Κατέβασμα παρουσίασης
Η παρουσίαση φορτώνεται. Παρακαλείστε να περιμένετε

1
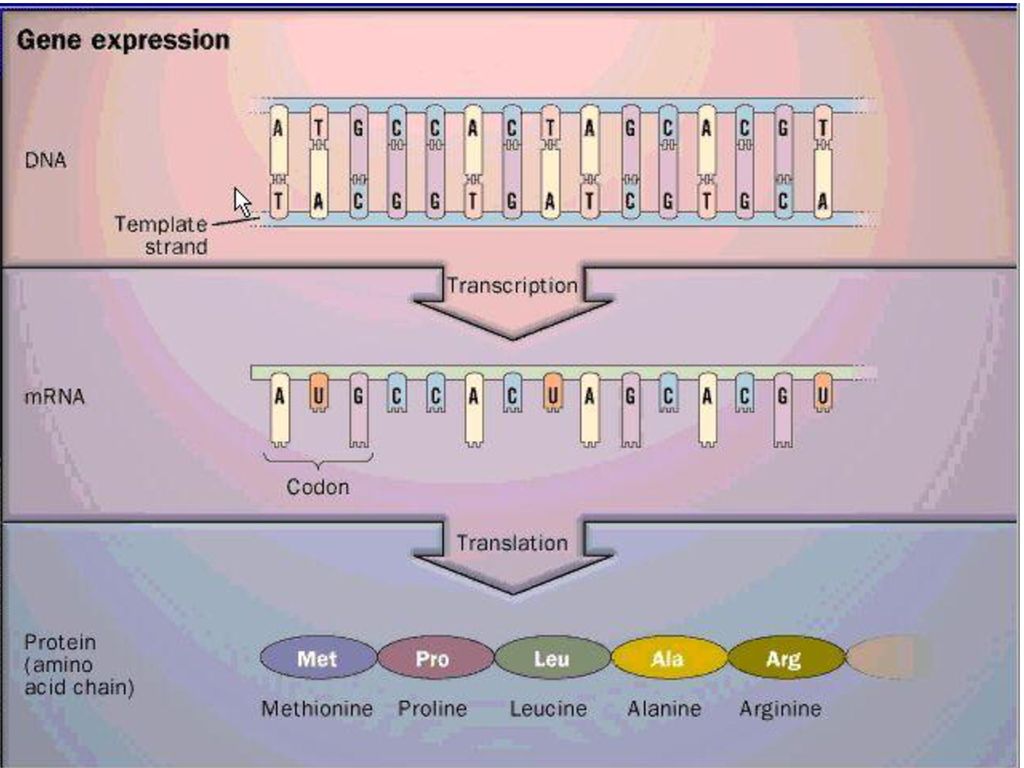
ΜΕΤΑΓΡΑΦΗ

3
Πρωτεΐνες Κυτταρικής Οικονομίας Διαφορική Γονιδιακή Έκφραση
vs. Μοναδικές Πρωτεΐνες Διαφορική Γονιδιακή Έκφραση

4
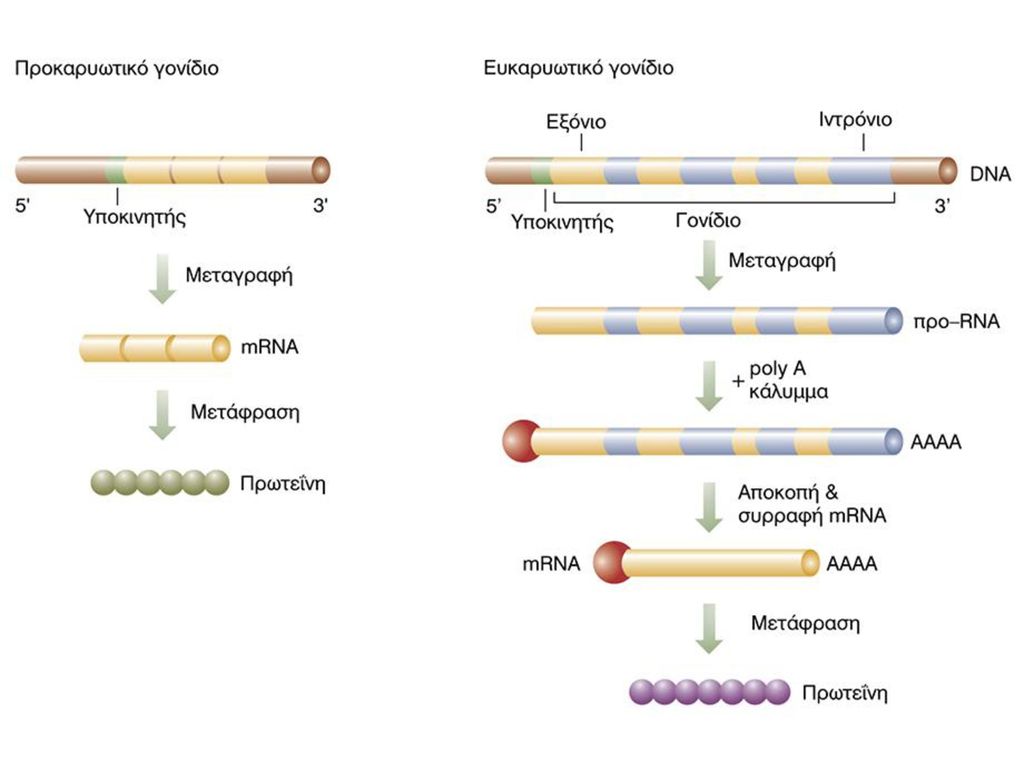
ΜΕΤΑΓΡΑΦΗ DNA ΣΕ RNA Ακρίβεια του σημείου έναρξης και λήξης Πολλά σημεία έναρξης/λήξης Πιστότητα μεταγράφου Απουσία μηχανισμών επιδιόρθωσης

