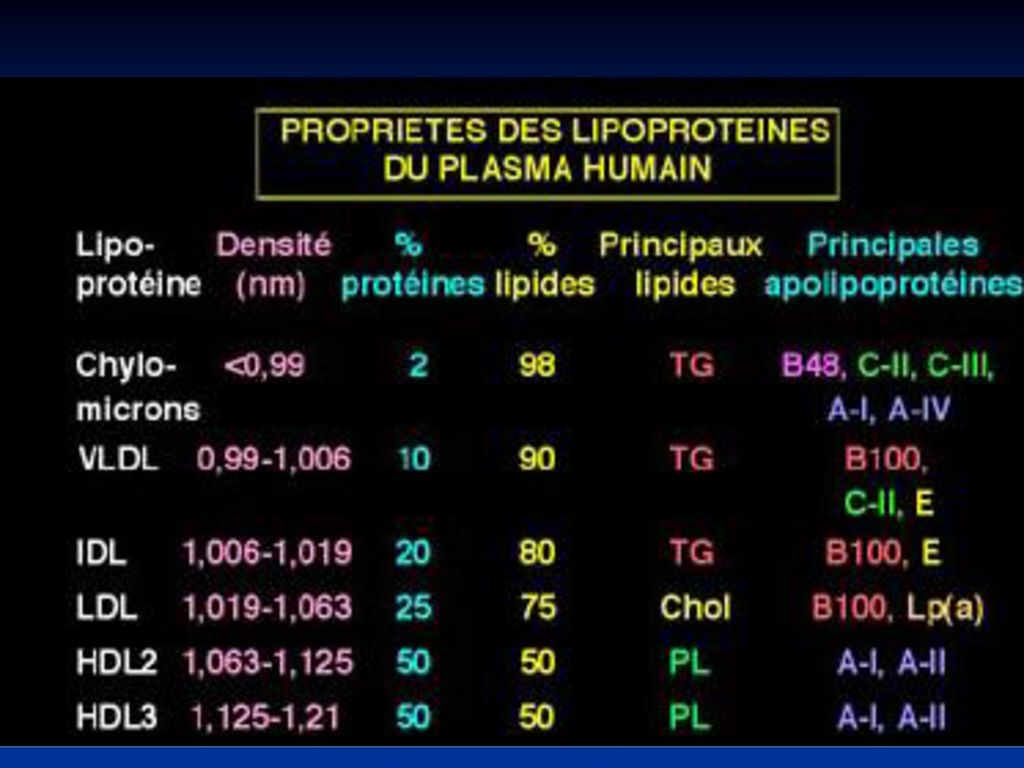
80
CM VLDL LDL HDL densitatea g/sm3 < 0,95 0,95-1,006 1,019- 1,063 1,12-1,21 Diametrul nm >100 25-75 22-24 6-12 % proteine 1-2 7-10 14-25 45-55 % lipide 98-99 90-93 75-86 lipidele Tg Col Fl Apo B48; C, E, A B100; C, E B100 AI; AII

81
Chilomicronii un conţinut mare de lipide (98-99% - Tg) şi puţine proteine (1-2%). Sunt sintetizaţi în celulele mucoasei intestinale şi încorporează lipidele alimentare absorbite (Tg). Sunt secretaţi în vasele limfatice care drenează intestinul şi la nivelul canalului toracic trec în plasmă. Particulile primare cuprind apo-B48 şi apoA. În plasmă CM primesc Apo C şi E de la -LP CM sînt prezenţi în plasmă după îngerare de alimente bogate în grăsimi. După 6-7 ore de la îngestia de grăsimi CM dispar din sînge.
. Sunt secretaţi în vasele limfatice care drenează intestinul şi la nivelul canalului toracic trec în plasmă. Particulile primare cuprind apo-B48 şi apoA. În plasmă CM primesc Apo C şi E de la -LP. CM sînt prezenţi în plasmă după îngerare de alimente bogate în grăsimi. După 6-7 ore de la îngestia de grăsimi CM dispar din sînge.")
82
Catabolismul chilomicronilor
are loc în două etape. În prima etapă sub acţiunea lipoproteinlipazei (LPL)Tg sînt hidrolizate în AG+glicerol. Ţesuturile utilizează AG pentru oxidare (muşchi scheletici, miocard), depozitare (ţesut adipos), secreţie de grăsimi (glanda mamară). pe cînd Col, FL, Apo C sunt transferate pe HDL. CM devin resturi CM A două etapă constă în captarea resturilor de CM de către ficat, facilitată de apo E (interacţionează cu E-receptorii din hepatocit). Pe această cale ajunge la ficat o parte din Col exogen şi cel intestinal.
Tg sînt hidrolizate în AG+glicerol. Ţesuturile utilizează AG pentru oxidare (muşchi scheletici, miocard), depozitare (ţesut adipos), secreţie de grăsimi (glanda mamară). pe cînd Col, FL, Apo C sunt transferate pe HDL. CM devin resturi CM. A două etapă constă în captarea resturilor de CM de către ficat, facilitată de apo E (interacţionează cu E-receptorii din hepatocit). Pe această cale ajunge la ficat o parte din Col exogen şi cel intestinal.")
84
Patologia Deficitul înăscut de LPL- hiperchilomicronemie
Mărirea concentraţiei CM, Tg, depunerea Tg în ţesuturi (xantoame) Anomalia sintezei de Apo B-48 – imposibilitatea formării CM şi transportului lipidelor exogene Tg se acumulează în celulele intestinale; este perturbată absorbţia lor; ele fiind eliminate prin masele fecale – steatoree În sînge: micşorarea lipidelor totale; Tg; Col
 Anomalia sintezei de Apo B-48 – imposibilitatea formării CM şi transportului lipidelor exogene. Tg se acumulează în celulele intestinale; este perturbată absorbţia lor; ele fiind eliminate prin masele fecale – steatoree. În sînge: micşorarea lipidelor totale; Tg; Col.")
85
Pre--LP (VLDL) sintetizate în ficat din lipidele endogene
au un conţinut ridicat de lipide (90-93%): 55-65% TG, 12-18% FL, Col esterificat 12-14% şi Col liber 6-8%. cuprind apo B-100 , în plasmă primesc apo C (apo CII) şi apo E de la -LP. Funcţia principală- transportul Tg sintetizate în ficat spre ţesuturile extrahepatice. VLDL sînt prezente în plasmă după îngerare de raţii bogate în glucide
: 55-65% TG, 12-18% FL, Col esterificat 12-14% şi Col liber 6-8%. cuprind apo B-100 , în plasmă primesc apo C (apo CII) şi apo E de la -LP. Funcţia principală- transportul Tg sintetizate în ficat spre ţesuturile extrahepatice. VLDL sînt prezente în plasmă după îngerare de raţii bogate în glucide.")
86
Catabolismul VLDL La nivelul ţesuturilor extrahepatice Tg din VLDL sub acţiunea LPL-azei se scindează în AG şi glicerol. Odată cu scăderea Tg are loc: pierderea de apo C-II care trece pe HDL Îmbogăţirea cu Col, furnizat de HDL - astefl VLDL sunt transformate în IDL. 50% sunt catabolizate în ficat prin interacţiunea cu E-R, iar 50% sunt transformate în LDL.

87
particulele se îmbogăţesc cu Col, prin 2 moduri:
Col esterificat este transferat din HDL pe VLDL sub acţiunea acil-Col-ester-transferazei (Apo D) Sub acţiunea lecitin-Col-acil-transferazei (LCAT) Apo C-I, component al VLDL, activează această E. LCAT catalizează reacţia: Lecitină Col lizolecitină + Acilcolesterol
 Sub acţiunea lecitin-Col-acil-transferazei (LCAT) Apo C-I, component al VLDL, activează această E. LCAT catalizează reacţia: Lecitină + Col lizolecitină + Acilcolesterol.")
88
Patologia Steatoza (infiltraţia grasă a ficatului) – acumularea grăsimilor în ficat Cauza: amplificarea sintezei Tg în ficat sau perturbarea căilor de sinteză şi transport (export) de VLDL Acţiune protectoare o au factorii lipotropi: Met; proteine bogate în Met; grăsimile nesaturate; vitamina E – ele favorizează exportul Tg hepatice
 de VLDL. Acţiune protectoare o au factorii lipotropi: Met; proteine bogate în Met; grăsimile nesaturate; vitamina E – ele favorizează exportul Tg hepatice.")
89
-LP se formează în plasmă din VLDL după îndepărtarea Tg (sub acţiunea LPL, TGL) şi îmbogăţire cu Col. au un conţinut lipidic de 75-86%, componenta majoră fiind Col: % - Col esterificat, 3-10% -Col liber; 20-25% - FL şi 8-12% - Tg. Apo majoritară este B-100. sunt prezente în sîngele recoltat dimineaţa după un post de 8-10 ore şi cuprinde 70% din Col total plasmatic. au rol de a furniza Col diverselor ţesuturi.

90
Catabolismul LDL În ficat, suprarenale, limfocite, celule musculare netede: LDL prin intermediul apo B-100 şi apo E interacţionează cu R fixate pe R, LDL sunt translocate în interiorul celulei şi fuzionează cu lizozomii În lizozomi LDL sub acţiunea E lizozomale sunt hidrolizate în: proteine, FL, Col esterificat, Tg. Col liber: este utilizat la construcţia membranelor, sinteza de hormoni steroidici, acizi biliari surplusul este esterificat şi depozitat în celulă. Esterificarea Col e catalizată de E - acil-CoA- Col-aciltransferaza (ACAT), rezultînd esteri ai Col cu acizii palmitic, palmitooleic, oleic.
, rezultînd esteri ai Col cu acizii palmitic, palmitooleic, oleic.")
91
Colesterolul liber: inhibă HMG-CoA-reductaza, ca rezultat sinteza Col în ţesuturile extrahepatice este menţinută la un nivel scăzut; inhibă sinteza R- B,E, astef determină micşorarea numărului lor pe membrană şi diminuarea captării LDL activează E microzomială ACAT

92
În macrofage: Macrofagele încorporează LDL care prezintă alterări ale componentelor proteice sau lipidice. Receptorii care recunosc LDL modificate nu sînt reglaţi prin feed-back negativ (cum se reglează receptorul clasic LDL) şi, ca urmare, prin încărcarea macrofagelor în mod excesiv cu Col apar “celulele spumoase”. Modificările biochimice ale LDL constau în acetilări, glicozilări ale apo (apo B-100, apo E) şi /sau alterări oxidative ale acizilor graşi nesatiuraţi.
 şi, ca urmare, prin încărcarea macrofagelor în mod excesiv cu Col apar celulele spumoase . Modificările biochimice ale LDL constau în acetilări, glicozilări ale apo (apo B-100, apo E) şi /sau alterări oxidative ale acizilor graşi nesatiuraţi.")
93
Patologia Hiperlipoproteinemia de tip II – deficienţă calitativă şi cantitativă a R membranari pentru LDL IIa – mărirea Col LDL – xantomatoză IIb – mărirea Col LDL+VLDL (Tg)- obezitate (lipsesc xantomele) Risc de aterogeneză înaltă
- obezitate (lipsesc xantomele) Risc de aterogeneză înaltă.")
94
-LP HDL conţinut lipidic de 45-55%, predominînd FL – 20-30%, Col 17-23% şi Tg 3-6%. Componenţa proteică - apo A (AI, AII) dar conţin cantităţi mici de apo C, D, E. Nu cuprind apo B. HDL sînt secretate şi sintetizate de hepatocite şi enterocite sub forma unor particule născînde de formă discoidală, alcătuite dintr-un strat dublu lipidic (FL; Col) şi apo A, apo E. Particulele născînde din intestin nu conţin apo E.
 dar conţin cantităţi mici de apo C, D, E. Nu cuprind apo B. HDL sînt secretate şi sintetizate de hepatocite şi enterocite sub forma unor particule născînde de formă discoidală, alcătuite dintr-un strat dublu lipidic (FL; Col) şi apo A, apo E. Particulele născînde din intestin nu conţin apo E.")
95
particulele născînde prin schimburi cu celelalte LP plasmatice se transformă în HDL mature.
Sub acţiunea LCAT - Col este esterificat şi migrează în interiorul particulei. Stratul superficial al HDL este ocupat de Col preluat din ţesuturi şi din alte LP plasmatice (CM).
.")
96
Catabolismul HDL HDL sînt catabolizaţi la nivelul ficatului:
Prin intermediul apo E particulele interacţionează cu R de pe suprafaţa hepatocitelor, sunt internalizaţi şi componentele degradate. Rolul principal al HDL este transportul Col din celulele extrahepatice în ficat, sediul catabolismului Col (transformare în AB şi excreţie prin bilă).
.")
97
Patologia Hipolipoproteinemia familiară – deficit de HDL
Cauza: deficit al sintezei de apoA Caracteristic: micşorarea HDL; Col HDL; FL Splenomegalie, hipotrofie amigdaliană; anomalii neurologice

99
Oxidarea glicerolului Sinteza corpilor cetonici
Oxidarea AG Oxidarea glicerolului Sinteza corpilor cetonici

100
Obiectivele Oxidarea acizilor graşi:
a) saturaţi cu număr par de atomi de carbon; b) nesaturaţi cu număr par de atomi de carbon; c) saturaţi cu număr impar de atomi de carbon; d) în peroxizomi. Reacţiile parţiale, enzimele, coenzimele, reglarea, randamentul energetic. Catabolismul triacilglicerolilor – reacţiile parţiale, enzimele, reglarea. Oxidarea glicerolului – reacţiile parţiale, enzimele, coenzimale, reglarea, randamentul energetic al oxidării anaerobe şi aerobe. Oxidarea fosfo-, sfingo- şi glicolipidelor. Metabolismul corpilor cetonici. Căile biosintezei şi utilizării lor – reacţiile parţiale, enzimele, coenzimele, reglarea. Rolul biologic al corpilor cetonici.
 saturaţi cu număr par de atomi de carbon; b) nesaturaţi cu număr par de atomi de carbon; c) saturaţi cu număr impar de atomi de carbon; d) în peroxizomi. Reacţiile parţiale, enzimele, coenzimele, reglarea, randamentul energetic. Catabolismul triacilglicerolilor – reacţiile parţiale, enzimele, reglarea. Oxidarea glicerolului – reacţiile parţiale, enzimele, coenzimale, reglarea, randamentul energetic al oxidării anaerobe şi aerobe. Oxidarea fosfo-, sfingo- şi glicolipidelor. Metabolismul corpilor cetonici. Căile biosintezei şi utilizării lor – reacţiile parţiale, enzimele, coenzimele, reglarea. Rolul biologic al corpilor cetonici.")
101
Oxidarea AG saturaţi cu număr par de atomi de carbon
β oxidarea AG (degradarea, scindarea, catabolizarea oxidativă a AG, spirala lui Lynen) – moleculele de AG suferă un atac oxidativ în poziţia β , urmat de desprinderea unui fragment cu 2C (Acetil Co A) Procesul e activ în muşchi, miocard, ficat, rinichi, testicul, splină, creier, plămîni şi ţesutul adipos.
 – moleculele de AG suferă un atac oxidativ în poziţia β , urmat de desprinderea unui fragment cu 2C (Acetil Co A) Procesul e activ în muşchi, miocard, ficat, rinichi, testicul, splină, creier, plămîni şi ţesutul adipos.")
102
3 etape: Activarea AG (citoplasmă) Transferul lui Acil CoA în mitocondrii b oxidarea propriu zisă (mitocondrii)
 .")
103
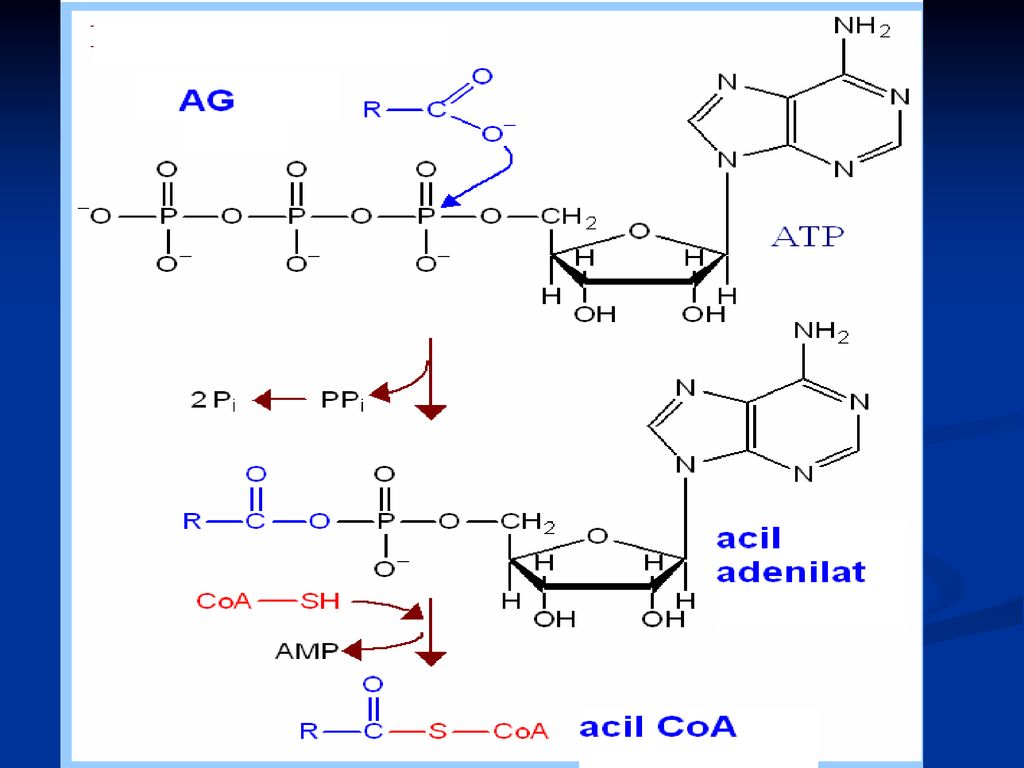
Activarea AG: Sumar: R-COOH + ATP R-COO-AMP + PPi aciladenilat
PPi 2 Pi R-COO-AMP + HS-CoA R-CO~SCoA + AMP acil-CoA Sumar: R-COOH + ATP + HS-CoA R-CO~SCoA + AMP + PPi R-COOH + ATP + HS-CoA +H2O R-CO~SCoA + AMP + 2 Pi E- acil Co A sintetaza Activatori: K; Mg Inhibitori: Na ; Li

