Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
03. Επιδιόρθωση δίκλωνων ρήξεων και ανασυνδυασμός

2
Επιδιόρθωση δίκλωνων ρήξεων Μια μονόκλωνη εντομή σε ένα δίκλωνο μόριο δεν αποτελεί κρίσιμο πρόβλημα για το κύτταρο επειδή η διπλή έλικα παραμένει ακέραιη

3
Επιδιόρθωση δίκλωνων ρήξεων Ανασυνδυασμός Μη ομόλογη σύνδεση άκρων

4
1. H αποσυγκρότηση της αντιγραφικής διχάλας (fork collapse) Όταν η λιγάση δεν έχει ακόμη κλείσει μια εγκοπή
 Όταν η λιγάση δεν έχει ακόμη κλείσει μια εγκοπή.")
5
Το μονοπάτι της επανεκκίνησης Σχηματισμός μονόκλωνης ουράς 3’-ΟΗ Διείσδυση κλώνου Μετανάστευση διακλάδωσης Λύση κόμβου Holliday

6
Πώς μελετάμε την επιδιόρθωση των δίκλωνων ρήξεων; Μελέτη μεταλλαγμάτων της E. coli που αδυνατούν να πραγματοποιήσουν γενετικό ανασυνδυασμό –recA –recB, recC, recD

7
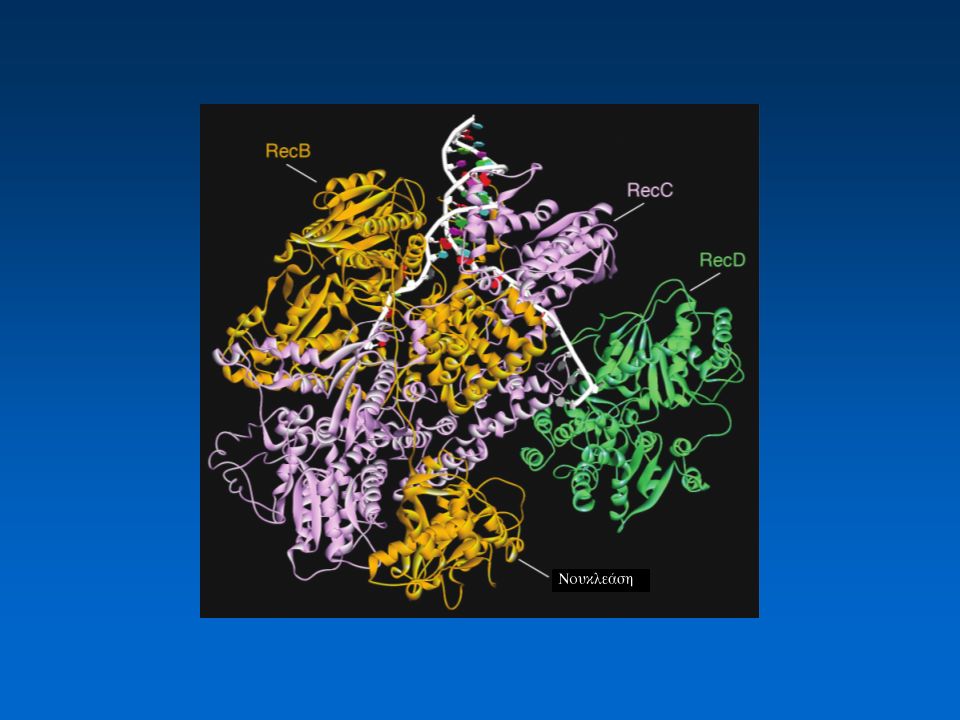
To σύμπλοκο RecBCD Αποπεριέλιξη του λείου άκρου του DNA –RecB ATP-dependent helicase, κατεύθυνση 3’ → 5’ ενεργότητα νουκλεάσης (3’ → 5’ και 5’ → 3’) –RecD ATP-dependent helicase, κατεύθυνση 5’ → 3’ –RecC αλληλεπιδρά και σταθεροποιεί τις RecB/RecD
 –RecD ATP-dependent helicase, κατεύθυνση 5’ → 3’ –RecC αλληλεπιδρά και σταθεροποιεί τις RecB/RecD")
9
Η επεξεργασία των δίκλωνων ρήξεων από το σύμπλοκο RecBCD Το σύμπλοκο RecBCD έχει δράση νουκλεάσης και ελικάσης. Προσδένεται στο λείο άκρο του DNA, καθοδικά μιας αλληλουχίας chi, ξετυλίγει το δίκλωνο μόριο και αποικοδομεί τον ένα κλώνο με κατεύθυνση 3'- 5', κινούμενο προς τη θέση chi. Αφού το σύμπλοκο RecBCD περάσει από τη θέση chi, η RecB αλλάζει στερεοδιαμόρφωση, υδρολύοντας την άλλη αλυσίδα, δημιουργώντας τελικά ένα 3’ ελεύθερο άκρο. Το σύμπλοκο RecBCD συμβάλλει στη φόρτωση της RecA στη νεοσχηματιζόμενη μονόκλωνη ουρά