5
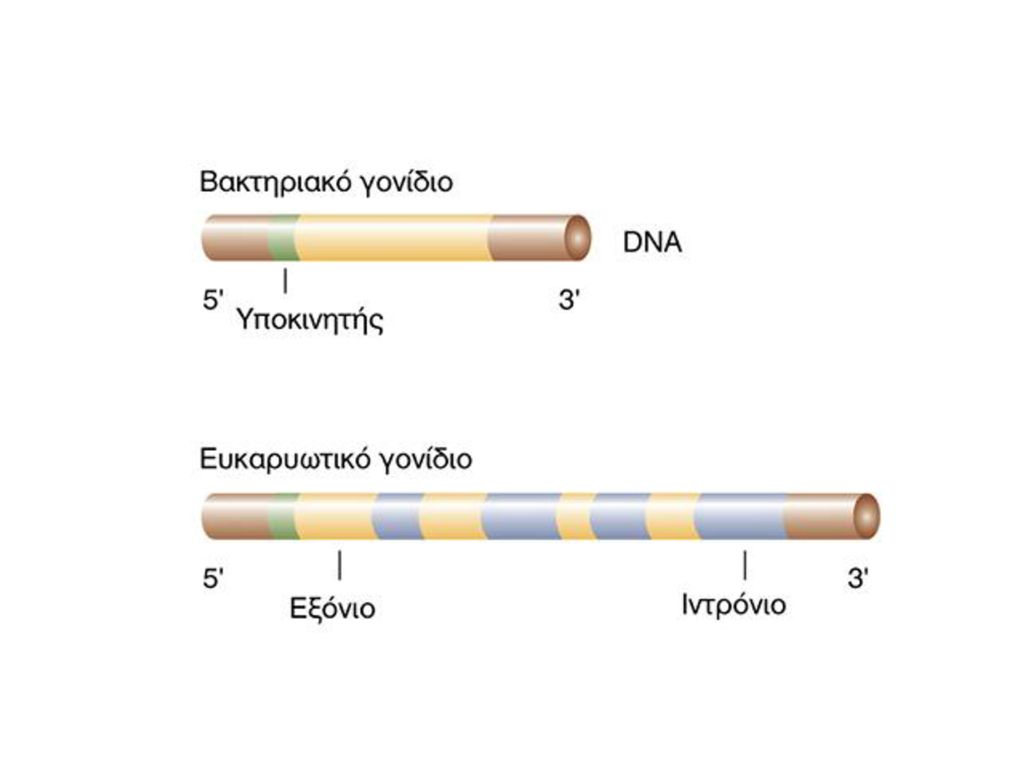
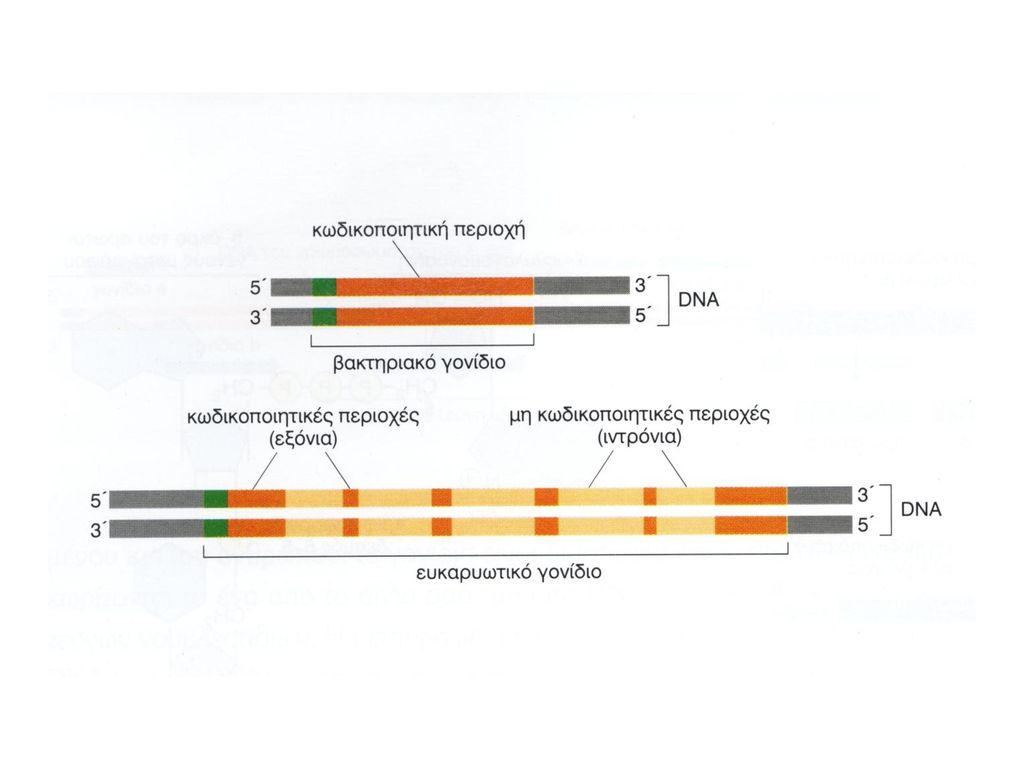
ΓΟΝΙΔΙΑ ΣΕ ΧΡΩΜΟΣΩΜΑΤΑ

6
ΔΙΑΦΟΡΙΚΗ ΕΚΦΡΑΣΗ ΓΟΝΙΔΙΩΝ ΔΟΓΜΑ «ΜΙΑ ΠΡΩΤΕΪΝΗ, ΕΝΑ ΓΟΝΙΔΙΟ»
ΠΟΙΟΤΙΚΕΣ ΔΙΑΦΟΡΕΣ ΔΟΓΜΑ «ΜΙΑ ΠΡΩΤΕΪΝΗ, ΕΝΑ ΓΟΝΙΔΙΟ»