105
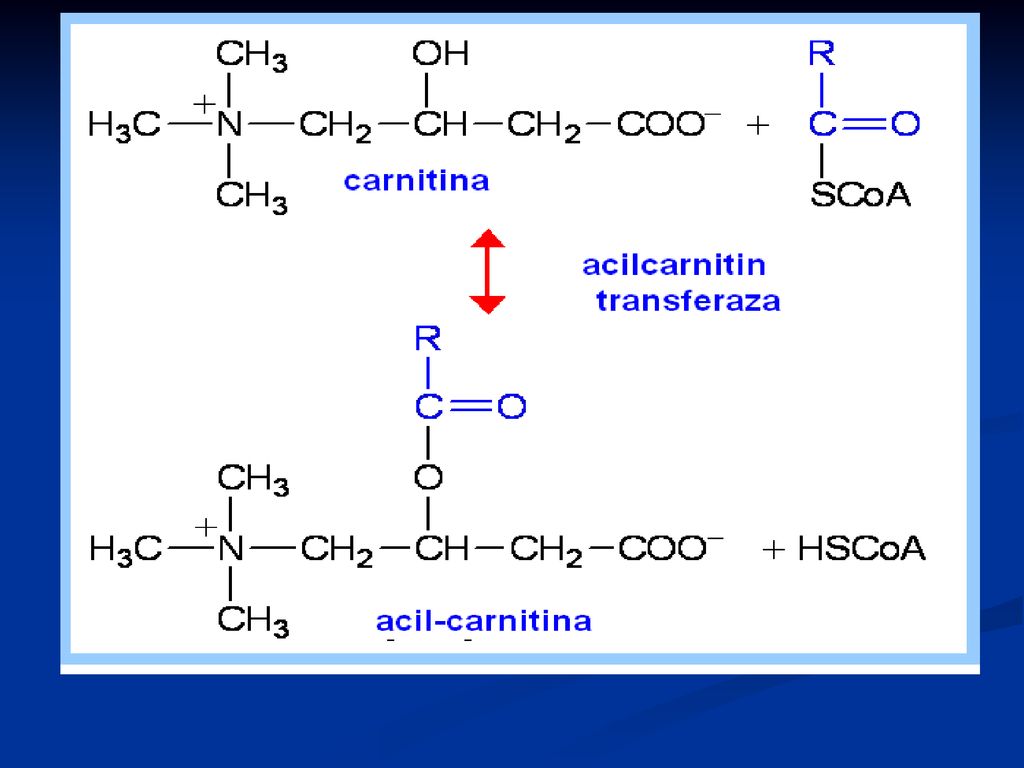
Transferul lui Acil CoA în mitocondrii
Acil CoA nu poate penetra membrana internă a MC Este transportat cu ajutorul carnitinei (β-hidroxi-γ-trimetilaminobutirat- vitamina Bt), ce se formează din Lyz şi Met activă cu participarea vitaminei C, B6, NAD
, ce se formează din Lyz şi Met activă cu participarea vitaminei C, B6, NAD.")
106
Transferul lui Acil CoA în mitocondrii

108
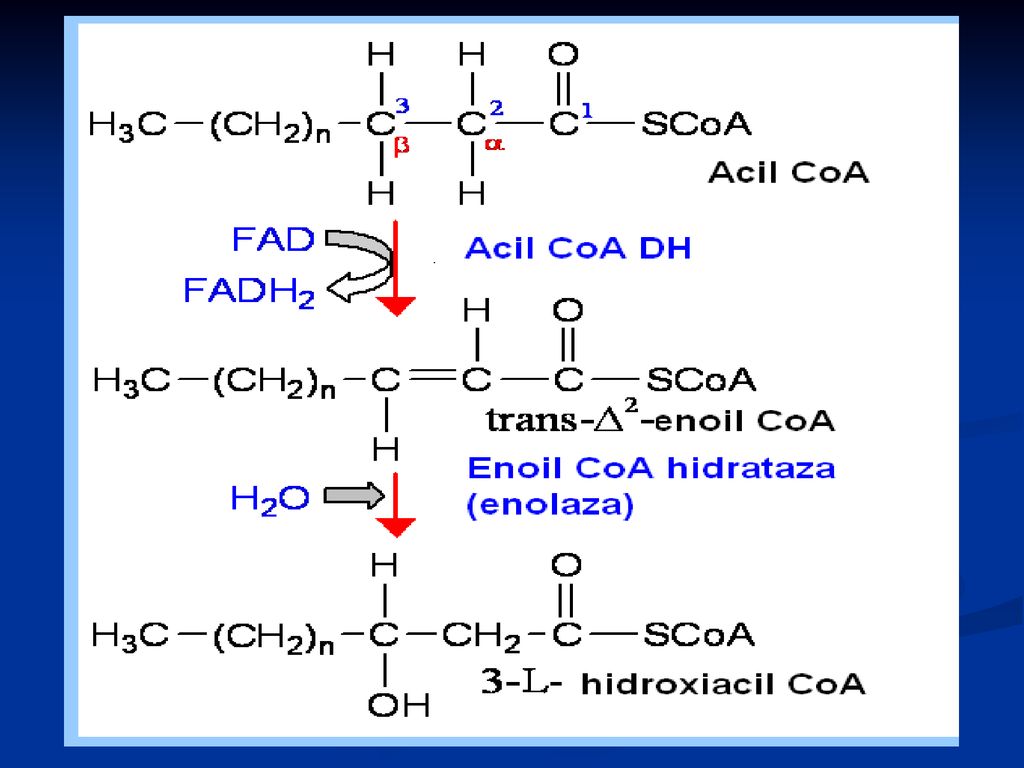
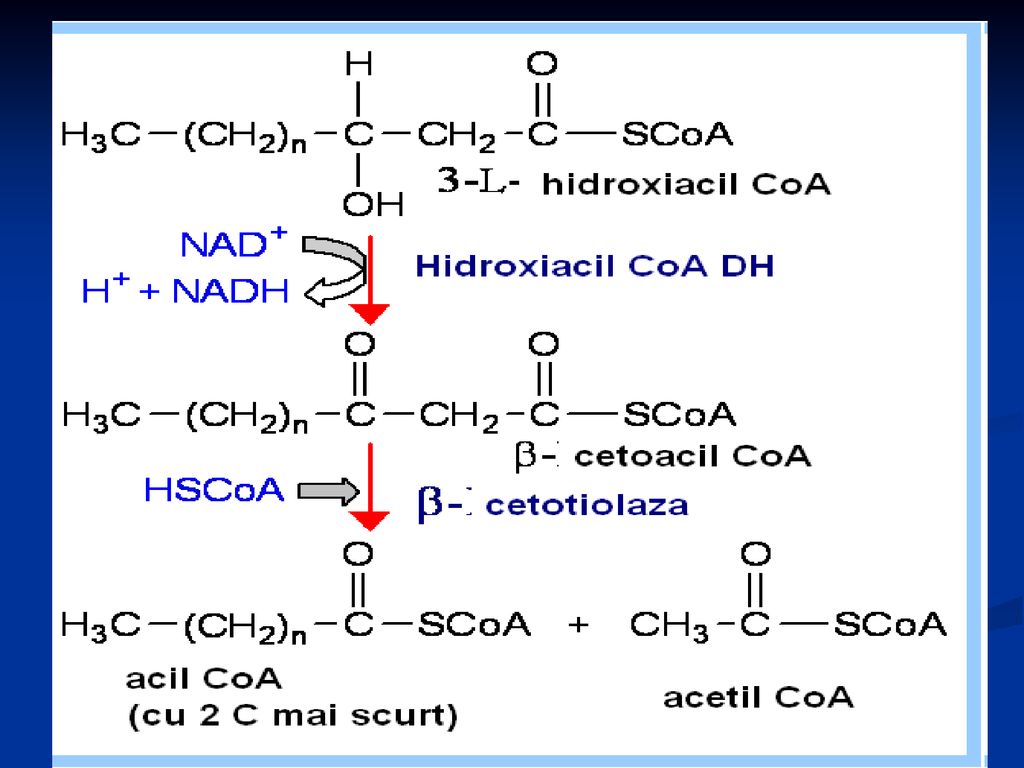
b oxidarea propriu zisă
Localizat în MC (matrixul MC) repetarea a 4 reacţii: Dehidrogenarea lui acil Co A (FAD) hidratare a doua dehidrogenare (NAD) reacţie tiolazică În rezultat - se formează acetil CoA şi acil CoA cu doi atomi de carbon mai puţin
 repetarea a 4 reacţii: Dehidrogenarea lui acil Co A (FAD) hidratare. a doua dehidrogenare (NAD) reacţie tiolazică. În rezultat - se formează acetil CoA şi acil CoA cu doi atomi de carbon mai puţin.")
111
Bilanţul energetic Stoichiometria unui ciclu de oxidare:
CH3- (CH2)n-CH2 – CH2-COSCoA +FAD+ H2O+NAD+HSCoA Acil CoA (Cn-2) +FADH2+NADH+H+ Acetil CoA Stoichiometria oxidării a. palmitic (C16): n/2 -1 – numărul de cicluri pînă la oxidarea completă n– numărul atomilor de C
n-CH2 – CH2-COSCoA +FAD+ H2O+NAD+HSCoA. Acil CoA (Cn-2) +FADH2+NADH+H+ Acetil CoA. Stoichiometria oxidării a. palmitic (C16): n/2 -1 – numărul de cicluri pînă la oxidarea completă. n– numărul atomilor de C.")
112
Stoichiometria oxidării a. palmitic
16/2 -1 = 7 cicluri 7FADH – 7 X 2=14ATP 7NADH+H X3=21 ATP 8 CH3COSCoA--- 8X12= 96 ATP Sumar: 131 mol de ATP Deoarece 2 legături macroergice sunt irosite la activarea acidului – beneficiul net este de 129

113
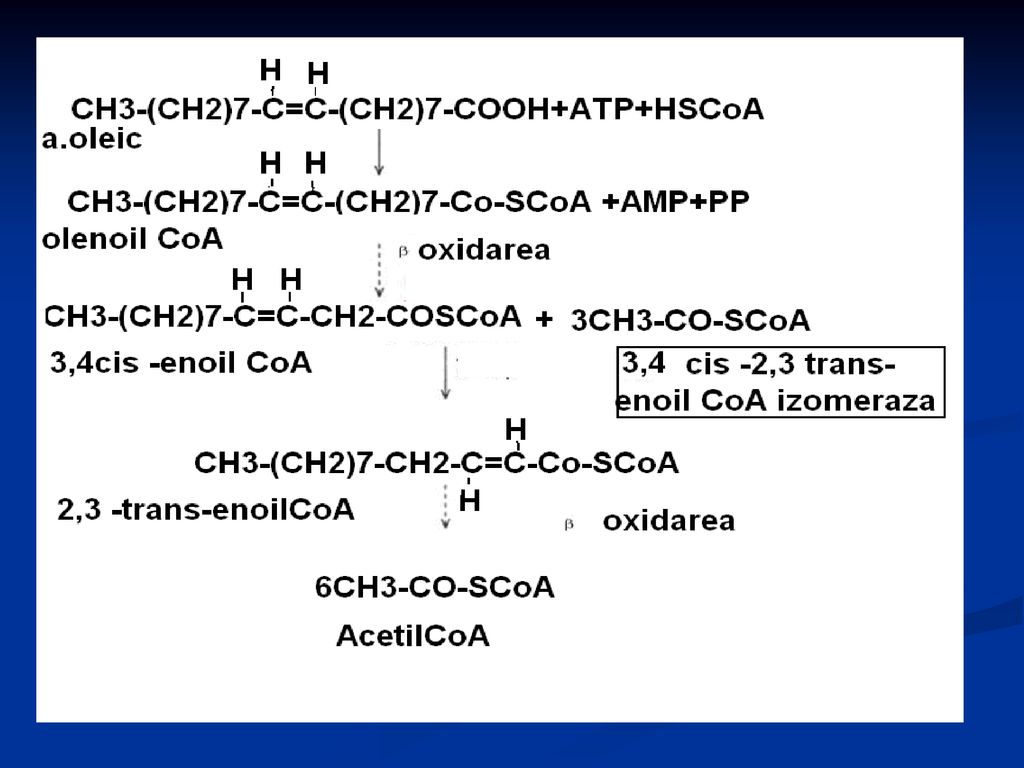
Oxidarea AG nesaturaţi
-oxidarea AG nesaturaţi se desfăşoară normal pînă în vecinătatea legăturii duble (cis configuraţie) După trei cicluri normale de -oxidare se ajunge la un cis – Δ3 – enoil – CoA. Sub acţiunea izomerazei legătura dublă din cis – Δ3 trece în trans-∆ 2 – se formează trans – Δ2 – enoil – CoA, intermediar normal al -oxidării. Exemplu: oxidarea acidului oleic (C18:1Δ9) CH3-(CH2)7-CH=CH-(CH2)7-COOH
 După trei cicluri normale de -oxidare se ajunge la un cis – Δ3 – enoil – CoA. Sub acţiunea izomerazei legătura dublă din cis – Δ3 trece în trans-∆ 2 – se formează trans – Δ2 – enoil – CoA, intermediar normal al -oxidării. Exemplu: oxidarea acidului oleic (C18:1Δ9) CH3-(CH2)7-CH=CH-(CH2)7-COOH.")
115
Pentru AG polienici e necesară şi o altă enzimă – epimeraza, care modifică configuraţia grupei OH la C3. Această E e rezultată din hidratarea legăturii duble D-izomer-3 hidroxiacil CoA, ce nu poate fi substrat al enzimei de tipul L