10
Η RecA καταλύει τη διείσδυση του κλώνου Το άκρο του κλώνου που διεισδύει εκτοπίζει τον κλώνο του ομολογου δίκλωνου DNA που φέρει την ίδια αλληλουχία με αυτό Έτσι προκαλείται μια ανταλλαγή κλώνων που οδηγεί στο σχηματισμό ενός βρόχου εκτόπισης Το σημείο διακλάδωσης όπου διασταυρώνεται ο κλώνος που πραγματοποίησε τη διείσδυση με τον κλώνο που εκτόπισε μπορεί να μεταναστεύσει

11
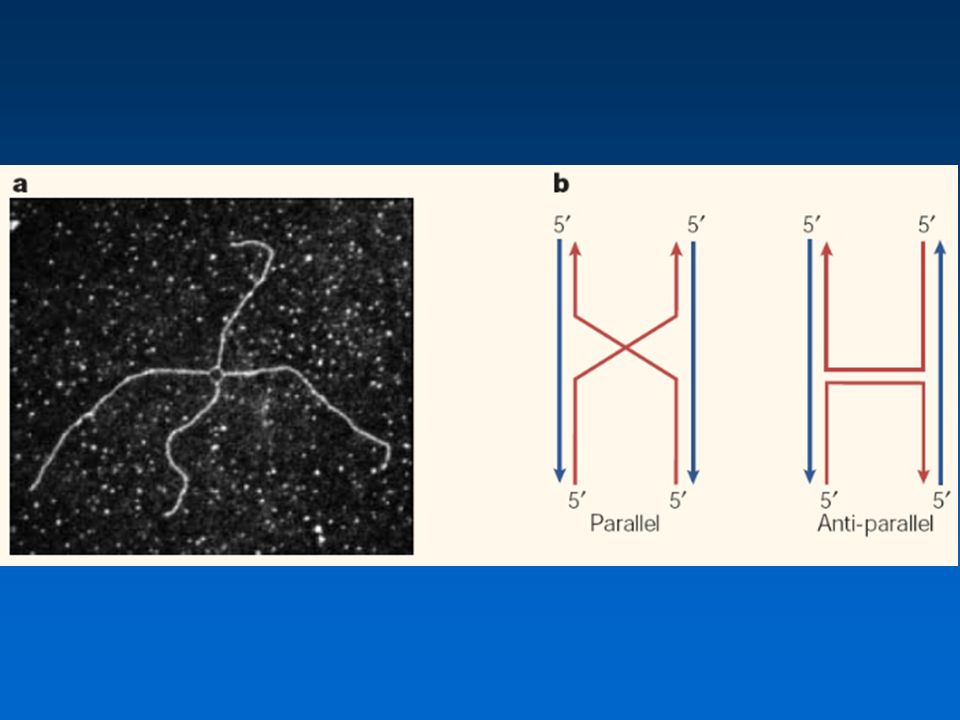
Μετανάστευση διακλάδωσης Το μοντέλο ομόλογου ανασυνδυασμού του Holliday

14
Οι πρωτεΐνες Ruv καταλύουν τη μετανάστευση της διακλάδωσης ενός κόμβου Holliday στην E. coli Το σύμπλοκο Ruv δρα πάνω στους κόμβους ανασυνδυασμού. Η RuvA αναγνωρίζει τη δομή του κόμβου και η RuvB είναι μια ελικάση που καταλύει τη μετανάστευση διακλάδωσης. Η RuvC αποσυγκροτεί τους κόμβους (resolvase), ώστε να δημιουργηθούν τα προϊόντα του ανασυνδυασμού.
, ώστε να δημιουργηθούν τα προϊόντα του ανασυνδυασμού..")
15
O επανασχηματισμός της αντιγραφικής διχάλας είναι δυνατός επειδή η σύνθεση του DNA μπορεί να ξεκινήσει σε άλλες θέσεις έναρξης εκτός από oriC Η φόρτωση της DnaB πραγματοποιείται με τη συμβολή της PriA

16
Αν ο δρόμος RecBCD αδρανοποιηθεί τότε αναλαμβάνει δράση το σύστημα RecE. Αν αδρανοποιηθεί και το RecE τότε τελευταία εφεδρεία είναι το RecF.