7
ΔΙΑΦΟΡΙΚΗ ΕΚΦΡΑΣΗ ΓΟΝΙΔΙΩΝ
ΠΟΣΟΤΙΚΕΣ ΔΙΑΦΟΡΕΣ

8
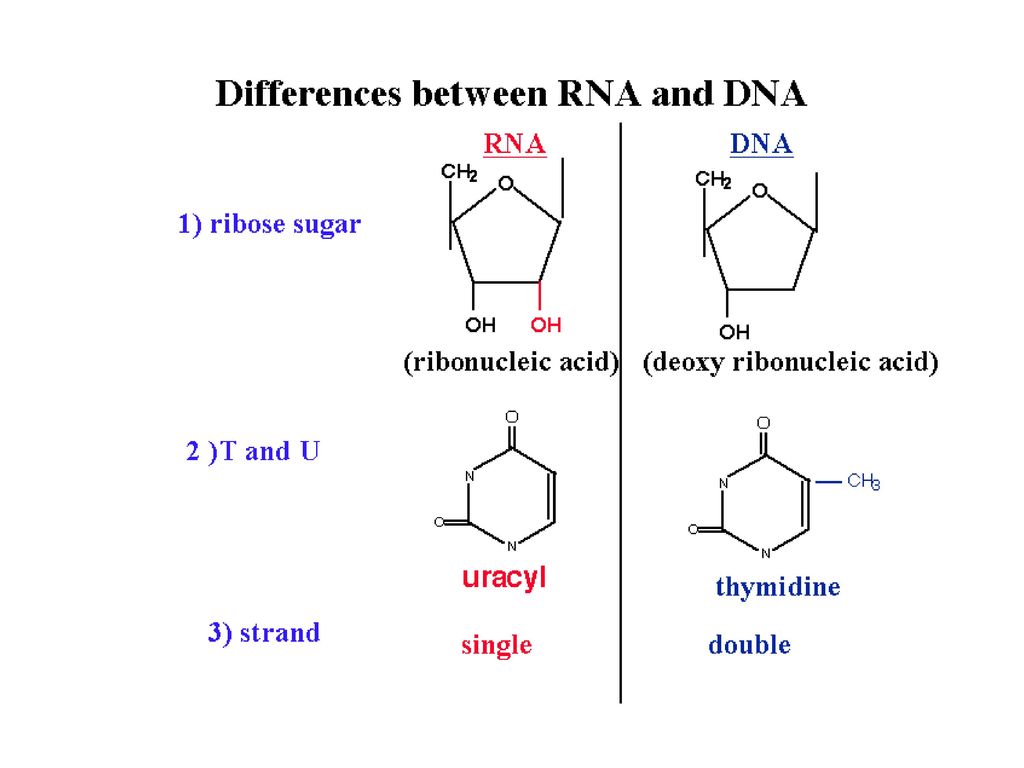
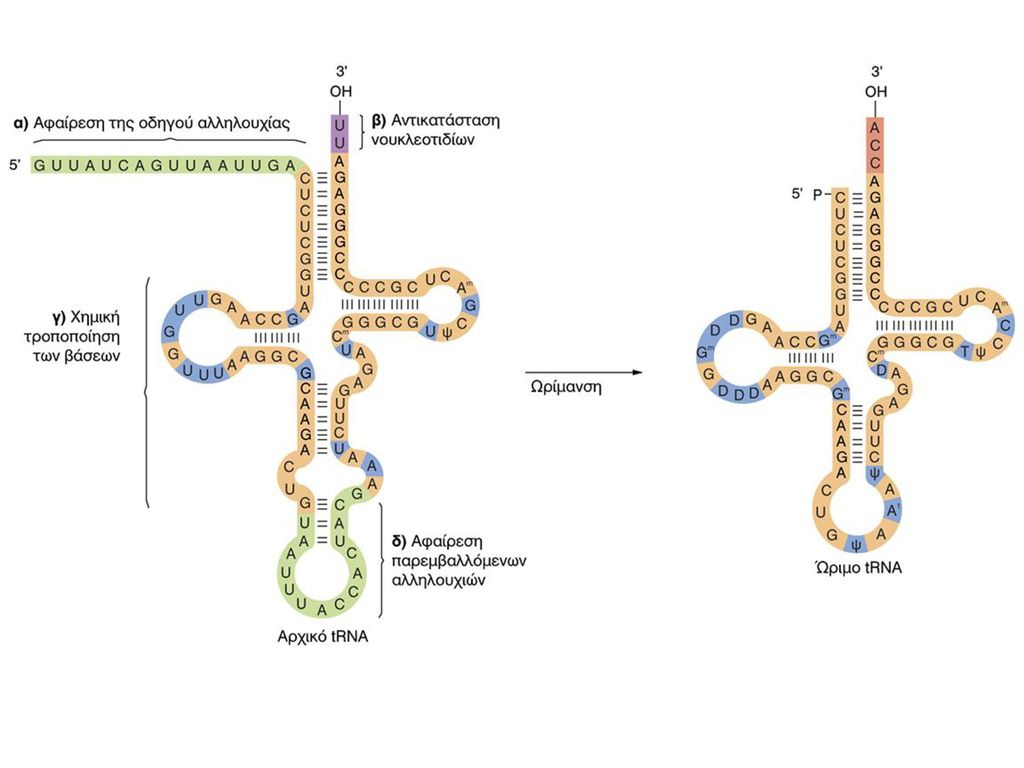
ΔΟΜΙΚΕΣ ΜΟΝΑΔΕΣ RNA

10
ΖΕΥΓΑΡΩΜΑ ΒΑΣΕΩΝ RNA-DNA

11
RNA ΠΟΛΥΜΕΡΑΣΗ Μήτρα DNA- sense strand Πολυμερισμός ριβονουκλεοτιδίων
Πολυμερισμός 5΄ 3΄ Ζεύγη βάσεων RNA-DNA Δεν απαιτείται εναρκτήριο RNA ΌΧΙ επιδιόρθωση λαθών Απαιτούνται βοηθητικοί παράγοντες μεταγραφής: Γενικοί και Ειδικοί

12
RNA ΠΟΛΥΜΕΡΑΣΕΣ ΕΥΚΑΡΥΩΤΙΚΩΝ RNA ΠΟΛΥΜΕΡΑΣΕΣ ΠΡΟΚΑΡΥΩΤΙΚΩΝ
RNA-πολυμεράση Ι rRNA RNA-πολυμεράση ΙI mRNA, snRNAs RNA-πολυμεράση ΙII tRNA, snRNAs RNA ΠΟΛΥΜΕΡΑΣΕΣ ΠΡΟΚΑΡΥΩΤΙΚΩΝ RNA-πολυμεράση Ριφαμπικίνη