116
Oxidarea AG cu număr impar de atomi de C
Se oxidează în acelaşi mod ca AG saturaţi, dar în ultima etapă se formează o moleculă de propionil CoA (C3) şi una de Acetil CoA.
 şi una de Acetil CoA.")
117
Oxidarea AG cu număr impar de atomi de C
E- propionil CoA carboxilaza Co- vitamina H (biotin dependentă) E- Metilmalonil-mutaza Co- vitamina B12 Lipsa acestei E – acidemie metilmalonică (în sînge şi urină apare acidul metilmalonic, micşorînd pH sîngelui (administrat vitamina B12)
 E- Metilmalonil-mutaza. Co- vitamina B12. Lipsa acestei E – acidemie metilmalonică (în sînge şi urină apare acidul metilmalonic, micşorînd pH sîngelui (administrat vitamina B12)")
118
Oxidarea AG în peroxisomi
Caracteristic AG C20-C26 Produsul final este Acetil CoA, dar nu este asociată cu sinteza de ATP (acetil CoA trece în MC unde este oxidat la CO2 şi H2O) Diferă de oxidarea mitocondrială prin reacţia de oxidare a acil-CoA la enoil-CoA (E- oxidază) R-(CH2)n-COSCoA+O2▬► R-(CH2)-CH=CH-COSCoA + H2O2 ( sub acţiunea catalazei 2H2O2 ▬►2H2O+O2) Amploarea acestui proces variază în dependenţă de factorii nutriţionali, hormonali, medicamentoşi. Numărul peroxisomilor creşte în diabet, inaniţie, la administrarea unor medicamente (aspirina, preparate hipolemiante) Absenţa peroxisomilor- sindromul Zellweger: creşterea AG cu catena lungă şi deces în primele luni de viaţă
 Diferă de oxidarea mitocondrială prin reacţia de oxidare a acil-CoA la enoil-CoA (E- oxidază) R-(CH2)n-COSCoA+O2▬► R-(CH2)-CH=CH-COSCoA + H2O2 ( sub acţiunea catalazei 2H2O2 ▬►2H2O+O2) Amploarea acestui proces variază în dependenţă de factorii nutriţionali, hormonali, medicamentoşi. Numărul peroxisomilor creşte în diabet, inaniţie, la administrarea unor medicamente (aspirina, preparate hipolemiante) Absenţa peroxisomilor- sindromul Zellweger: creşterea AG cu catena lungă şi deces în primele luni de viaţă.")
119
Oxidarea α Predomină în ţesutul nervos (creier)
Se formează hidroxiacizii graşi superiori, proprii lipidelor SNC Necesită: NAD, Vitamina C, ATP, O2, Fe2+ Nu intervine CoA şi nu se formează ATP E- acid gras peroxidaza (necesită H2O2, ce rezultă prin autooxidarea flavinenzimelor) Au loc concomitent 2 procese: eliminarea carboxilului sub formă de CO2 oxidarea lui C α la aldehidă Aldehida poate fi redusă la alcool sau oxidată la acidul corespunzător Nu are loc degradarea totală a AG, deoarece E este activă numai la AG C13-C18.
 Au loc concomitent 2 procese: eliminarea carboxilului sub formă de CO2. oxidarea lui C α la aldehidă. Aldehida poate fi redusă la alcool sau oxidată la acidul corespunzător. Nu are loc degradarea totală a AG, deoarece E este activă numai la AG C13-C18.")
120
Oxidarea ω Are loc în microsomi Necesită: O2, NADPH, citocromul P450
E – monooxigenaza hepatică Generează acizi dicarboxilici ce se degradează în final prin β oxidare

121
Metabolismul TG În plasmă există 2 fluxuri de TG:
CM – transportă TG exogene – de la intestin la ţesuturi VLDL – transportă TG endogene - de la ficat spre ţesuturi Mobilizarea TG din ţesutul adipos are loc în etape, pînă la glicerol şi AG, sub acţiunea lipazelor (mono-; di- , triacilglicerollipaza).
.")
122
Soarta AG şi glicerolului:
AG sunt transportaţi spre ţesuturi de albumina serică, unde: se supun oxidării - acetil-CoA (pentru a obţine ATP sau poate fi utilizat la sinteza Col, corpilor cetonici). se activează şi participă la sinteza TG, depozitate în ţesutul adipos difuzează în plasmă şi circulă sub formă de AG liberi (sunt captaţi de ţesuturile periferice: muşchii scheletici, miocard, rinichi, ficat) Eritrocitele şi creierul nu pot utiliza AG ca sursă de energie Glicerolul: Sinteza de TG şi FL Sinteza glucozei - gluconeogeneză Oxidează pînă la CO2 şi H2O
. se activează şi participă la sinteza TG, depozitate în ţesutul adipos. difuzează în plasmă şi circulă sub formă de AG liberi (sunt captaţi de ţesuturile periferice: muşchii scheletici, miocard, rinichi, ficat) Eritrocitele şi creierul nu pot utiliza AG ca sursă de energie. Glicerolul: Sinteza de TG şi FL. Sinteza glucozei - gluconeogeneză. Oxidează pînă la CO2 şi H2O.")
123
Oxidarea glicerolului
E1 glicerolkinaza E2 glicerolfosfatDH

124
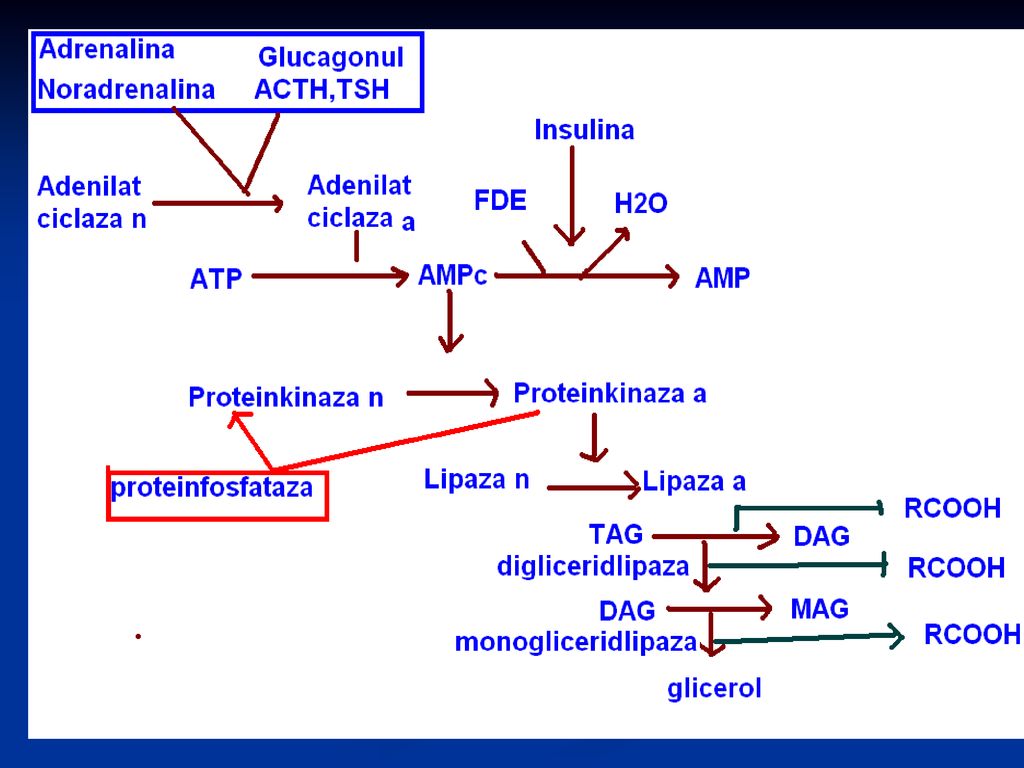
Trigliceridlipaza Enzima cheie a lipolizei - trigliceridlipaza adipocitară, cunoscută ca “lipaza hormonsensibilă”. Enzima este convertibilă prin fosforilare –defosforilare. Forma fosforilată este activă. Catecolaminele (adrenalina, noradrenalina) sînt factori majori lipolitici. Glucagonul are acelaş efect. Insulina, prostoglandina E sînt factori antilipolitici, ei favorizează sinteza de TG în ţesutul adipos.
 sînt factori majori lipolitici. Glucagonul are acelaş efect. Insulina, prostoglandina E sînt factori antilipolitici, ei favorizează sinteza de TG în ţesutul adipos.")
126
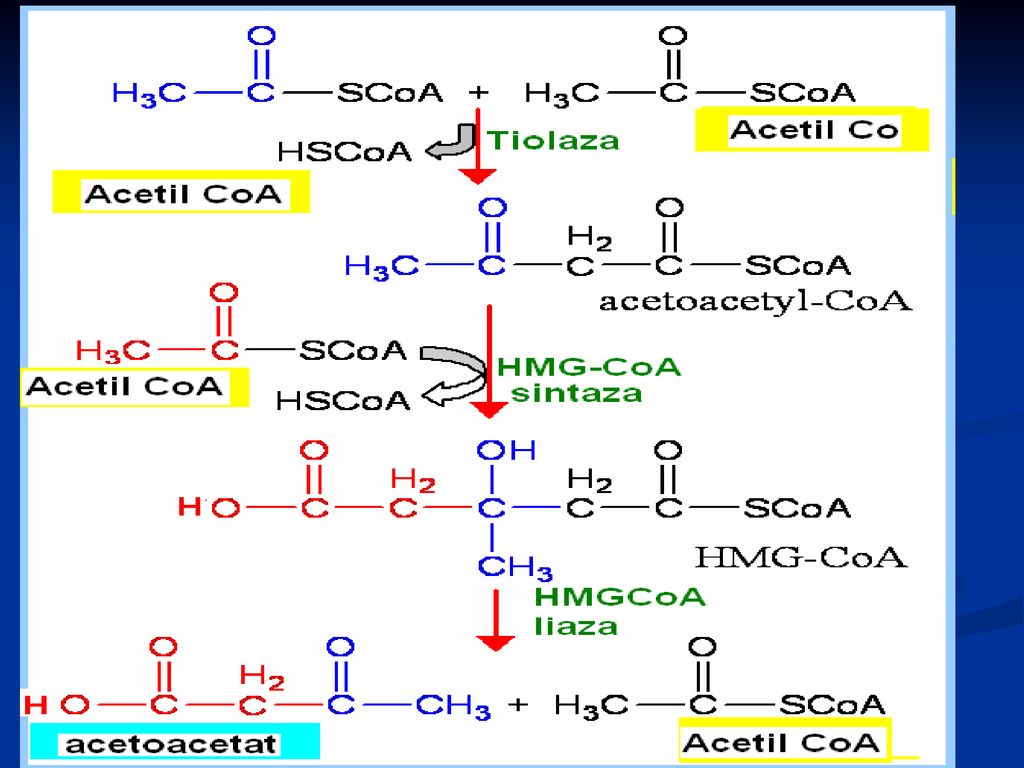
Sinteza corpilor cetonici (cetogeneza)
Principala cale de metabolizare a acetil CoA – includerea în ciclul Krebs (în condiţiile în care scindarea lipidelor şi a glucidelor este echilibrată) - “lipidele ard în flacăra glucidelor” În lipsa glucidelor; inaniţie, diabet - OA se utilizează pentru generarea Gl. În lipsa OA, Acetil Co A recurge la formarea corpilor cetonici: acetoacetatul, β-hidroxibutiratul şi acetona Sinteza lor are loc în ficat, dar se utilizează de ţesuturile periferice Au rol energetic (muşchiul cardiac, stratul cortical al rinichilor)
 - lipidele ard în flacăra glucidelor În lipsa glucidelor; inaniţie, diabet - OA se utilizează pentru generarea Gl. În lipsa OA, Acetil Co A recurge la formarea corpilor cetonici: acetoacetatul, β-hidroxibutiratul şi acetona. Sinteza lor are loc în ficat, dar se utilizează de ţesuturile periferice. Au rol energetic (muşchiul cardiac, stratul cortical al rinichilor)")
128
cetogeneza

129
Utilizarea corpilor cetonici
Calea tiokinazică: - Acetoacetatul – 2 mol de acetil CoA, utilizate ulterior în ciclul Krebs (23 ATP)
")
130
A doua cale de activare a acetoacetatului:
Calea tioforazică: Reacţie de schimb cu succinil CoA

131
Acetona: pînă la propandiol (CH3-CHOH-CH2OH) , scindat la fragmente acetil şi formil Transformată în piruvat (prin hidroxilare dublă)
 .")
132
Cetonemie, cetonurie Cetonemie- mărirea c% de corpi cetonici în sînge
Cetonurie – apariţia CC în urină Diete bogate în lipide, sărace în glucide; inaniţie, diabet, dereglări gastrointestinale la copii sau gravide; glucozurie renală Eliminarea hidroxibutiratului şi acetoacetatului din organism (fiind anioni la excreţie) conduce la pierderea de cationi – Na - rezultă cetoacidoza Pierderea H2O – deshidratarea organismului
 conduce la pierderea de cationi – Na - rezultă cetoacidoza. Pierderea H2O – deshidratarea organismului.")
133
Biosinteza lipidelor

134
Obiectivele: Biosintaza acizilor graşi:
saturaţi cu număr par de atomi de carbon; nesaturaţi cu număr par de atomi de carbon; saturaţi cu număr impar de atomi de carbon. Enzimele, coenzimele, reglarea. Biosinteza TAG: substanţele iniţiale, enzimele şi coenzimele, reglarea. Biosinteza fosfogliceridelor: substratele, reacţiile parţiale ale I şi a II căi; Biosinteza sfingolipidelor: precursorii, reacţiile principale, enzimele, reglarea. Metabolismul colesterolului. Biosinteza colesterolului – substratele, etapele, reacţiile parţiale ale I etape (până la acidul mevalonic), enzimele, coenzimele, reglarea. Căile de utilizare şi eliminare ale colesterolului.
, enzimele, coenzimele, reglarea. Căile de utilizare şi eliminare ale colesterolului.")
135
Sinteza AG Sinteza AG şi încorporarea lor în Tg constituie mecanismul principal de stocare a excesului de glucide alimentare (Gl nu se mai transformă în glicogen dar în Tg) Etapele: Sinteza de novo cu formarea acidului palmitic Elongarea acidului palmitic Introducerea de legături duble în AG
 Etapele: Sinteza de novo cu formarea acidului palmitic. Elongarea acidului palmitic. Introducerea de legături duble în AG.")
136
Particularităţile sintezei AG
Are loc în citozol E – acid gras sintetaza – alcătuită din 8 proteine (domenii)- 7 sunt enzime, a 8-a – proteina (purtătoare) transportatoare de acil -ACP. ACP cuprinde 2 grupe SH: –SH furnizat de un rest de cisteinil: SH-Cis - SH - fosfopantoteina, ataşată prin legătura fosfat-Ser: SH-Pant Ca iniţiator este acetil CoA (rezultat din glicoliză), pe când sursa majoră – malonil CoA rolul reducător îi revine NADPH+H
- 7 sunt enzime, a 8-a – proteina (purtătoare) transportatoare de acil -ACP. ACP cuprinde 2 grupe SH: –SH furnizat de un rest de cisteinil: SH-Cis. - SH - fosfopantoteina, ataşată prin legătura fosfat-Ser: SH-Pant. Ca iniţiator este acetil CoA (rezultat din glicoliză), pe când sursa majoră – malonil CoA. rolul reducător îi revine NADPH+H.")
137
Sinteza de novo cu formarea acidului palmitic
Etapele: transferul lui Acetil CoA din mitocondrii în citozol Sinteza de malonil CoA Sinteza acidului palmitic

138
Transferul lui Acetil CoA din mitocondrii în citozol

139
Sinteza de malonil CoA acetil-CoA + HCO3- + ATP ADP + Pi + malonil-CoA E- acetil CoA carboxilaza A-citrat, insulina I- palmitoil CoA glucagonul

140
Sinteza acidului palmitic

141
Sinteza acidului palmitic

142
Sinteza acidului palmitic
Ciclu de reacţii este reluat: butiril+ACP se condensează cu malonil+ACP- formînd în final C6-acil ACP. Catena AG creşte pînă la formarea palmitil-S-ACP

143
deoarece malonil CoA se sintetizează din acetil CoA:
Reacţia sumară: Acetil-ACP+7 malonil-CoA +14 NADPH+H Palmitat +7CO2+14NADP + + 8HSCoA+6H2O deoarece malonil CoA se sintetizează din acetil CoA: 8 acetil-CoA + 14 NADPH +H ATP palmitat+ 14 NADP+ + 8HSCoA + 7 ADP + 7 Pi +6H2O