17
2. Ο μιτωτικός ανασυνδυασμός Στη φάση S οι δύο χρωματίδες είναι πολύ κοντά, έτσι μέσω ομόλογου ανασυνδυασμού το κύτταρο μπορεί να επιδιορθώσει μια δίκλωνη ρήξη στη μία χρωματίδα με βάση την αλληλουχία της αδελφής χρωματίδας. Θα μπορούσε αυτό να γίνει και με βάση το ομόλογο χρωμόσωμα, όμως αυτό είναι πολύ μακρύτερα.

18
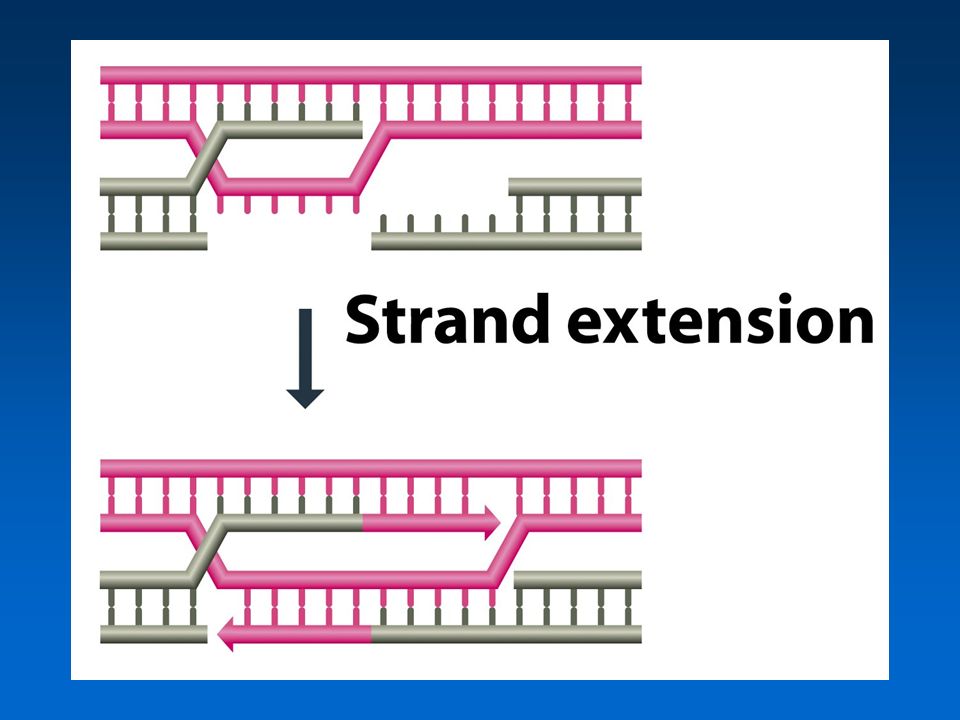
Δύο μονοπάτια επιδιόρθωσης δίκλωνων ρήξεων στους ευκαρυώτες: –Εξαρτώμενη από τη σύνθεση DNA υβριδοποίηση του κλώνου –Επιδιόρθωση δίκλωνης ρήξης