13
ΕΠΙΜΗΚΥΝΣΗ & ΤΕΡΜΑΤΙΣΜΟΣ
ΕΛΕΓΧΟΣ ΜΕΤΑΓΡΑΦΗΣ ΕΠΙΜΗΚΥΝΣΗ & ΤΕΡΜΑΤΙΣΜΟΣ

14
ΕΛΕΓΧΟΣ ΜΕΤΑΓΡΑΦΗΣ ΥΠΟΚΙΝΗΤΗΣ
Αριστερά (5΄) από την έναρξη της μεταγραφής
 από την έναρξη της μεταγραφής.")
16
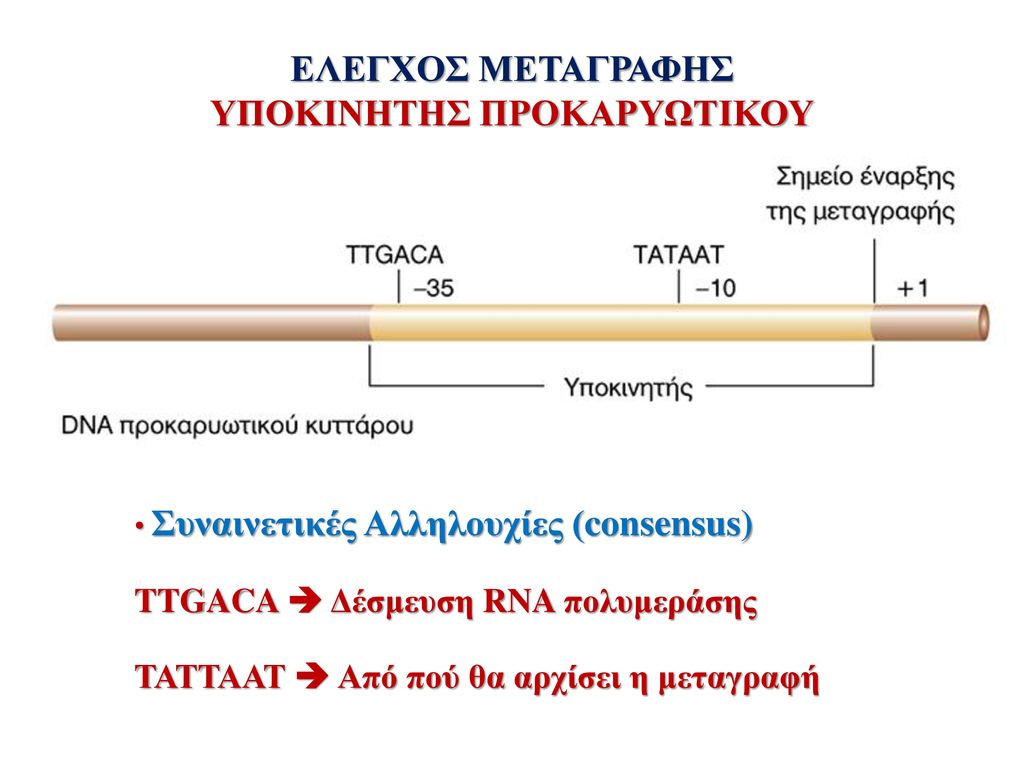
ΥΠΟΚΙΝΗΤΗΣ ΠΡΟΚΑΡΥΩΤΙΚΟΥ
ΕΛΕΓΧΟΣ ΜΕΤΑΓΡΑΦΗΣ ΥΠΟΚΙΝΗΤΗΣ ΠΡΟΚΑΡΥΩΤΙΚΟΥ Συναινετικές Αλληλουχίες (consensus) TTGACA Δέσμευση RNA πολυμεράσης ΤΑΤΤΑΑΤ Από πού θα αρχίσει η μεταγραφή
 TTGACA Δέσμευση RNA πολυμεράσης. ΤΑΤΤΑΑΤ Από πού θα αρχίσει η μεταγραφή.")
17
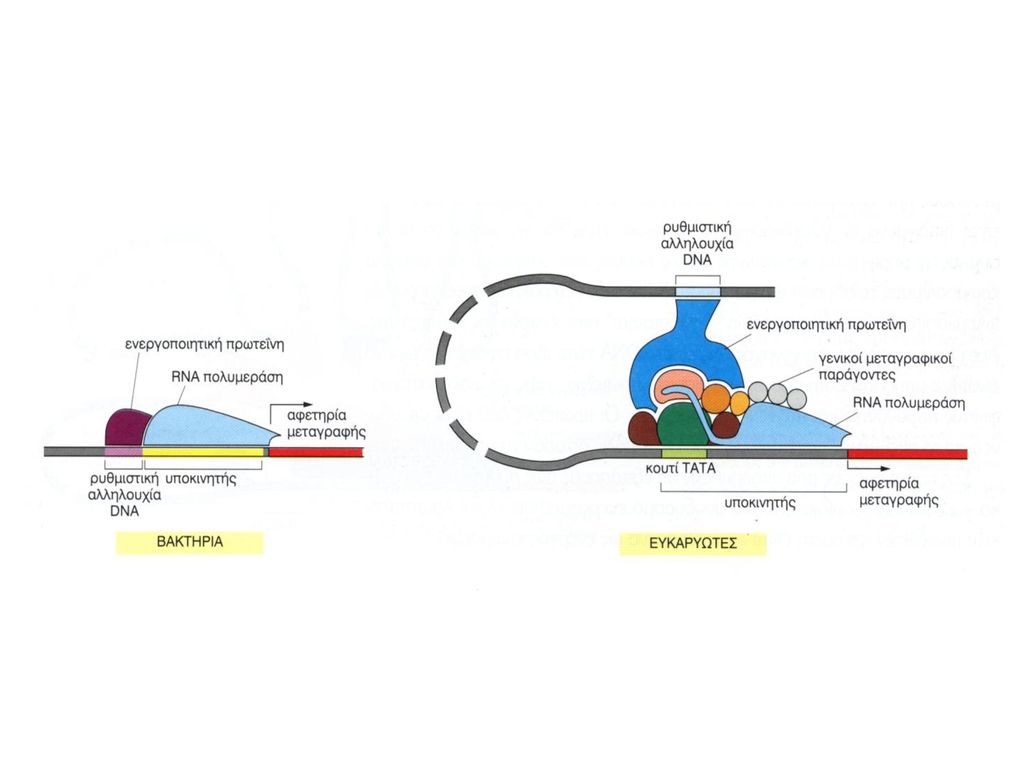
ΥΠΟΚΙΝΗΤΗΣ ΕΥΚΑΡΥΩΤΙΚΟΥ
ΕΛΕΓΧΟΣ ΜΕΤΑΓΡΑΦΗΣ ΥΠΟΚΙΝΗΤΗΣ ΕΥΚΑΡΥΩΤΙΚΟΥ Συναινετικές Αλληλουχίες (consensus) TΑΤΑ Δέσμευση Γενικών Μεταγραφικών Παραγόντων CAAT & GC Δέσμευση Ειδικών Μεταγραφικών Παραγόντων Ρύθμιση Αποδοτικότητας Μεταγραφής
 TΑΤΑ Δέσμευση Γενικών Μεταγραφικών Παραγόντων. CAAT & GC Δέσμευση Ειδικών Μεταγραφικών Παραγόντων Ρύθμιση Αποδοτικότητας Μεταγραφής.")