144
Elongarea AG Localizată: reticulul endoplasmatic AG este activat
La acidul preexistent (palmitil CoA) se ataşează malonil CoA
 se ataşează malonil CoA.")
145
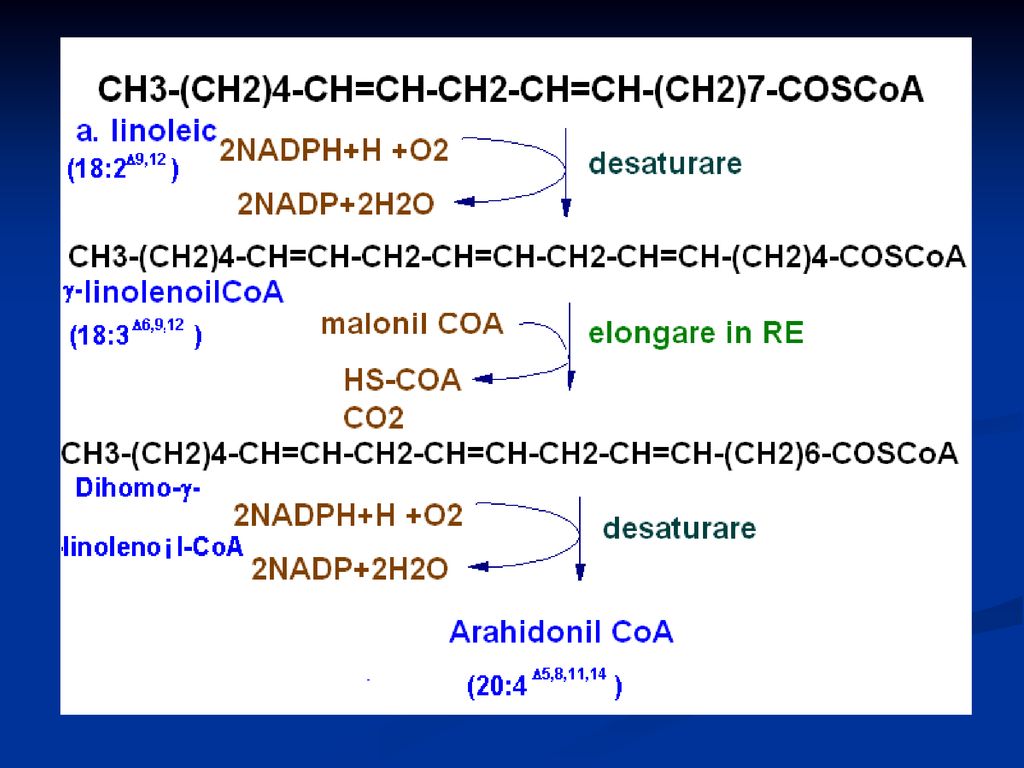
Biosinteza AG nesaturaţi
Pot fi sintetizaţi AC mononesaturaţi. Introducerea unei duble legături are loc prin acţiunea unei monooxigenaze (introduce gruparea hidroxil), urmată de deshidratare Acidul linoleic şi linolenic sunt esenţiali (exogen) Acidul linoleic se transformă în acidul arahidonic conform reacţiilor
, urmată de deshidratare. Acidul linoleic şi linolenic sunt esenţiali (exogen) Acidul linoleic se transformă în acidul arahidonic conform reacţiilor.")
147
Sinteza TAG 2 căi: calea monoacilglicerolului: are loc în peretele intestinal (enterocite) din produşi absorbiţi (resinteza lipidelor). calea glicerolfosfatului: în toate ţesuturile (activă: ţesutul adipos şi ficat)
")
148
AG sunt incorporaţi în TAG sub formă activă de acilCoA:
R-COOH + ATP + HS-CoA +H2O R- CO~SCoA + AMP + 2 Pi E- acil Co A sintetaza

149
1. calea monoacilglicerolului
TG împreună cu FL, Col, proteine sunt incorparate în CM şi secretaţi mai departe în vasele limfatice.

150
calea glicerolfosfatului

151
originea glicerol fosfatului
În ficat, rinichi: În ţesut adipos, ficat

152
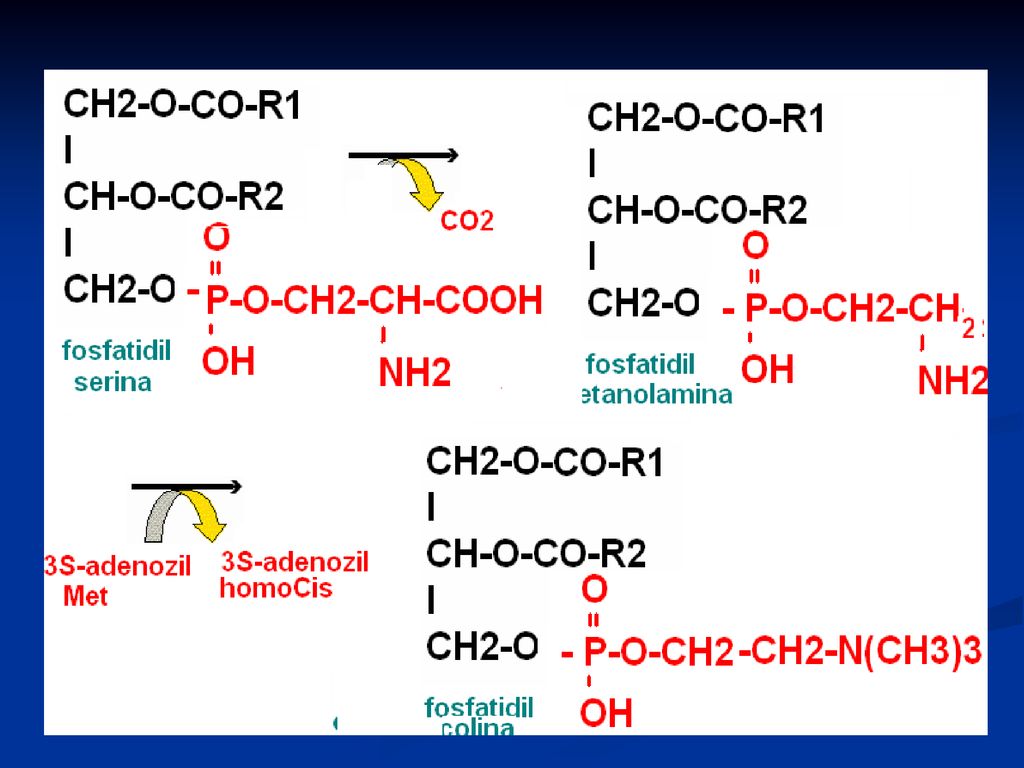
Sinteza glicerofosfolipidelor
2 căî de sinteză: Sinteza de novo - utilizează ca intermediar comun acidul fosfatidic Calea de rezervă – o sinteză din produse formate Particularitatea biosintezei FL este participarea precursorilor în forme active de derivaţi ai citidin fosfatului (CDP) ca CDP-colina, CDP-etanolamina, CDP-diglicerid.
 ca CDP-colina, CDP-etanolamina, CDP-diglicerid.")
153
Sinteza de novo

155
2. sinteza din produse formate

156
Sinteza sfingolipidelor
Se formează din palmitoil CoA şi Ser Sfingozina liberă se formează din ceramidă Sinteza are loc pe suprafaţa citozolică a membranelor reticulului endoplasmatic

158
Sinteza sfingolipidelor

159
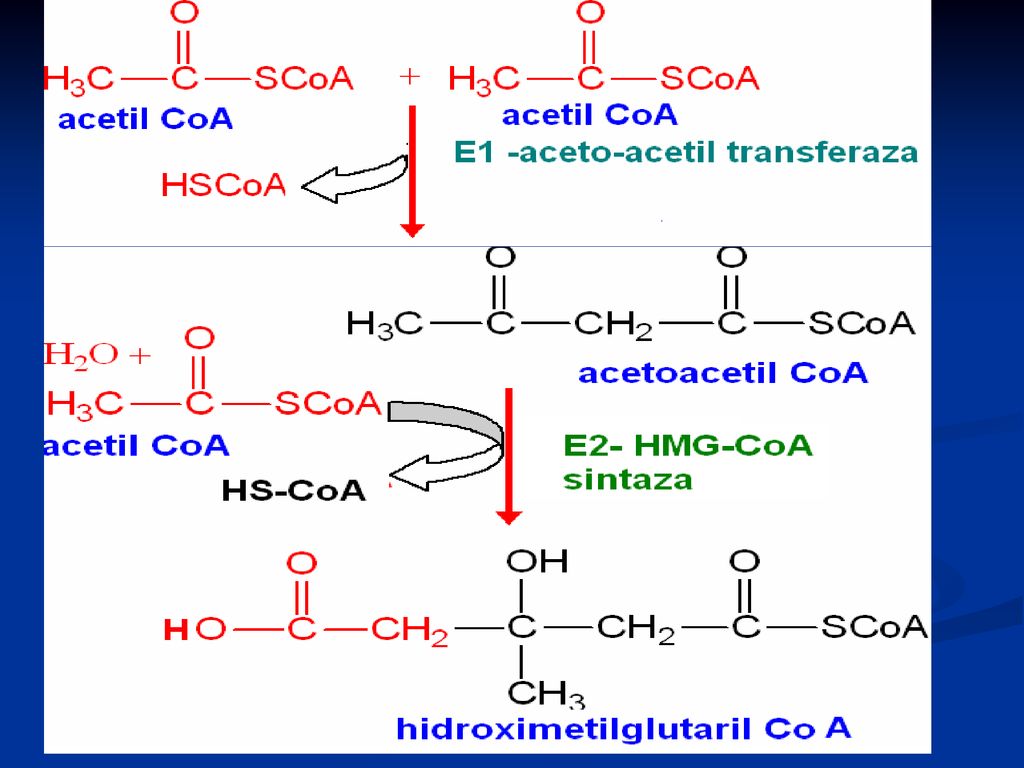
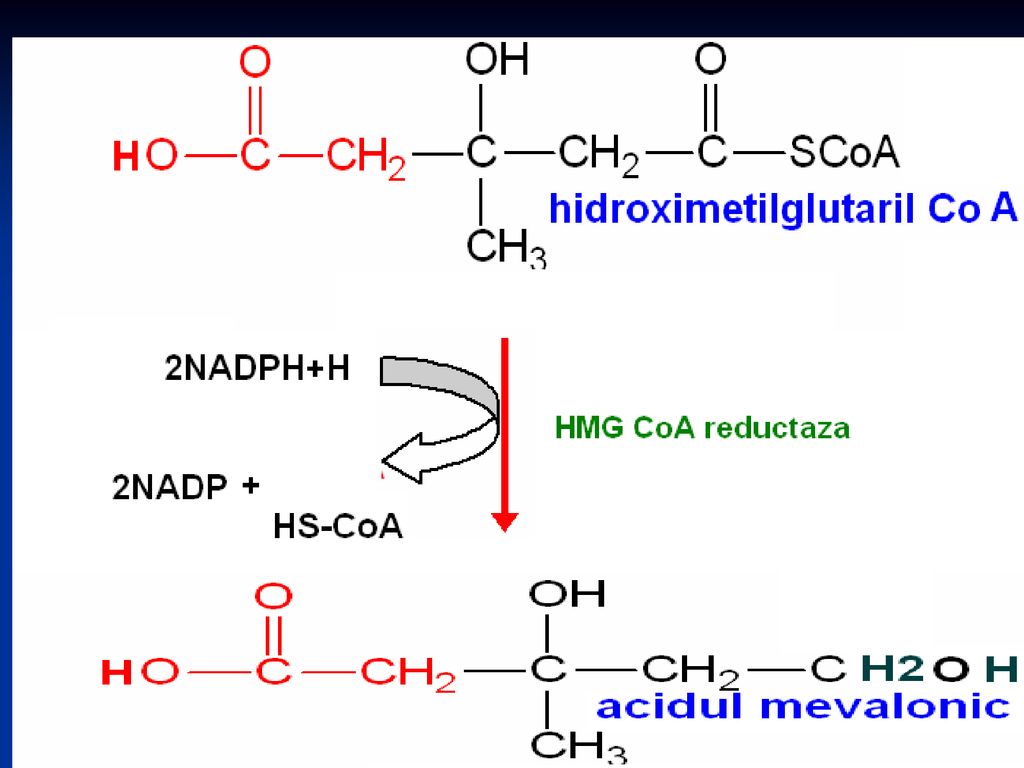
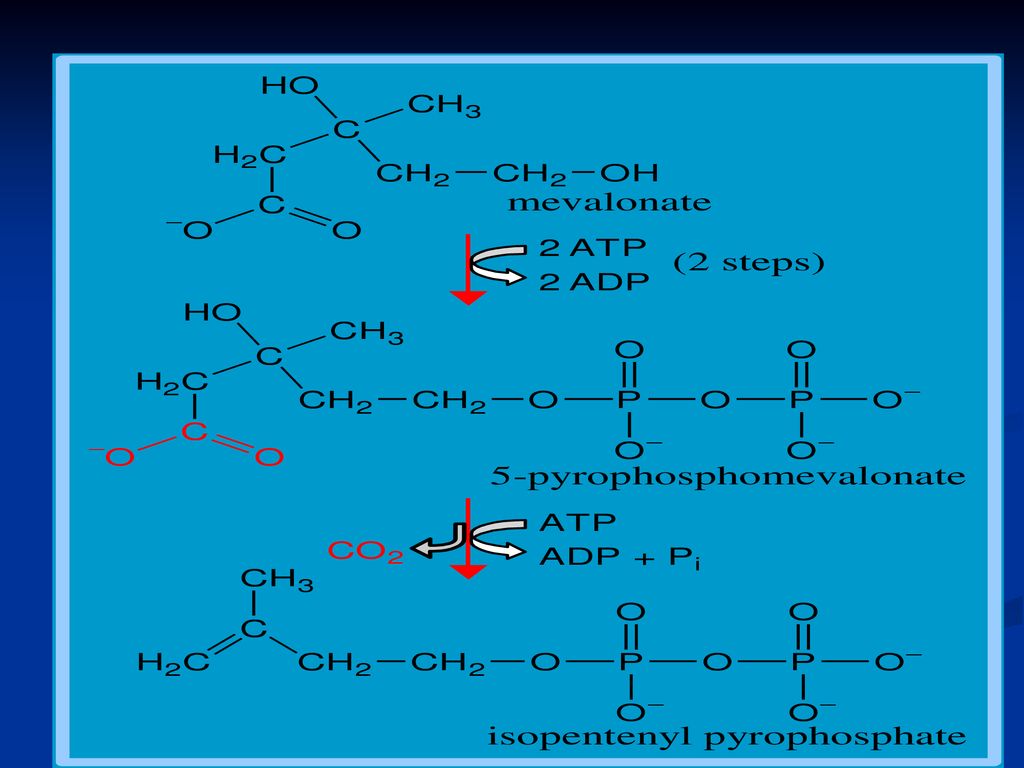
Sinteza Colesterolului
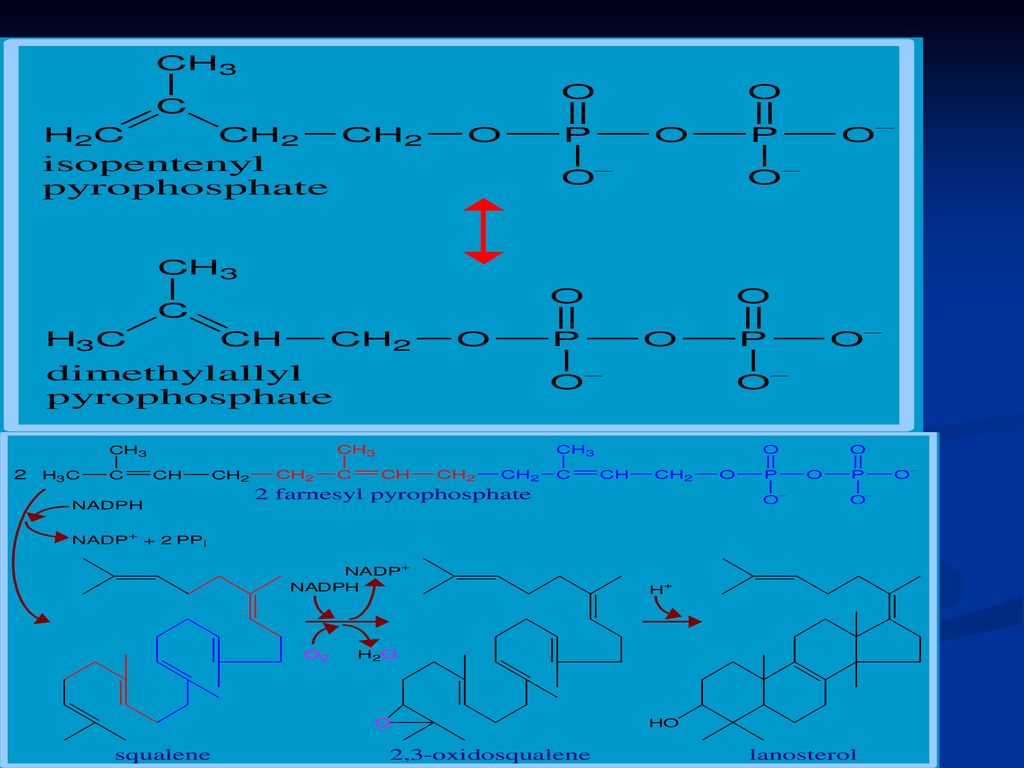
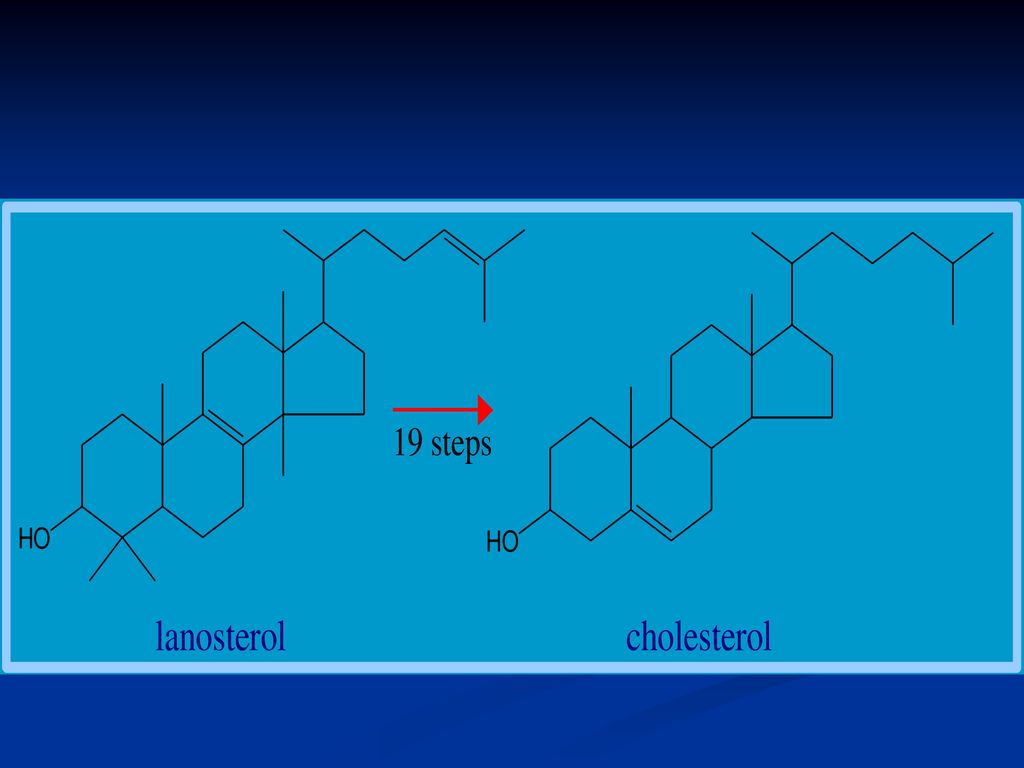
Se sintetizează din Acetil-CoA Necesită 18 moli de Acetil-CoA, 16 NADPH+H şi 18 de ATP Principalul organ de metabolizare este ficatul, dar are loc şi în intestin, suprarenale, tegumente Are loc în 3 etape: Sinteza acidului mevalonic mevalonatul prin mai multe reacţii - 3∆-izopentenil pirofosfat. 6 molecule de 3∆-izopentenil pirofosfat – scualen Scualenul se supune ciclizării – lanosterol -- Col