19
Βήμα 1: Επεξεργασία κομμένων άκρων

20
Βήμα 2: Σχηματισμός νουκλεοπρωτεϊνικών ινιδίων και διείσδυση κλώνου

21
Βήμα 3α: Εξαρτώμενη από DNA υβριδοποίηση κλώνου

22
Βήμα 3β: Επιδιόρθωση δίκλωνης ρήξης

23
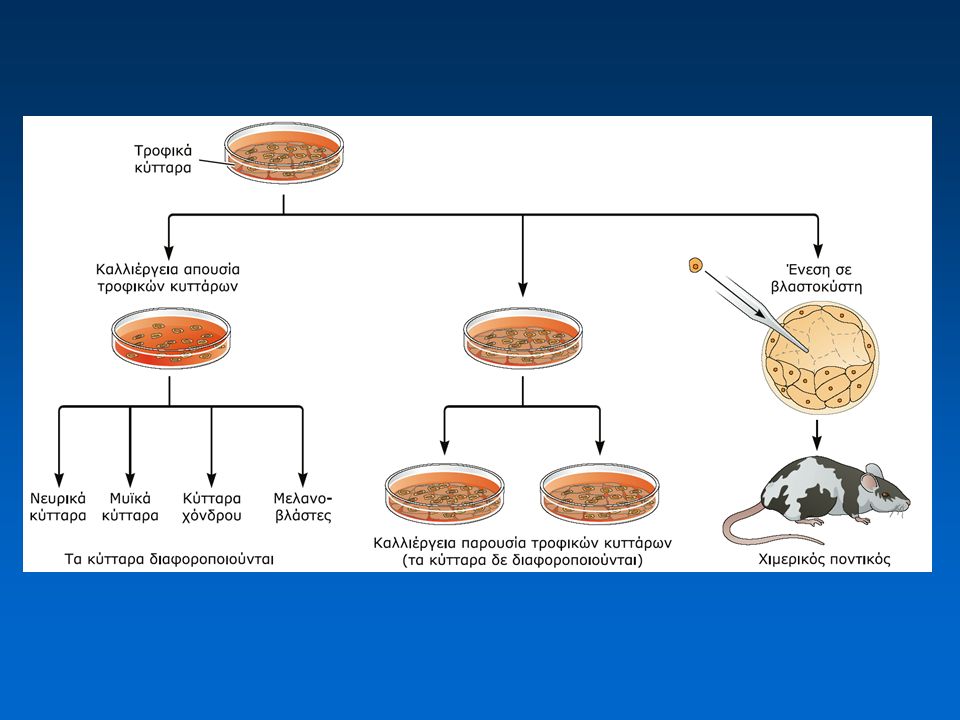
3. Γονιδιακή στόχευση (gene targeting), αδρανοποίηση γονιδίων-στόχων (gene knockout) Απομόνωση βλαστικών κυττάρων (ES cells) Εισαγωγή εξωγενούς DNA Επιλογή κλώνων με ενσωματωμένο το εξωγενές DNA Επιλογή κλώνων στους οποίους το ενσωματωμένο DNA έχει υποκαταστήσει το ενδογενές τμήμα Εισαγωγή των γενετικά τροποποιημένων ES κυττάρων σε πρώιμα έμβρυα ποντικών, δημιουργία διαγονιδιακών ποντικών
, αδρανοποίηση γονιδίων-στόχων (gene knockout) Απομόνωση βλαστικών κυττάρων (ES cells) Εισαγωγή εξωγενούς DNA Επιλογή κλώνων με ενσωματωμένο το εξωγενές DNA Επιλογή κλώνων στους οποίους το ενσωματωμένο DNA έχει υποκαταστήσει το ενδογενές τμήμα Εισαγωγή των γενετικά τροποποιημένων ES κυττάρων σε πρώιμα έμβρυα ποντικών, δημιουργία διαγονιδιακών ποντικών.")
24
Δημιουργία διαγονιδιακών ποντικών

25
Η άμεση μεταφορά DNA στους προπυρήνες γονιμοποιημένων ωαρίων ποντικού είναι αρκετά αποδοτική, όμως δεν έχουμε τη δυνατότητα επιλογής της θέσης ένθεσης του DNA ή τη δυνατότητα ελεγχόμενης τροποποίησης συγκεκριμένων ενδογενών γονιδίων Εισαγωγή DNA σε εμβρυϊκά βλαστικά κύτταρα → επιλογή των κυττάρων με τα ζητούμενα χαρακτηριστικά → ενσωμάτωση στο αναπτυσσόμενο έμβρυο Ομόλογος ανασυνδυασμός 25