18
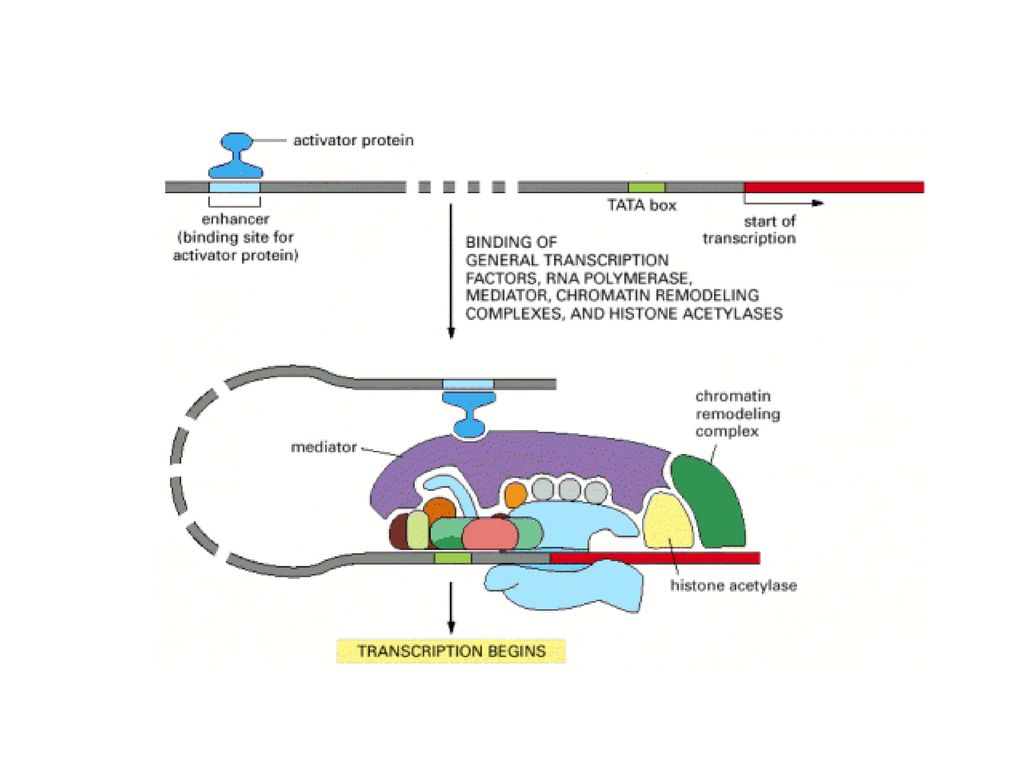
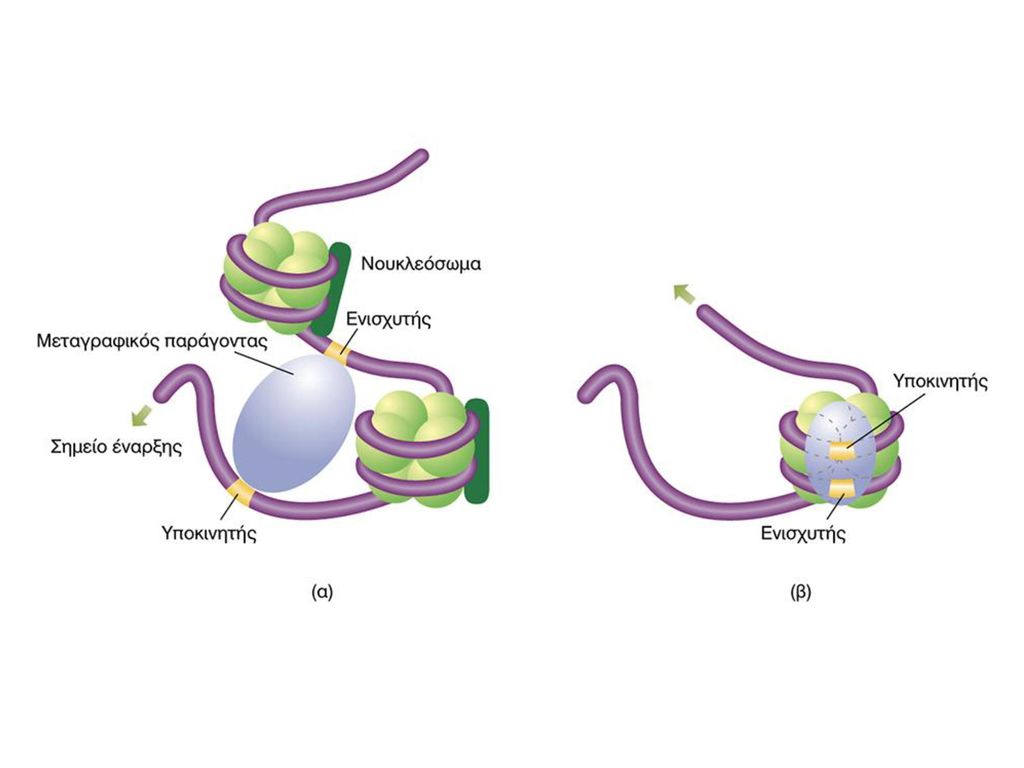
ΕΝΙΣΧΥΤΕΣ & ΚΑΤΑΣΤΟΛΕΙΣ
ΕΛΕΓΧΟΣ ΜΕΤΑΓΡΑΦΗΣ ΕΝΙΣΧΥΤΕΣ & ΚΑΤΑΣΤΟΛΕΙΣ Figure 1 Comparison of a simple eukaryotic promoter and extensively diversified metazoan regulatory modules. a, Simple eukaryotic transcriptional unit. A simple core promoter (TATA), upstream activator sequence (UAS) and silencer element spaced within 100–200 bp of the TATA box that is typically found in unicellular eukaryotes. b, Complex metazoan transcriptional control modules. A complex arrangement of multiple clustered enhancer modules interspersed with silencer and insulator elements which can be located 10–50 kb either upstream or downstream of a composite core promoter containing TATA box (TATA), Initiator sequences (INR), and downstream promoter elements (DPE). cis-δραστικές αλληλουχίες DNA Δέσμευση Ρυθμιστικών Μεταγραφικών Παραγόντων Όχι συγκεκριμένη θέση-Μακριά από έναρξη μεταγραφής