162
Reglarea Reacţia – limită a biosintezei e formarea acidului mevalonic, catalizată de o enzimă compusă reglatoare – hidroximetil-glutaril-CoA reductaza HMG-CoA reductaza – localizată la nivelul RE Structura: 887 AA cu 2 domenii structurale: unul hidrofob (membranar) şi altul citoplasmatic (posedă activitate catalitică).
 şi altul citoplasmatic (posedă activitate catalitică).")
163
Reglarea biosintezei colesterolului
la nivelul sintezei (v de transcriere a genelor) la nivelul modificării v de degradare prin cantitatea Col celular (represor al sintezei E dar creşte v de degradare) forma activă este forma defosforilată (insulina) Enzima este inhibată de mevalonat şi de colesterol
 la nivelul modificării v de degradare. prin cantitatea Col celular (represor al sintezei E dar creşte v de degradare) forma activă este forma defosforilată (insulina) Enzima este inhibată de mevalonat şi de colesterol.")
164
Conţinutul Col depinde de regimul alimentar
Conţinutul Col depinde de regimul alimentar. Acest colesterol alimentar frânează sinteza reductazei în ficat insulina şi hormonii tiroidieni activeză HMG-CoA reductaza, iar glucagonul la animale are efect antagonist Reglarea e dependentă şi de c % proteinei specifice ce transportă Col

168
REGLAREA ŞI PATOLOGIA METABOLISMULUI LIPIDIC

169
Obiectivele Metabolismul eicosanoizilor. Căile ciclooxigenazică şi lipooxigenazică ale biosintezei lor. Inactivarea. Metabolismul vitaminelor liposolubile: sursele alimentare, necesităţile diurne, transformările Reglarea metabolismului lipidelor la nivelul celulei. Reglarea neurohormonală a metabolismului lipidelor. Rolul lipotropinelor, ACTH, hormonilor tiroizi, insulinei, glucagonului, glucocorticoizilor şi catecolaminelor. Relaţiile reciproce dintre metabolismul energetic, glucidic şi lipidic. Dereglările digestiei şi absorbţiei lipidelor. Steatoreea pancreatică, hepatică şi intestinală. Dislipidemiile: a) hipolipoproteinemiile familiale – afecţiunea Tangier, - şi -lipoproteinemia familială; b) hiperlipoproteinemiile primare şi familiale; c) hiperlipoproteinemiile secundare (dobândite) – în diabet zaharat, alcoolism, afecţiuni ale glandelor endocrine. Cauze, mecanismele dereglării metabolismului lipidelor, manifestările biochimice. 6. Lipidozele tiszlare: a) ereditare – Neimann-Pick, Tay-Sachs, Krabbe, Gaucher, Farber, leucodistrofia metacromatică, gangliozidoza GM1; b) dobândite – obezitate, ateroscleroză, alcoolism. 7. A-, hipo- şi hipervitaminozele A, D, E, K – cauze, manifestări metabolice. 8. Rolul eicosanoizilor în procesele inflamatorii, reacţiile alergice, dereglările fluidităţii sanguine.
 hipolipoproteinemiile familiale – afecţiunea Tangier, - şi -lipoproteinemia familială; b) hiperlipoproteinemiile primare şi familiale; c) hiperlipoproteinemiile secundare (dobândite) – în diabet zaharat, alcoolism, afecţiuni ale glandelor endocrine. Cauze, mecanismele dereglării metabolismului lipidelor, manifestările biochimice. 6. Lipidozele tiszlare: a) ereditare – Neimann-Pick, Tay-Sachs, Krabbe, Gaucher, Farber, leucodistrofia metacromatică, gangliozidoza GM1; b) dobândite – obezitate, ateroscleroză, alcoolism. 7. A-, hipo- şi hipervitaminozele A, D, E, K – cauze, manifestări metabolice. 8. Rolul eicosanoizilor în procesele inflamatorii, reacţiile alergice, dereglările fluidităţii sanguine.")
170
Reglarea metabolismului lipidic
Se efectuiază în mai multe aspecte: Factorii Enzimatici: a. Modificarea activităţii E incluse în sinteza lipidelor b.Modificarea V sintezei şi degradării E Factorii de mediu: stresul emoţional negativ, inaniţia, hipotermia, efortul muscular cauzează pierderea din greutate a organismului.

171
Factorii hormonali: Catecolaminele stimulează lipoliza prin mesagerul secund, activează lipaza hormono-dependentă.Glucagonul are acţiune asemănătoare. La fel şi hormonul de creştere, dar cu o fază mai tardivă (stimulează sinteza adenilat ciclazei de novo). Steroizii, tiroxina au efecte benefice – insuficienţa lor favorizează depozitarea de grăsime. Insulina posedă efect contrar, activează fosfodiesteraza – inhibă lipoliza şi stimulează lipogeneza.
. Steroizii, tiroxina au efecte benefice – insuficienţa lor favorizează depozitarea de grăsime. Insulina posedă efect contrar, activează fosfodiesteraza – inhibă lipoliza şi stimulează lipogeneza.")
172
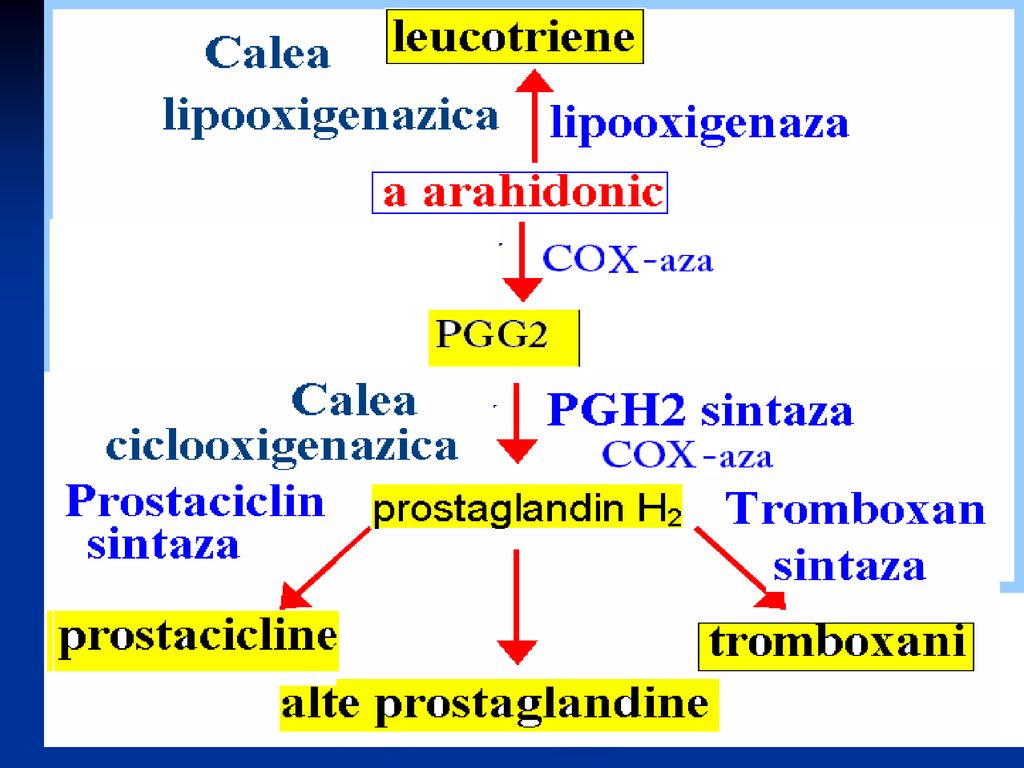
Eicosanoizii Sunt substanţe biologic active de natură lipidică, derivate din AG polinesaturaţi (a.arahidonic sau prostanoic) şi îndeplinesc în organism roluri multiple. Deosebim: Prostaglandinele (clasice- PG şi endoperoxizii prostaglandinici (PGG2 şi PGH2) Prostaciclina (PGI2) Tromboxanii (TX) Leucotrienele (LT)
 şi îndeplinesc în organism roluri multiple. Deosebim: Prostaglandinele (clasice- PG şi endoperoxizii prostaglandinici (PGG2 şi PGH2) Prostaciclina (PGI2) Tromboxanii (TX) Leucotrienele (LT)")
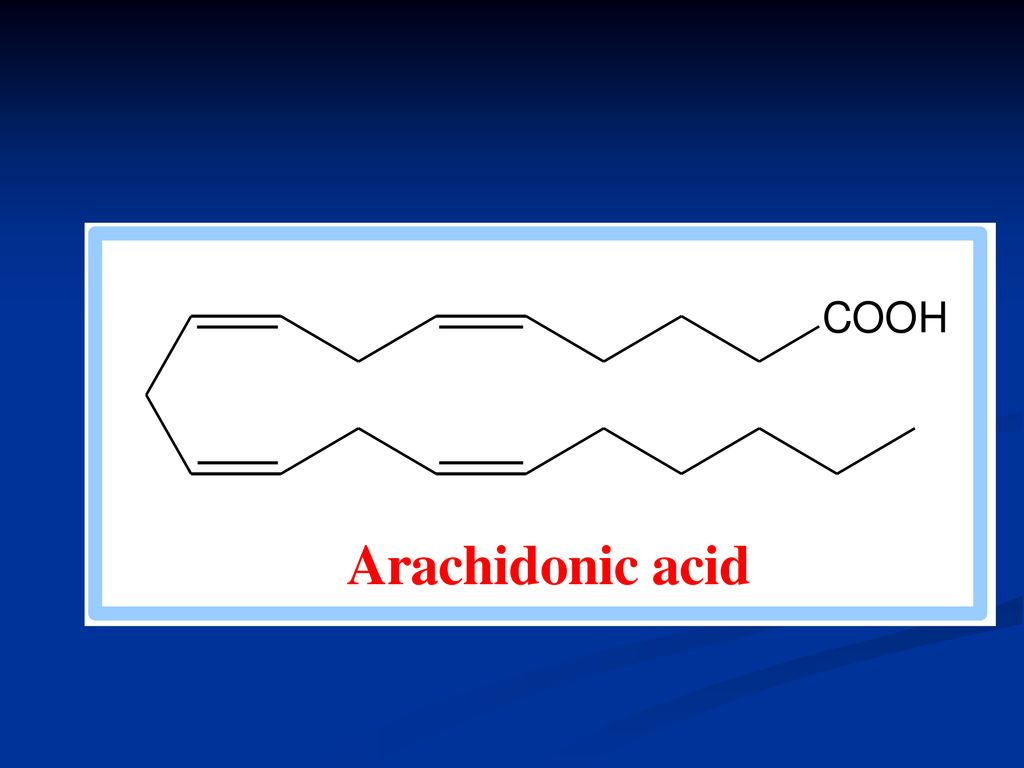
173
Structura chimică Reprezintă AG nesaturaţi cu 20 C
Sunt derivaţi ai acidului prostanoic Existenţa unui ciclu pentanic format între C8-C12 PG se deosebesc între ele prin: 1. natura substituienţilor din nucleul pentanic (PGA-PGI) 2. Catenele laterale pot conţine un număr diferit de legături duble (PG1, PG2, PG3)
 2. Catenele laterale pot conţine un număr diferit de legături duble (PG1, PG2, PG3)")
175
PGE2

176
Biosinteza eicosanoizilor
Are loc la nivelul tuturor ţesuturilor După sinteză nu se depozitează dar acţionează imediat Are loc în 2 etape: Sinteza acidului arahidonic (comună) Sinteza PG, TH, LT
 Sinteza PG, TH, LT.")
177
Sinteza acidului arahidonic
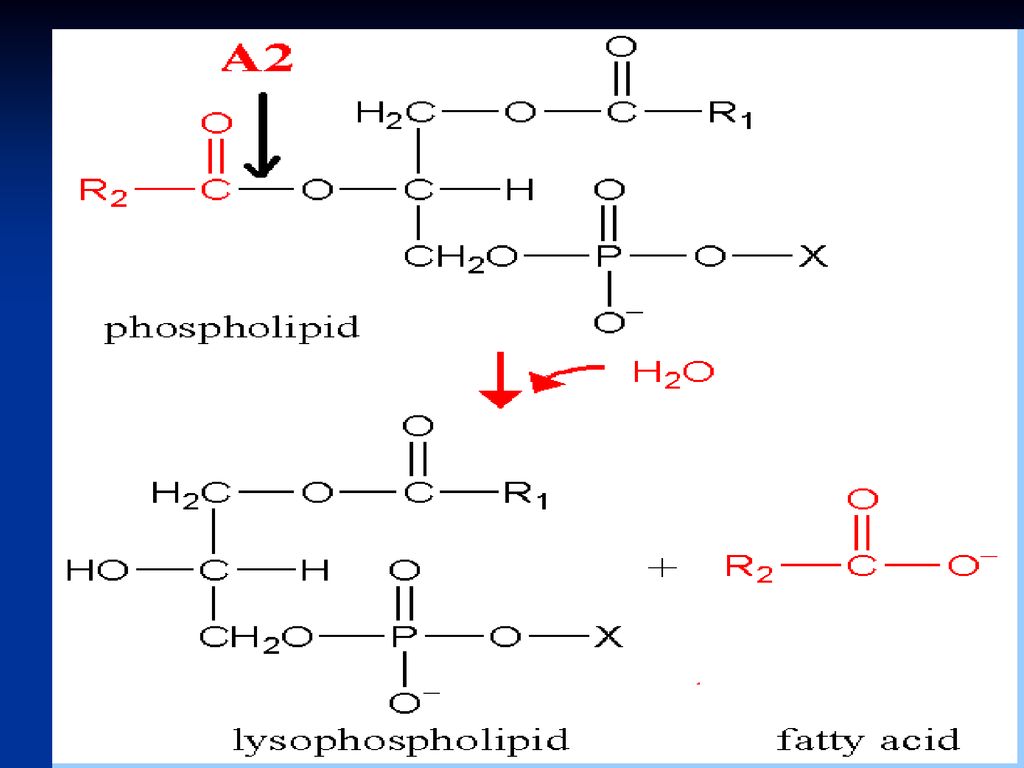
Derivat al acidului linoleic Este prezent la nivelul membranelor celulare Se eliberează prin activarea fosfolipazei A2