26
Παραγωγή εμβρυϊκών βλαστικών κυττάρων (Embryonic Stem cells, ES) από βλαστοκύστεις ποντικού
 από βλαστοκύστεις ποντικού")
28
Ομόλογος ανασυνδυασμός Ο ομόλογος ανασυνδυασμός αποτελεί ισχυρό εργαλείο για τη γονιδιακή στόχευση Όμως: Είναι σχετικά σπάνιος στα θηλαστικά (1 ομόλογος : 1000 ετερόλογους) Με κατάλληλη τεχνολογία: 5-20% επί του συνόλου των ενθέσεων
 Με κατάλληλη τεχνολογία: 5-20% επί του συνόλου των ενθέσεων")
29
Δημιουργία HGPRT knockouts HGPRT: Hypoxanthine-guanine phosphoribosyltransferase. Είναι μια τρανσφεράση που καταλύει τη μετατροπή της υποξανθίνης σε φωσφορική ινοσίνη και της γουανίνης σε φωσφορική γουανίνη. Η HGPRT παίζει σημαντικό ρόλο στη δημιουργία των πουρινών μέσω του μονοπατιού της διάσωσης των πουρινών.

30
Θετική-αρνητική επιλογή Τυχαία ένθεση: κύτταρα ανθεκτικά στο G418 και ευαίσθητα στη φιαλουριδίνη Στοχευμένη ένθεση: κύτταρα ανθεκτικά στο G418 και ανθεκτικά στη φιαλουριδίνη

31
Knockout γονιδίου στα ποντίκια

32
4. Μη ομόλογη σύνδεση άκρων Στη φάση G1 και στην πρώιμη φάση S, δεν είναι διαθέσιμη η αδελφή χρωματίδα. Έτσι, οι δίκλωνες ρήξεις επιδιορθώνονται με μη- ομόλογη σύνδεση άκρων. Απαραίτητη η πυρηνική πρωτεΐνη Ku.

33
Non-homologous end-joining (NHEJ) Η δομή του ετεροδιμερούς Ku-DNA
 Η δομή του ετεροδιμερούς Ku-DNA")
34
Σύνδεση μη- ομόλογων άκρων (Non-homologous end-joining: NHEJ) στους ανθρώπους. Η διαδικασία επιδιόρθωσης

35
H σύνδεση των άκρων επιτελείται από τη DNA λιγάση IV, εφόσον προηγουμένως νουκλεάσες και DNA πολυμεράσες έχουν ‘λειάνει’ τα άκρα τους Η επεξεργασία αυτή συνήθως προκαλεί μεταλλαγές στο σημείο της ένωσης Ο μηχανισμός αυτός χρησιμοποιείται στο σχηματισμό των ώριμων γονιδίων ανοσοσφαιρίνης, των οποίων οι επί μέρους περιοχές βρίσκονται αρχικά σε ξεχωριστές θέσεις στο γονιδίωμα Η πρωτεΐνη Ku εντοπίστηκε ως στόχος των αυτο- αντισωμάτων ασθενών σκληροδέρματος (αυτοάνοση ασθένεια)
")
36
5. Ο μειωτικός ανασυνδυασμός Ο ομόλογος ανασυνδυασμός συμβαίνει κατά το παρατεταμένο στάδιο της πρόφασης στη μείωση

37
Ο ομόλογος ανασυνδυασμός συμβάλλει στην αύξηση της γενετικής ποικιλότητας μέσω της δημιουργίας χρωμοσωμάτων με νέους συνδυασμούς αλληλομόρφων Είναι επίσης απαραίτητος για το σωστό διαχωρισμό των ζευγών των ομόλογων χρωμοσωμάτων κατά την πρώτη μειωτική διαίρεση: αν δεν εξελιχθεί σωστά τα ομόλογα χρωμοσώματα δεν θα ζευγαρώσουν σωστά → διαχωρίζονται με ανώμαλο τρόπο και προκύπτουν ανευπλοειδίες

38
Η δημιουργία του ασκού στους μύκητες

39
Απόκλιση από την τετράδα: γονιδιακή μετατροπή

40
Μοντέλο δίκλωνων ρήξεων (εντομών) Δημιουργία δίκλωνης ρήξης (SPO11) σε ένα από τα δύο ανασυνδυαζόμενα μόρια Μια εξωνουκλεάση (EXO1 ή DNA2/helicase) δημιουργεί 3’ προεξέχοντα άκρα και στα δύο μέρη Ένα από τα δύο 3’ άκρα εισχωρεί στο ομόλογο DNA μόριο (Rad51/Dmc1)
 Δημιουργία δίκλωνης ρήξης (SPO11) σε ένα από τα δύο ανασυνδυαζόμενα μόρια Μια εξωνουκλεάση (EXO1 ή DNA2/helicase) δημιουργεί 3’ προεξέχοντα άκρα και στα δύο μέρη Ένα από τα δύο 3’ άκρα εισχωρεί στο ομόλογο DNA μόριο (Rad51/Dmc1)")
41
(SPO11) (EXO1 or DNA2+ helicase)
 (EXO1 or DNA2+ helicase)")
42
Rad51 and Dmc1