, upstream activator sequence (UAS) and silencer element spaced within 100–200 bp of the TATA box that is typically found in unicellular eukaryotes. b, Complex metazoan transcriptional control modules. A complex arrangement of multiple clustered enhancer modules interspersed with silencer and insulator elements which can be located 10–50 kb either upstream or downstream of a composite core promoter containing TATA box (TATA), Initiator sequences (INR), and downstream promoter elements (DPE). cis-δραστικές αλληλουχίες DNA. Δέσμευση Ρυθμιστικών Μεταγραφικών Παραγόντων. Όχι συγκεκριμένη θέση-Μακριά από έναρξη μεταγραφής.")
24
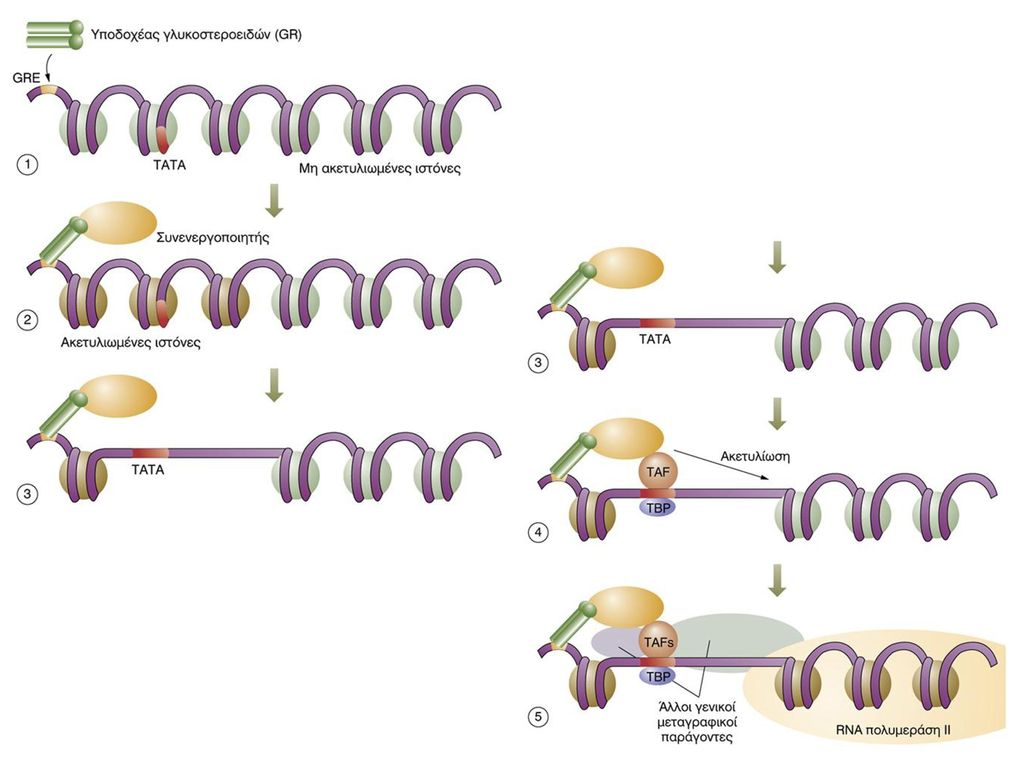
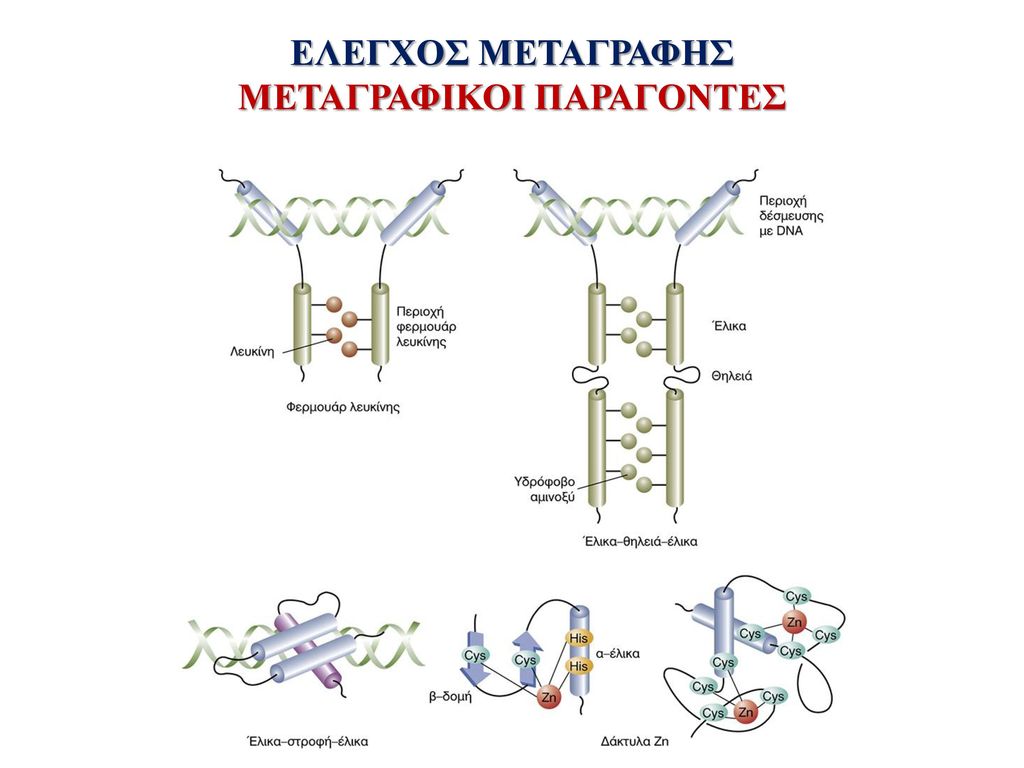
ΜΕΤΑΓΡΑΦΙΚΟΙ ΠΑΡΑΓΟΝΤΕΣ
ΕΛΕΓΧΟΣ ΜΕΤΑΓΡΑΦΗΣ ΜΕΤΑΓΡΑΦΙΚΟΙ ΠΑΡΑΓΟΝΤΕΣ