179
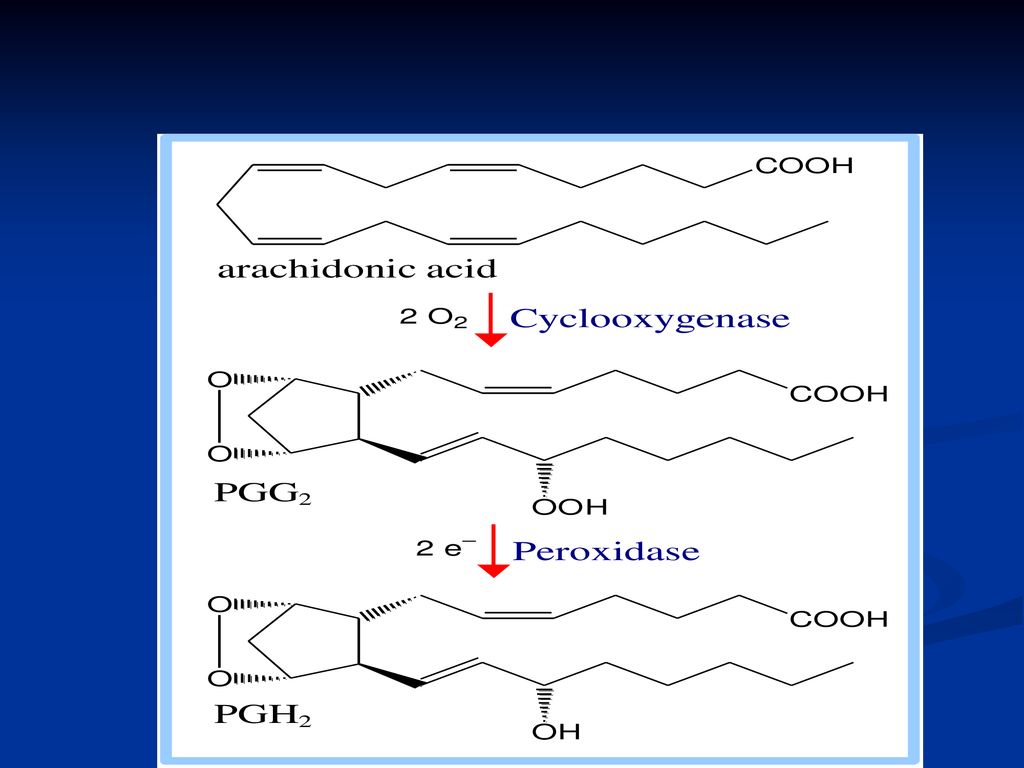
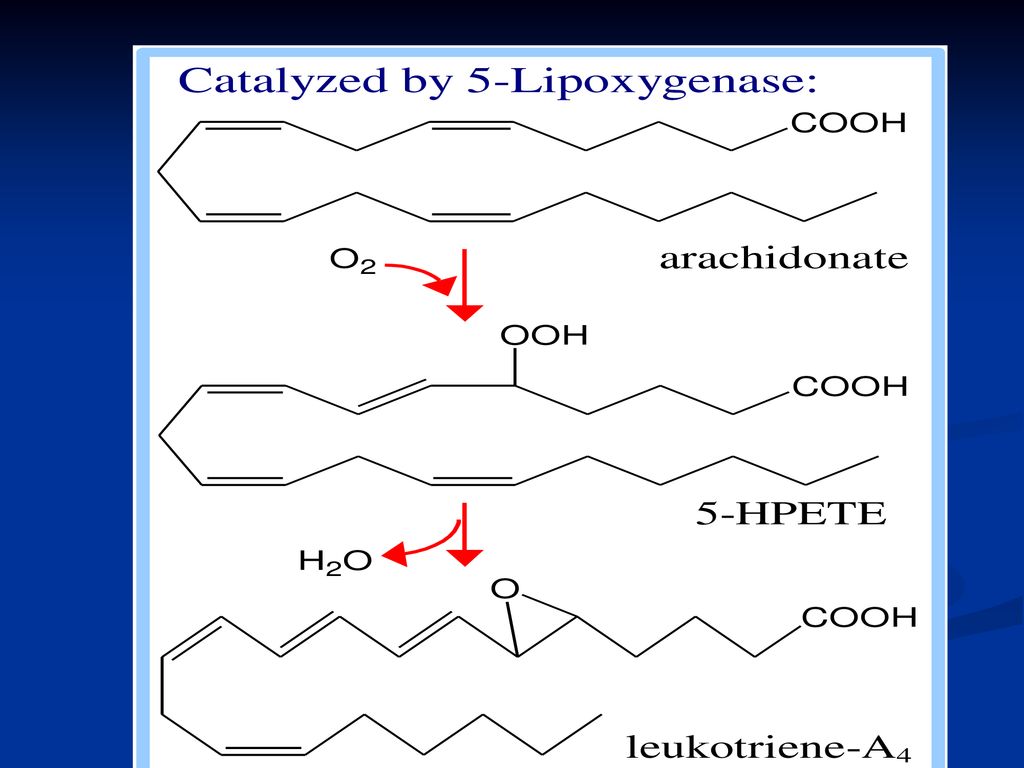
Sinteza PG, TX şi LT 2 căi: Calea ciclooxigenazei conduce la sinteza PG, TX Calea lipooxigenazei – la sinteza LT

181
Ciclooxigenaza este o hemoproteină, cu o activitate dublă:
a. dioxigenazică (încorporează O2 în substrat) b. peroxidazică (descompune peroxidul).
 b. peroxidazică (descompune peroxidul).")
183
TX conţin în structura lor ciclul piranic (TXA2 şi TXB2)
TXA2 este sintetizat de PGH2 de către sintaza tromboxanică microzomală.

184
Structura chimică LT (LTA-LTE) – cuprind în structura lor mai multe legături duble, trei fiind conjugate. Cele mai active la om sunt LTA4.

185
LTA4

186
Inhibitorii sintezei:
Corticosteroizii – inhibă fosfolipaza A2 Aspirina (medicamente antiinflamatoare nesteroide) - inhibă ireversibil COX (prin acetilarea grupelor hidroxil a resturilor de Ser localizate le situsul activ al enzimelor). Derivaţii imidazolului – inhibă tromboxan sintaza.
 - inhibă ireversibil COX (prin acetilarea grupelor hidroxil a resturilor de Ser localizate le situsul activ al enzimelor). Derivaţii imidazolului – inhibă tromboxan sintaza.")
187
Catabolismul PG Viaţă biologică scurtă:
TXA2 are T1/2= 30 secunde, şi suferă o hidroliză rapidă, nonenzimatică la inactivul TXB2. Prostaciclina (PGI2) - T ½ - 3 minute la 37 °C şi pH 7,5 şi este convertită printr-o hidroliză nonenzimatică la forma inactivă. PG clasice sunt eliminate la nivelul plămînului Principalii derivaţi sunt 15-ceto- iar ai prostaciclinelor 6 ceto-, care se elimină cu urina.
 - T ½ - 3 minute la 37 °C şi pH 7,5 şi este convertită printr-o hidroliză nonenzimatică la forma inactivă. PG clasice sunt eliminate la nivelul plămînului. Principalii derivaţi sunt 15-ceto- iar ai prostaciclinelor 6 ceto-, care se elimină cu urina.")
188
Proprietăţile biologice ale prostanoidelor
O serie de efecte ale prostanoidelor sunt mediate prin intermedizul adenilat-ciclazelor sau prin mobilizarea Ca2+ intracelular. PG creşte AMPc în adenohipofiză, corpus luteum, osul fetal, plachetele pulmonare şi tiroidă, însă descreşte în ţesutul adipos. Tromboxanii blochează producerea de AMPc de către PG şi mobilizează Ca2+ intracelular.

189
Acţiunile biologice Diferă nu numai de natura chimică (PG, LT, TX) dar şi de cea a tipului celular La nivelul TGI: PGF2α – acţiune contractilă asupra musculaturii netede longitudinală şi circulară; PGE2 – relaxează musculatura circulară, acţiune inhibitorie asupra secreţiei de suc gastric şi absorbţiei intestinale.

190
Acţiunile biologice Sistemul respirator: PGE şi PGF – fiind implicate în stări patologice ca edem pulmonar, embolie, astm. PGF2α – acţiune contractilă asupra traheei, bronhiilor PGE2- bronhodilatatoare SNC: reglarea transmisiei sinaptice. PGE2- scade c% de GMPc – efecte tranchilizante, anticonvulsive PGF2α – creşte c% GMPc – efecte contrare

191
Acţiunile biologice Sistemul de reproducere: la bărbaţi influenţează spermatogeneza; la femei controlează ciclul ovarian. PGF2α - acţiune contractilă asupra musculaturii netede a uterului. Sistem circulator: Prostaciclina şi TX – controlează tonusul vaselor sanguine şi agregarea plachetară. TXA2 – acţiune contractilă – creşte TA. La nivelul plachetelor – stimulează agregarea şi formarea de tromb Prostaciclina – acţiuni opuse (relaxează musculatura vaselor; efect antitrombic)
")
192
Acţiunile biologice LT (LTB4) – acţionează ca agenţi chemotactici şi chemocinetici, determinând acumularea de neutrofile în focarul inflamator. La nivelul plămânului – efecte contractile asupra musculaturii netede. LTC4 - responsabilă de producerea şocului anafilactic.
 – acţionează ca agenţi chemotactici şi chemocinetici, determinând acumularea de neutrofile în focarul inflamator. La nivelul plămânului – efecte contractile asupra musculaturii netede. LTC4 - responsabilă de producerea şocului anafilactic.")
194
Vitaminele sunt substanţe biologic active necesare pentru creşterea, dezvoltarea şi supravieţuirea organismului. nu se sintetizează în organismul uman (prezenţa lor în alimente este obligatorie). nu servesc în calitate de sursă energetică, dar îndeplinesc funcţii coenzimatice.
. nu servesc în calitate de sursă energetică, dar îndeplinesc funcţii coenzimatice.")
195
avitaminoze şi hipovitaminoze
avitaminoze - lipsa vitaminelor în alimente hipovitaminoze - insuficienţa vitaminelor în alimente Ambele apar şi se dezvoltă în stări patologice specifice Cauzele apariţiei lor: pot fi exogene (atunci când vitamina respectivă lipseşte sau se conţine în produsele alimentare insuficient) endogene (dereglări în absorbţie, transport, antivitamine etc.)
 endogene (dereglări în absorbţie, transport, antivitamine etc.)")
196
Provitamine - predecesorii vitaminelor, care în organism se transformă în vitaminele respective (de ex. în morcovi, caise se conţine -carotena, care în ficat se va transformă în vitamina A).
.")
197
Antivitaminele – substanţe (inclusiv şi unele medicamente) ce se aseamănă mult ca structură cu unele vitamine. La majorarea c% lor - în mod competitiv ele se vor uni cu E în locul vitaminelor --- se formează complexul apoenzimă-antivitamină lipsit de activitate --- se dereglează reacţiile metabolice catalizate de aceste enzime.

198
Preparatele sulfanilamidice prezintă antivitamine pentru microorganisme deoarece ele mult se aseamănă după structură cu acidul p-aminobenzoic, factorul de creştere a bacteriilor. La majorarea concentraţiei în organism a acestor medicamente, ele se vor include în sistemele enzimatice ale bacteriilor şi bacteriile vor pierde capacitatea de a se înmulţi (efect bacteriostatic).
.")
199
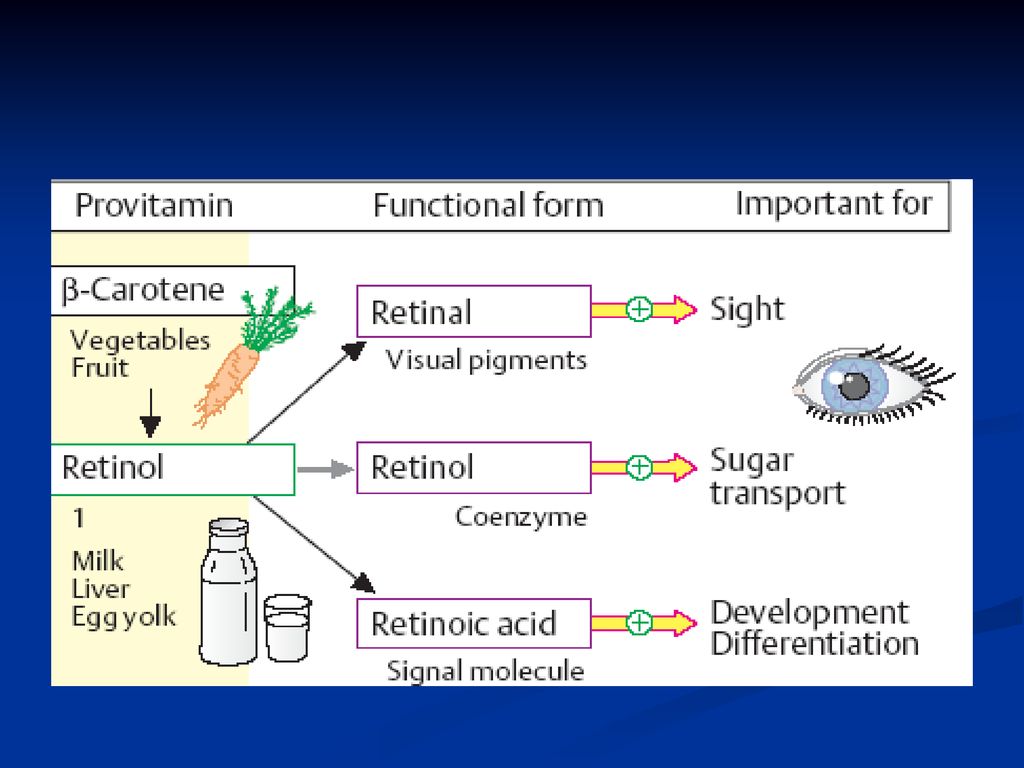
Vitamina A vitamina creşterii, retinol, antixeroftalmică
La baza structurii acestei vitamine se află ciclul -ionon legat cu 2 resturi de izoprenă

201
Metabolism Digestia are loc în intestin
Absorbţia – în regiunea superioară a intestinului Retinolul absorbit uneşte AG saturaţi şi este încorporat în CM – sânge - ficat

202
Rolul vitaminei A Retinol – acţiune hormonală decesivă pentru funcţia de reproducere Retinal – component al pigmentului vizual – rodopsina (din bastonaşele retinei) A. retinoic – participă la sinteza glicoproteinelor
 A. retinoic – participă la sinteza glicoproteinelor.")
204
Rodopsina este constituită dintr-o proteină simplă – opsina -11-cis-retinal
Absorbţia cuantelor de lumină de către rodopsină conduce la descompunerea rodopsinei în opsină şi retinal cu structura trans El va trece în 11-cis retinal prin absorbţie de fotoni care refac înapoi rodopsina (E- retinal-izomeraza)
")
205
Alte efecte: La oxidarea AG
Biosinteza gliceridelor, fosfolipidelor, Col, h. steroizi Creşte asimilarea purinelor şi sinteza AN Sunt reglatori ai permeabilităţii membranelor Afectează expresia genetică implicată atât în proliferarea cât şi diferenţierea celulelor normale sau maligne