46
Το μοντέλο DSB θεωρείται σήμερα ως ικανοποιητική ερμηνεία του ομόλογου ανασυνδυασμού σε όλους τους οργανισμούς. Σε αυτό συνετέλεσε: –Κατά τη μείωση τα χρωμοσώματα υφίστανται δίκλωνες ρήξεις με 100-1000 φορές μεγαλύτερη συχνότητα από ότι στα βλαστικά κύτταρα –Ο ομόλογος ανασυνδυασμός εμπλέκεται στην επιδιόρθωση του DNA, ιδιαίτερα των εντομών που δημιουργούνται κατά την αντιγραφή

47
Ο όρος ‘ανασυνδυασμός’ αρχικά χρησιμοποιήθηκε από τους γενετιστές για να περιγράψει το αποτέλεσμα του επιχιασμού ανάμεσα σε ζεύγη ομόλογων χρωμοσωμάτων κατά τη μείωση Σήμερα πιστεύουμε ότι ο πρωταρχικός ρόλος του ανασυνδυασμού είναι η επιδιόρθωση του DNA

48
Η συχνότητα ανασυνδυασμού δεν είναι ίδια σε όλο το γονιδίωμα Η συχνότητα διαφέρει στα ωοκύτταρα και στο σπέρμα: Ο ανασυνδυασμός συμβαίνει δύο φορές συχνότερα στις γυναίκες απ' ό,τι στους άνδρες Η συχνότητα ανασυνδυασμού ποικίλλει σε διαφορετικές θέσεις του γονιδιώματος και εξαρτάται από τη δομή του χρωμοσώματος. Πχ, ο διασκελισμός καταστέλλεται στις περιοχές συμπυκνωμένης και ανενεργής ετεροχρωματίνης Ο ανασυνδυασμός συμβαίνει κατά το παρατεταμένο στάδιο της πρόφασης στη μείωση

49
Ομόλογος ανασυνδυασμός και επιδιόρθωση Λόγω της αρχικής συσχέτισης του ομόλογου ανασυνδυασμού με τη γενετική, οι αρχικές μελέτες αφορούσαν τη μείωση Όμως, στελέχη E. coli με ελαττώματα στο σύστημα RecBCD βρέθηκαν προβληματικά και ως προς την επιδιόρθωση του DNA Σήμερα πιστεύουμε ότι ο κύριος ρόλος του ανασυν- δυασμού είναι η μετα-αντιγραφική επιδιόρθωση

50
Επιδιόρθωση μονόκλωνου χάσματος μέσω RecF: όταν ένα τμήμα του γονιδιώματος φέρει πολλές βλάβες (πχ διμερή Τ), η DNA pol αναπηδά
, η DNA pol αναπηδά")
51
Αν η βλάβη δεν μπορεί να παρακαμφθεί, τότε η διχάλα αντιγραφής θα σταματήσει, θα οπισθοχωρήσει δημιουργώντας δίκλωνο ανάμεσα στα θυγατρικά μόρια. Στη συνέχεια η διχάλα θα συνεχίσει μπροστά με ένα μηχανισμό ανάλογο της μετανάστευσης της διακλάδωσης Holiday.

52
Ο βακτηριακός ανασυνδυασμός συμβαίνει ως αντίδραση επιδιόρθωσης που ενεργοποιείται από την παρουσία βλαβών στο DNA. Ο ανασυνδυασμός στους ευκαρυώτες συμβαίνει κατά τη μείωση σε επίπεδο ολόκληρου χρωμοσώματος. Παρόλα αυτά, και στις δύο περιπτώσεις πρέπει να εμπλέκονται παρόμοιες μοριακές διαδικασίες χειρισμού του DNA.

53
Τι να μελετήσετε: Tropp, Chapter 10 Genes VIII: –15.1 έως 15.4 –15.8 –15.10 –15.16-15.17 –15.20-15.22 –15.25-15.27 –15.29

Παρόμοιες παρουσιάσεις