25
ΕΛΕΓΧΟΣ ΜΕΤΑΓΡΑΦΗΣ Trans-Ρύθμιση

27
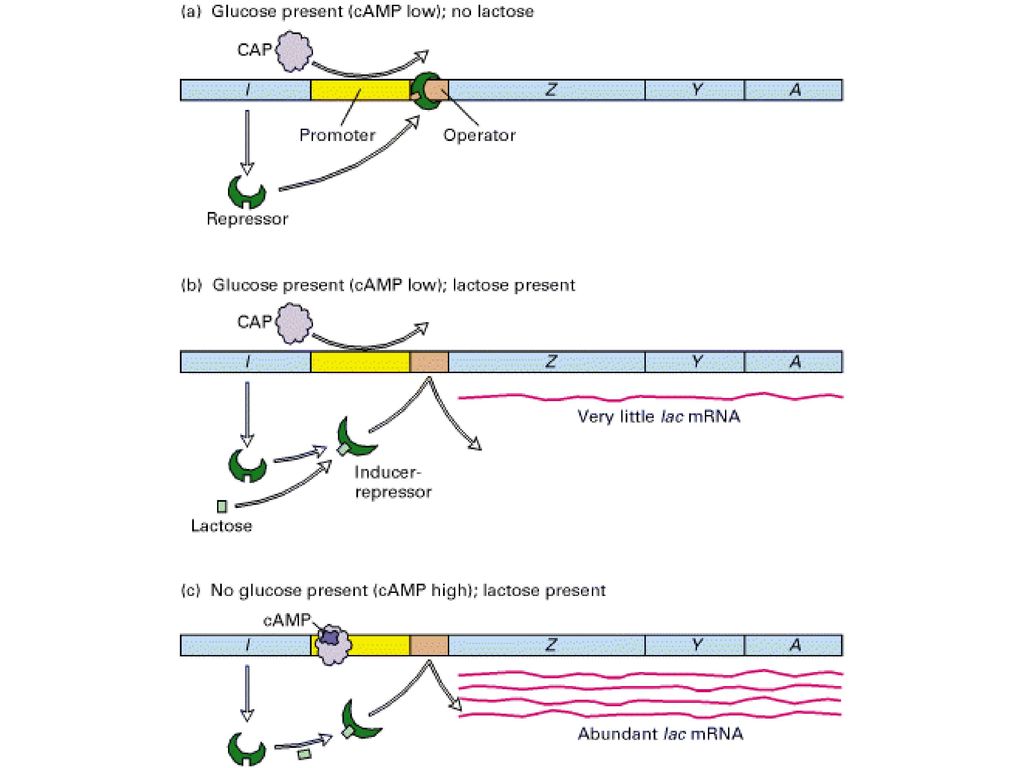
Figure Negative and positive transcriptional control of the lac operon by the lac repressor and cAMP-CAP, respectively. (a) In the absence of lactose, no lac mRNA is formed because repressor bound to the lac operator prevents transcription. (b) In the presence of glucose and lactose, the lac repressor binds lactose and undergoes a conformational change, so that it does not bind to the lac operator. However, cAMP is low, because glucose is present, and thus cAMP-CAP does not bind to the CAP site in the operator. As a result, RNA polymerase does not bind efficiently to the lac promoter and only a little lac mRNA is synthesized. (c) In the presence of lactose and the absence of glucose, maximal transcription of the lac operon occurs. In this situation, the lac repressor does not bind to the lac operator, the concentration of cAMP increases, and the cAMP-CAP complex that forms binds at the CAP site, stimulating binding and initiation by RNA polymerase.
 In the absence of lactose, no lac mRNA is formed because repressor bound to the lac operator prevents transcription. (b) In the presence of glucose and lactose, the lac repressor binds lactose and undergoes a conformational change, so that it does not bind to the lac operator. However, cAMP is low, because glucose is present, and thus cAMP-CAP does not bind to the CAP site in the operator. As a result, RNA polymerase does not bind efficiently to the lac promoter and only a little lac mRNA is synthesized. (c) In the presence of lactose and the absence of glucose, maximal transcription of the lac operon occurs. In this situation, the lac repressor does not bind to the lac operator, the concentration of cAMP increases, and the cAMP-CAP complex that forms binds at the CAP site, stimulating binding and initiation by RNA polymerase..")
29
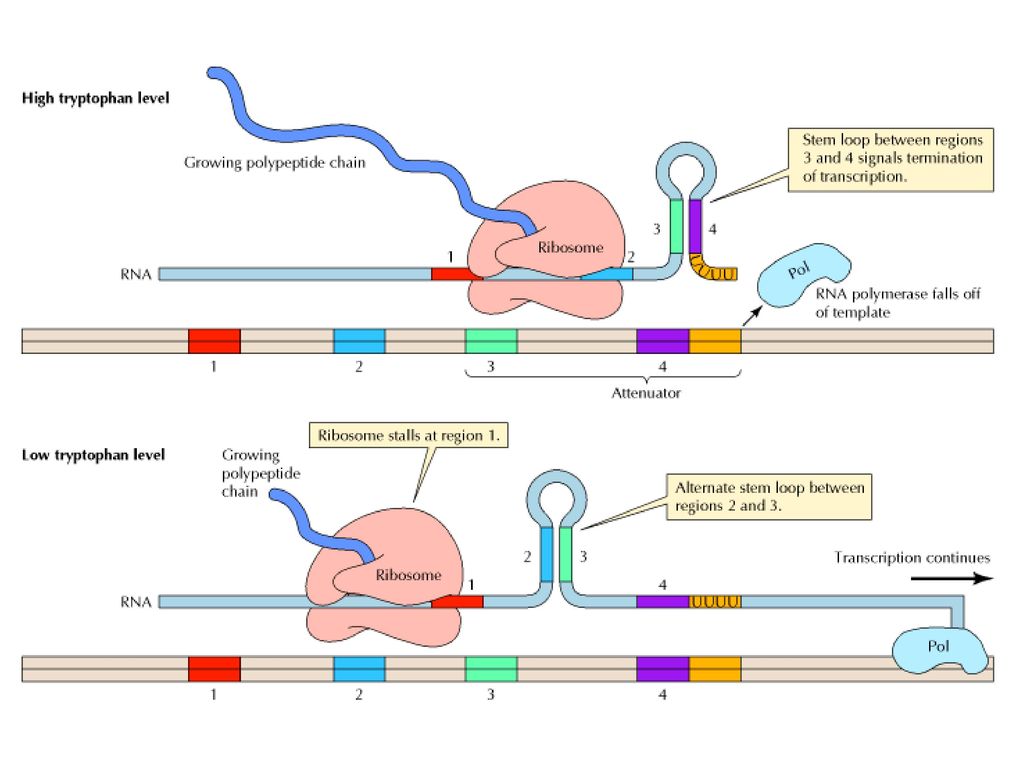
Figure Mechanism of transcriptional attenuation The trp mRNA is translated while still being synthesized. In the presence of high levels of tryptophan, the ribosomes proceed along the message slightly behind the site of transcription. Under these conditions, the mRNA regions designated 3 and 4 hybridize to form a stem-loop structure that signals the termination of transcription. In the presence of low levels of tryptophan, however, the ribosomes stall at region 1 of the mRNA, which contains two adjacent codons for tryptophan. In this case, since region 2 is not bound to a ribosome, it is free to form an alternative stem-loop structure by hybridizing to region 3. This hybridization prevents formation of the 3 4 stem loop, and transcription is able to continue past the attenuator sequence. (Albert´s)
.")
30
Genomic imprinting is an epigenetic mechanism of transcriptional regulation through which expression of a subset of mammalian genes is restricted to one parental allele