206
Vitamina A Se conţine în produse de origine animală (unt, ficat etc.). Unele produse de origine vegetală (morcov, caise etc) conţin carotina (provitamina), care în ficat sub influenţa anumitor enzime este convertită în vitamina A.
. Unele produse de origine vegetală (morcov, caise etc) conţin carotina (provitamina), care în ficat sub influenţa anumitor enzime este convertită în vitamina A.")
207
Carenţa Vitaminei A se caracterizează prin:
La nivelul ochilor: xeroftalmia (uscarea corneei); în cazuri grave keratomalaţia (degradarea globului ocular); hemeralopia (orbul găinei). La nivelul pielei şi mucoaselor: hiperkeratoza, descuamaţie, ulcerări, inflamaţii.
; în cazuri grave keratomalaţia (degradarea globului ocular); hemeralopia (orbul găinei). La nivelul pielei şi mucoaselor: hiperkeratoza, descuamaţie, ulcerări, inflamaţii.")
208
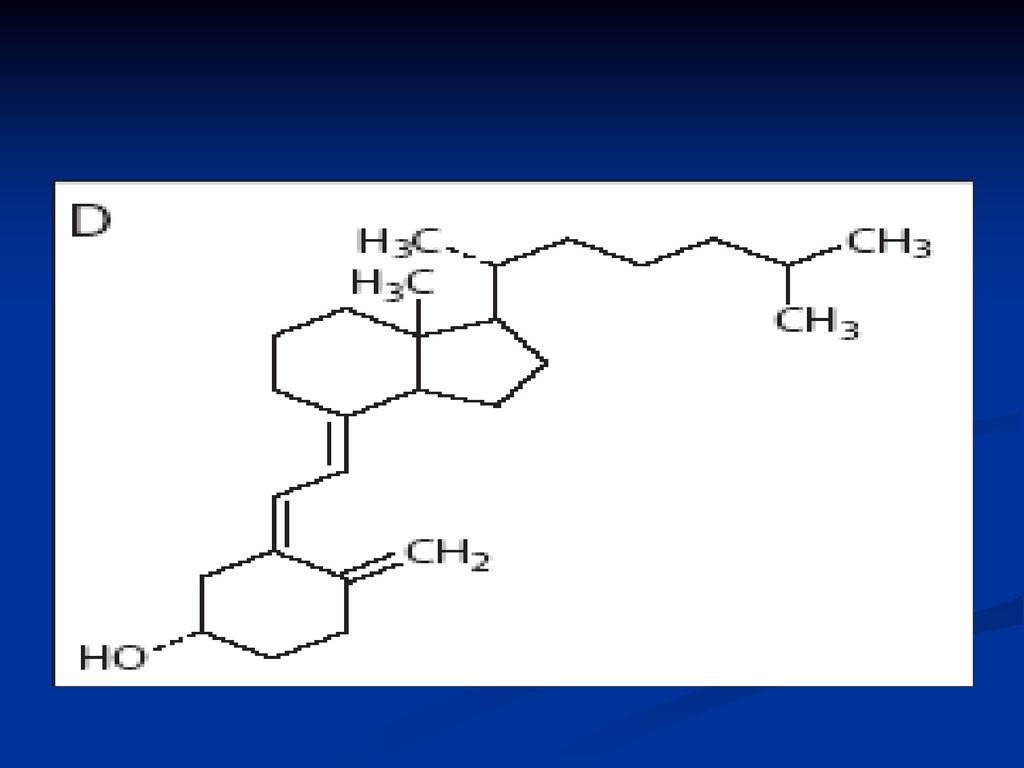
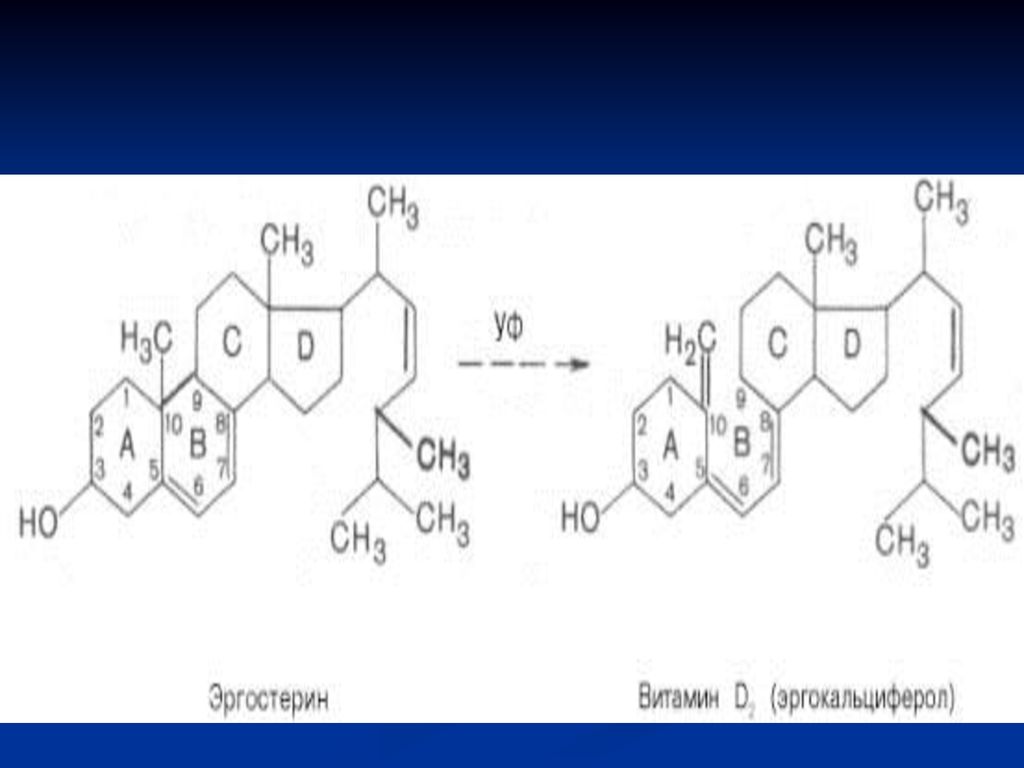
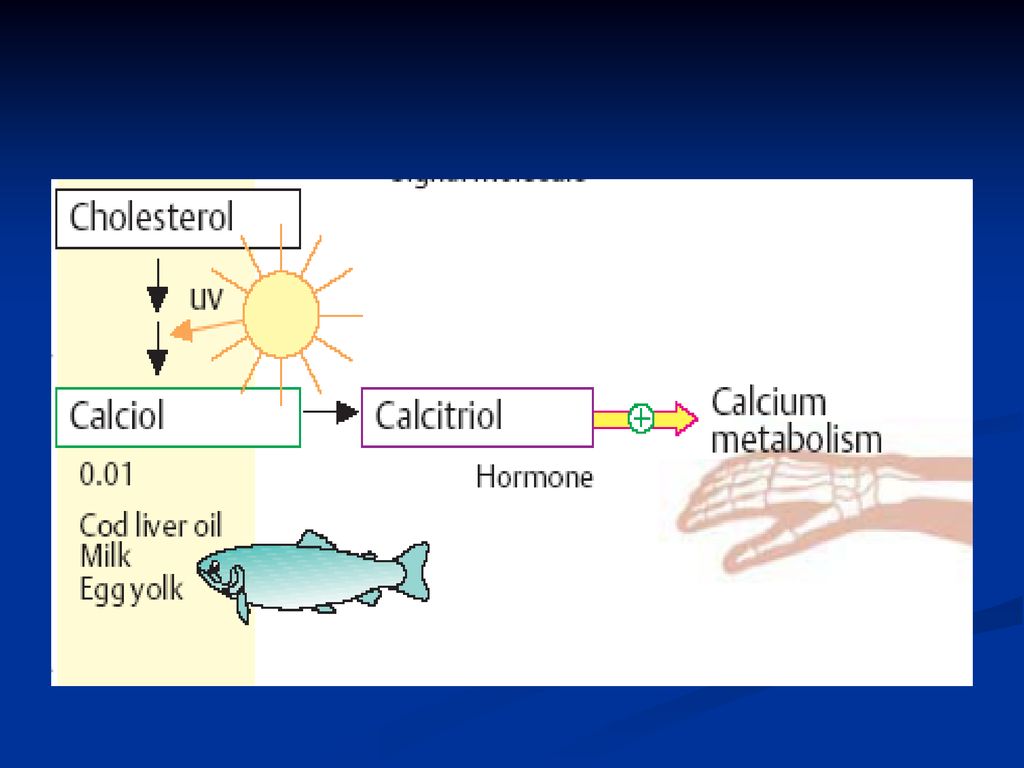
Vitamina D (antirahitică)
Se cunosc mai multe forme: D2 – ergocalciferol (ergosterol) D3 – colecalciferol Structura chimică: derivaţi ai ciclului steran:
 D3 – colecalciferol. Structura chimică: derivaţi ai ciclului steran:")
212
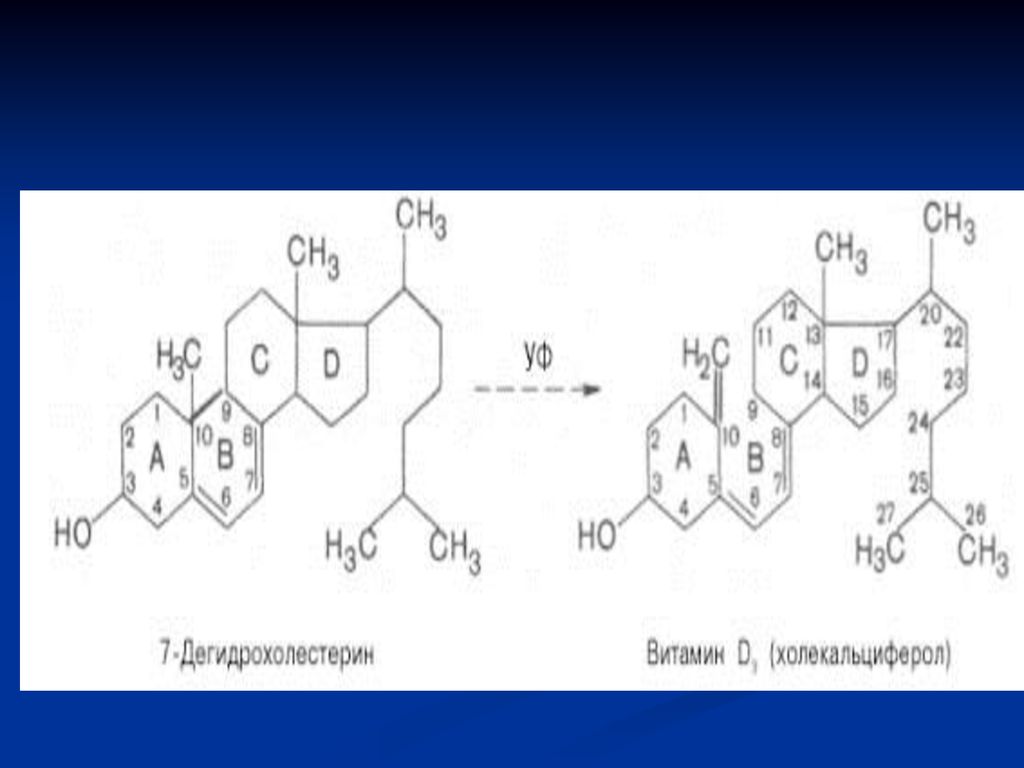
Metabolism: În intestin se supune acţiunii emulsionante a bilei, apoi este absorbită la nivelul intestinului subţire – în sânge (se leagă de o proteină) – D2 şi D3 - transportate la ficat. Prin sânge – în ficat ajunge şi vit D3 formată din 7- dehidrocolesterol în piele, sub acţiunea razelor ultraviolete din lumina solară.
 – D2 şi D3 - transportate la ficat. Prin sânge – în ficat ajunge şi vit D3 formată din 7- dehidrocolesterol în piele, sub acţiunea razelor ultraviolete din lumina solară.")
213
Vitamina D (antirahitică)
În ficat vit D3 suferă o hidroxilare – 25- hidroxicolecalciferol (E- colecalciferol-hidroxilază) - forma cea mai abundentă de vit D în circulaţia sanguină ţi ficat. În rinichi 25- hidroxicolecalciferol suferă o nouă hidroxilare – 1,25-dihidroxicolecalciferol (E- 1α-hidroxilază) – transportat la ţesuturile ţintă: intestin şi oase. Are rol important în menţinerea nivelului Ca seric. Reglarea – PTH şi nivelul de Ca în sânge.
 - forma cea mai abundentă de vit D în circulaţia sanguină ţi ficat. În rinichi 25- hidroxicolecalciferol suferă o nouă hidroxilare – 1,25-dihidroxicolecalciferol (E- 1α-hidroxilază) – transportat la ţesuturile ţintă: intestin şi oase. Are rol important în menţinerea nivelului Ca seric. Reglarea – PTH şi nivelul de Ca în sânge.")
215
Efectele vitaminei D La nivel de intestin: măresc absorbţia Ca şi fosfatului (stimulează sinteza unei proteine ce facilitează transportul Ca prin mucoasa intestinală) La nivelul tubilor renali: favorizează reabsorbţia fosfaţilor (contracarând eliminarea lor stimulată de PTH) La nivelul osului: intervine în mineralizarea oaselor prin mărirea formării de hidroxiapatită.
 La nivelul tubilor renali: favorizează reabsorbţia fosfaţilor (contracarând eliminarea lor stimulată de PTH) La nivelul osului: intervine în mineralizarea oaselor prin mărirea formării de hidroxiapatită.")
216
Vitamina D (antirahitică)
Carenţa acestei vitamine are ca consecinţă apariţia şi dezvoltarea rahitismului la copii din cauza dereglării metabolismului calciului şi fosforului – mineralizarea întîrziată a oaselor. este afectat scheletul: oasele sunt mai moi, picioarele se deformează sub greutatea corpului; fontanelele se închid cu întârziere, se reţine dezvoltarea dinţilor; tonusul muscular este redus etc. În sânge scade conţinutul calciului şi fosfatului.

217
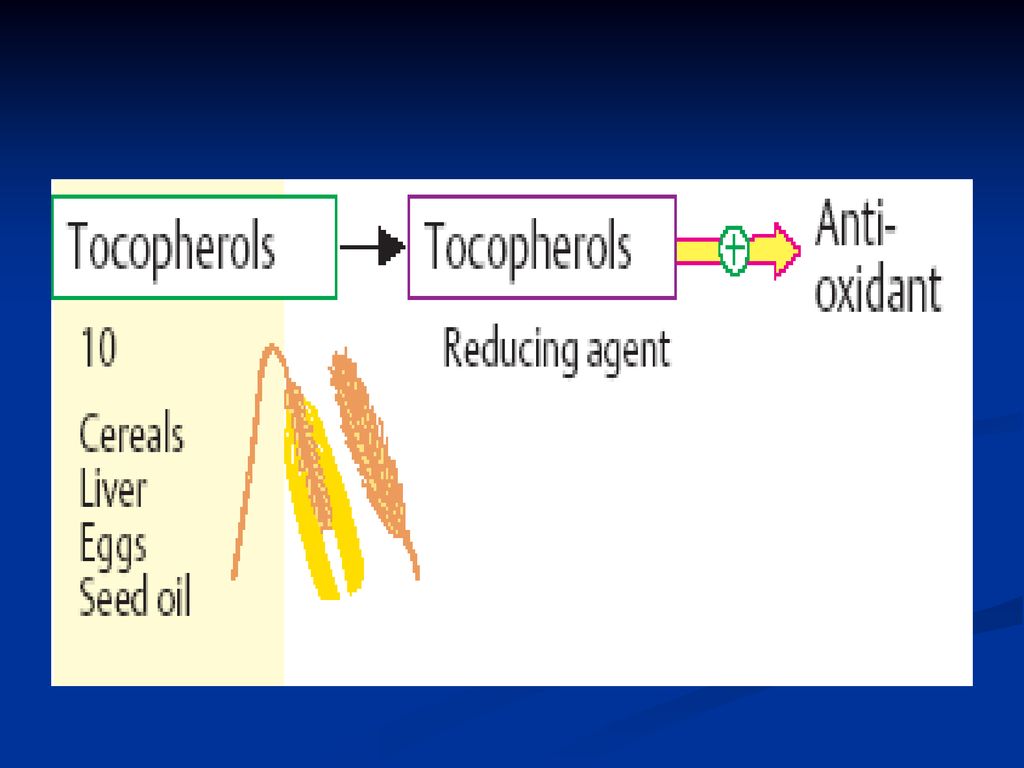
Vitamina E (-tocoferol, antisterilă)
Structura chimică: are la bază tocolul (5,7,8-trimetiltocol)
")
218
Metabolism Absorbită la nivelul intestinului subţire - sânge-ficat şi alte organe Se excretă prin bilă şi intestin (în prealabil se supine unei oxidări şi se conjugă cu a glucuronic)
")
219
Rolul vitaminei E Asupra organelor de reproducere: - măreşte fertilitatea la femei şi ajută la restaurarea activităţii sexuale la bărbaţi Este antioxidant puternic - protejează oxidarea AG nesaturaţi în MB intervine în metabolismul seleniului (care întră în componenţa glutation peroxidazei ce asigură protecţia membranelor faţă de acţiunea radicalilor peroxizi). Sunt implicate în sistemul E respiratorii, favorizând cuplarea fosforilării oxidative
. Sunt implicate în sistemul E respiratorii, favorizând cuplarea fosforilării oxidative.")
220
Deficienţa Vitaminei E
are ca urmare afectarea aparatului reproductiv: stagnarea embriogenezei, modificări degenerative a organelor reproductive ce duc la sterilitate. distrofie musculară, infiltrare adipoasă a ficatului, Distrugerea globulelor roşii Reducerea stabilităţii membranelor şi sărăcirea în colagen a ţesutului conjuctiv Ateroscleroză, cancer.

222
Se conţine în uleiuri vegetale (porumb, floarea soarelui etc
Se conţine în uleiuri vegetale (porumb, floarea soarelui etc.) carne, unt, gălbenuş de ou, varză, cereale. Necesarul: Zilnic: 7mg la adult şi 2-3 mg la copii
 carne, unt, gălbenuş de ou, varză, cereale. Necesarul: Zilnic: 7mg la adult şi 2-3 mg la copii.")
223
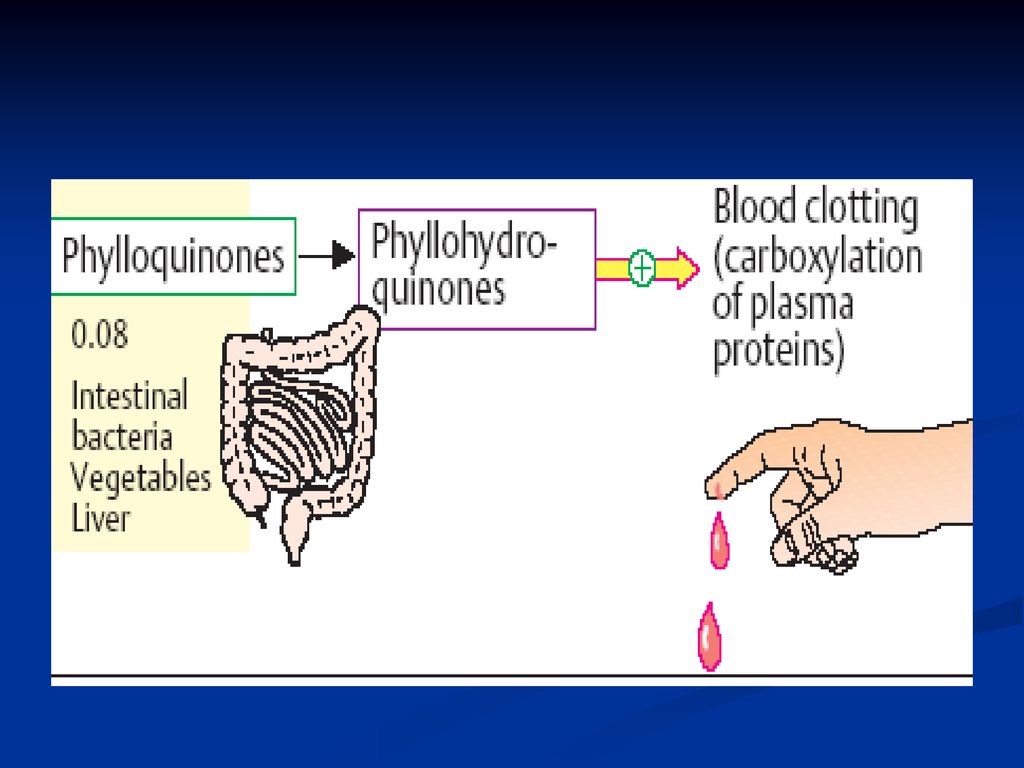
Vitamina K (antihemoragică).
K1- fitochinona (izolată din vegetale) K2- fariochinona (izolată din ţesuturile animale) La baza structurii: nucleul p-naftochinonei
 K2- fariochinona (izolată din ţesuturile animale) La baza structurii: nucleul p-naftochinonei.")
224
Vitamina K (antihemoragică).
.")
225
Rolul Participă la coagularea sângelui – este direct implicată în biosinteza factorilor II, VII, IX, X. Vitamina K ajută la modificarea restului de Clu în γ-carboxiglutamic (se adaugă o grupă carboxil ce poate fixa Ca) Intervine şi în procesele de fosforilare
 Intervine şi în procesele de fosforilare.")
226
Vitamina K (antihemoragică).
Carenţa acestei vitamine are ca consecinţă apariţia hemoragiilor în diferite organe şi ţesuturi din cauza dereglării procesului de coagulare a sângelui. La om hipovitaminoza în condiţii obişnuite se întâlneşte relativ rar deoarece această vitamină se sintetizează de microflora intestinală. Hemoragiile pot apărea când bolnavul utilizează cantităţi considerabile de antibiotici ”per os”, care vor distruge microflora (disbacterioză) intestinală, sau când este dereglat mecanismul de absorbţie a lipidelor.
 intestinală, sau când este dereglat mecanismul de absorbţie a lipidelor.")
227
Vitamina K (antihemoragică).
Antivitaminele acestei vitamine, de ex. dicumarolul sunt utilizate în clinică ca anticoagulanti. La baza structurii acestei vitamine este ciclul 1,4-naftochinonă. Există analogi sintetici hidrosolubili a acestei vitamine, de ex. vicasolul.

229
substanţe lipotrofe Ficatul este un organ specific de sinteză a fosfolipidelor plasmei. În lipsa colinei sau a grupelor CH3, necesare pentru sinteza lor, se diminuează viteza de sinteză a fosfolipidelor şi, respectiv, viteza cu care AG sunt eliminaţi din ficat. În consecinţă, are loc depozitarea lipidelor în ficat. Orice substanţă donatoare de CH3 pentru sinteza colinei este denumită lipotropă.

230
Dislipoproteidemie sau dislipidemie
modificarea c% lipidelor totale plasmatice, a uneia dintre fracţiuni sau modificarea raportului diverselor componente. Se clasifică: Hiperlipiproteidemii (primare şi secundare) Hipolipoproteidemii primare (ereditare, familiale)
 Hipolipoproteidemii primare (ereditare, familiale)")
231
Hiperlipidemiile primare sunt de natură genetică,
Hiperlipidemiile secundare sunt determinate de diferite afecţiuni renale, diabet, hipotireoză, intoxicaţii de alcool.

232
HLP primare după Fredrickson, în funcţie de aspectul fenotipic , HLP primare au fost clasificate în şase tipuri, I-VI, tipul II avînd două variante IIa şi IIb

233
Tipul Vârsta Aspecte biochimice Clinic I copilărie hiperCM hiperTg HiperCol Xantoame eruptive, hepatospleno-megalie IIa La orice vârstă LDL ↑ hiperCs Xantomatoză Ateroscleroză prematură IIb LDL, VLDL ↑ hiperTg hiperCs Obezitate, lipsesc xantoamele, Ateroscleroză prematură

234
Tipul Vârsta Aspecte biochimice Clinic III adult IV IDL hiperTg
hiperCol Xantoame eruptive ateroscleroza accelerată IV VLDL ↑ Obezitate, hepatospleno-megalie, hiperuricemie, ateroscleroză prematură V adolescenţă CM↑ VLDL ↑ Xantoame ateroscleroză prematură VI HDL ↑; diminuarea Col VLDL şi LDL. creşterea uşoară a longevităţii şi protecţie anti ATS

235
Hiperlipidemiile secundare
Diabetul zaharat: Lipsa de insulină: lipoliza activată, lipogeneza inhibată se manifestă prin diminuarea lipoproteinlipazei celulare, iar nivelul mare de AG amplifică producerea de pre-β-LP. apariţia hiperlipidemiei (II , IV)
")
236
Hiperlipidemia alcoolică:
se observă o lipidemie alimentară o dată cu consumul de alcool; e însoţită şi de o secreţie intensă a pre-β-LP, de o hiperchilomicronemie - ↑Tg un clirens lent al ambelor particule Steatoza hepatică

237
Alcoolul amplifică sinteza şi secreţia pre-β-lipoproteinelor determinate de valorile majorate ale NADH+H (formate la metabolizarea alcoolului sub acţiunea alcool DH). NADH +H inhibă oxidarea AG şi măreşte sinteza TAG şi a pre-β-lipoproteinelor (VLDL). Alcoolul favorizeză inducţia enzimelor microzomale în ficat.
. Alcoolul favorizeză inducţia enzimelor microzomale în ficat.")
238
Obezitatea Excesul de aport caloric, raportat la cheltuielile energetice ale organismului conduce la activarea marcată a lipogenezei şi dezvoltarea excesivă a ţesutului adipos IMC=greutatea (kg)/ înălţime (m²) =20-25 Obezitatea abdominală – depunerea ţesutului adipos în jurul organelor din cavitatea peritoneală. Lipoliză excesivă – creşterea AGL. Este asociată cu IR, modificări lipidice (hiperTg, ↑ VLDL; ↓HDL); infiltraţia grasă a ficatului, DZ tip 2. Markerul ei : T/Ş.
/ înălţime (m²) = Obezitatea abdominală – depunerea ţesutului adipos în jurul organelor din cavitatea peritoneală. Lipoliză excesivă – creşterea AGL. Este asociată cu IR, modificări lipidice (hiperTg, ↑ VLDL; ↓HDL); infiltraţia grasă a ficatului, DZ tip 2. Markerul ei : T/Ş.")
239
Ateroscleroza Se caracterizează prin depunerea Col în intima vaselor sub formă de plăci aterosclerotice, ceea ce duce la îngustarea lumenului capilarelor şi împedică fluxul circulator normal. Se reduce elasticitatea pereţilor arteriali favorizează BIC şi a accidentelor vasculare cerebrale. Factorii favorizanţi: supraalimentaţia, sedentarismul, stresul, obezitatea, fumatul ce determină: Scăderea nr de receptori pentru LDL (↑ Col total în plasmă şi creşterea Col din LDL) ↓ HDL ↑IDL, ↑ LDL ↑Tg
 ↓ HDL. ↑IDL, ↑ LDL. ↑Tg.")
240
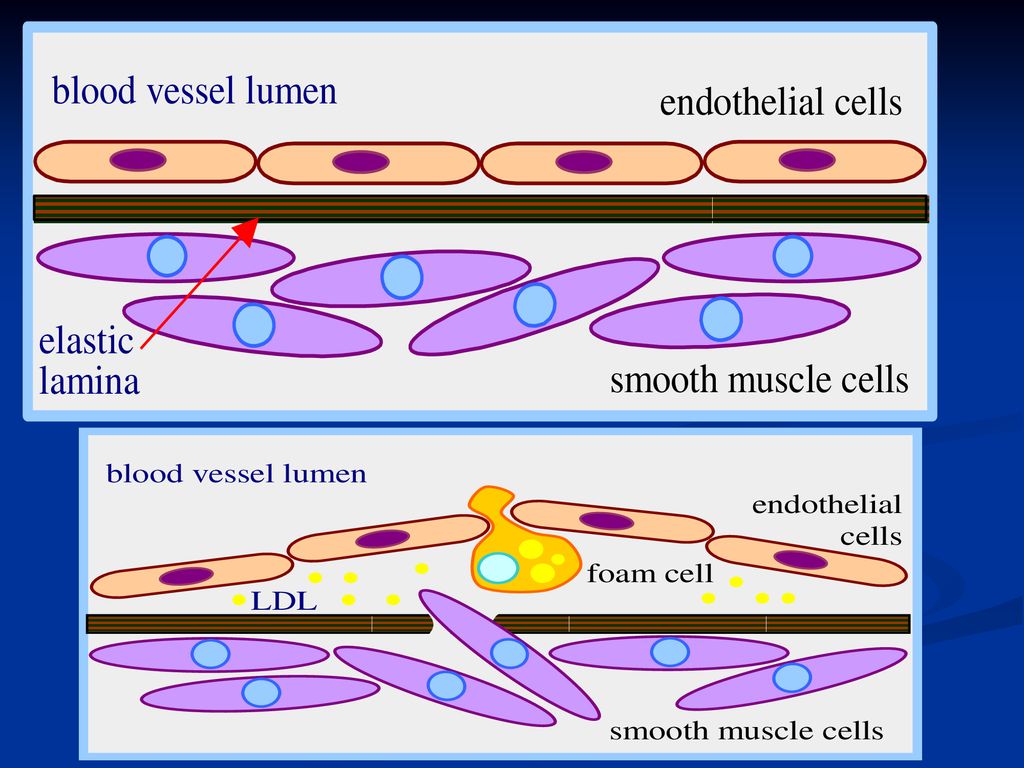
Ateroscleroza Patogenie: leziunea pereţilor endoteliali
Infiltrarea LDL oxidate Proliferarea “celulelor spumoase” Formarea unor depozite lipidice la nivelul pereţilor arteriali formate din: cristale de Col şi esteri ai Col; “celule spumoase”; celule necrotizate. Alte modificări: proliferarea fibrelor musculare netede, infiltrarea macrofagelor şi a limfocitelor, eliberarea locală de citokine; agregarea trombocitelor şi extinderea leziunilor endoteliale iniţiale.

242
Hipolipoproteidemiile primare (ereditare, familiale)
sunt datorate genelor care controlează sinteza, transportul sau utilizarea LP. afecţiunea Tangier, A beta-lipoproteinemia familială;

243
Patologia lipidelor afecţiunea Tangier - cauzată de insuficienţa sau lipsa totală de α-LP (HDL), datorate deficitului sintezei de apo A acumularea în ţesuturi a esterilor colesterolului. Splenomegalie, anomalii neurologice, hipertrofie amigdaliană Afecţiunea este incurabilă.

244
A beta-lipoproteinemia familială
determinată de o sinteză defectuoasă a apo-B şi apo-C (din CM şi VLDL) Reducerea Tg, Col, FL; sunt absente LDL, VLDL, CM Apare malabsorbţia lipidelor
 Reducerea Tg, Col, FL; sunt absente LDL, VLDL, CM. Apare malabsorbţia lipidelor.")
245
Lipidoze tisulare Boli ereditare, caracterizate prin acumularea de sfingolipide în creier şi ţesuturi, datorate deficienţei enzimelor implicate în catabolizarea lor (scindarea are loc în lisosomi)
")
246
Lipidoze tisulare 1. Neimann-Pick - deficienţa de sfingomielinază
sfingomielina SM- se acumulează în ficat, splină, creier Hepato- şi splenomegalie, tulburări digestive, întîrziere psihomotorie, convulsii Decedează în vârsta fragedă

247
Lipidoze tisulare 2. afecţiunea Tay-Sachs – lipsa β-N-acetilhexozaminidazei (hidrolază ce scindează N acetil-galactozamina din GM2) Acumularea de GM2 - tumefierea celulelor ganglionare ale scoarţei creierului – creşte în volum pe retina ochiului – apar pete de culoare roşie-vişinie – gangliozid GM2

248
Lipidoze tisulare 3. Boala Krabbe – deficit de galactocerebrozidază
Demielinizări la nivelul creierului Paralizie, convulsii, cecitate, surditate, deces în vârsta fragedă

249
Lipidoze tisulare 4. boala Gaucher – deficit de hidrolază ce scindează glucoza din glucocerebrozide Splenomegalie (se acumulează în splină); tulburări hematologice ( membrana eritrocitelor are cantităţi mari de gangliozide) Osteoporoză, retard mintal
; tulburări hematologice ( membrana eritrocitelor are cantităţi mari de gangliozide) Osteoporoză, retard mintal.")
250
Lipidoze tisulare 5. leucodistrofia metacromatică – deficit de sulfatidază Acumularea sulfatidelor în substanţa albă nervoasă, ficat, rinichi Tulburări neurologice şi psihice

251
Lipidoze tisulare boala lui Fabry (lipogranulomatoza) - deficienţa ceramidazei lizozomale (acide) cauzează acumulări de ceramide. moştenită autosomal recesiv leziuni granulomatoase în piele, articulaţii şi laringe, disfuncţii moderate a sistemului nervos; de asemenea poate implica inima, plămînii, şi nodulii limfatici. De obicei fatală în timpul primilor câţiva ani de viaţă.

Παρόμοιες παρουσιάσεις












>")