31
Σύνδρομο Prader-Willi

32
M Π 15q11-13

36
7-13

37
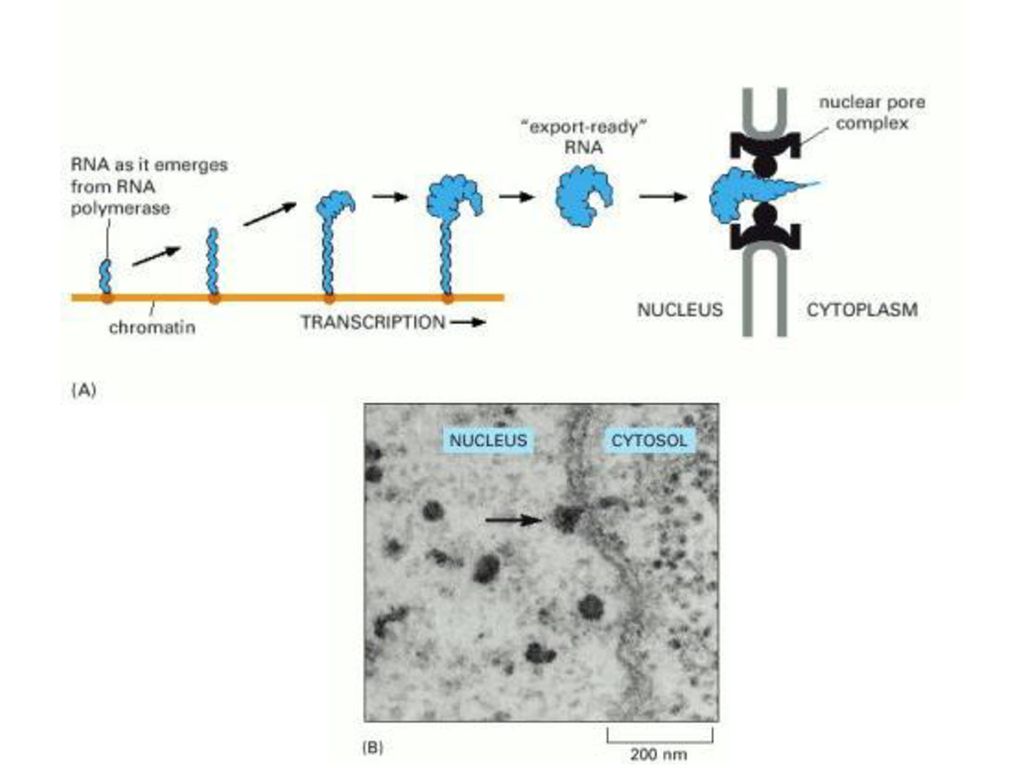
ΠΡΟΣΘΗΚΗ 5’ ΚΑΛΥΜΜΑΤΟΣ

38
ΠΡΟΣΘΗΚΗ ΠΟΛΥ-A ΟΥΡΑΣ Figure Some of the major steps in generating the 3 end of a eucaryotic mRNA. This process is much more complicated than the analogous process in bacteria, where the RNA polymerase simply stops at a termination signal and releases both the 3 end of its transcript and the DNA template. CstF=cleavage stimulation factor © 2002 by Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walter.

39
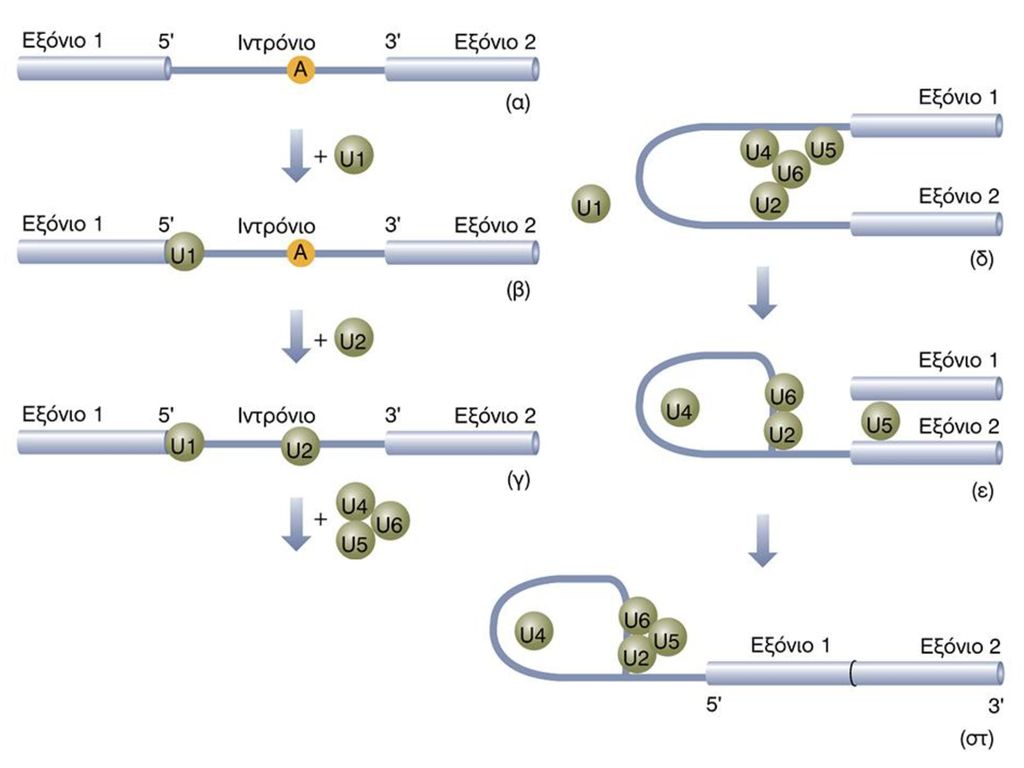
SPLICING / ΜΑΤΙΣΜΑ Figure Splicing mechanisms in group I and group II self-splicing introns and spliceosome-catalyzed splicing of pre-mRNA. The intron is shown in blue; the exons to be joined in red. In group I introns, a guanosine cofactor (G) that is not part of the RNA chain associates with the active site. The 3 -hydroxyl group of this guanosine participates in a transesterification reaction with the phosphate at the 5 end of the intron; this reaction is analogous to that involving the 2 -hydroxyl groups of the branch-site A in group II introns and pre-mRNA introns spliced in spliceosomes (see Figure 11-16). The subsequent transesterification that links the 5 and 3 exons is similar in all three splicing mechanisms. Note that spliced-out group I introns are linear structures, unlike the branched intron products in the other two cases. [Adapted from P. A. Sharp, 1987, Science 235:769.] (Molecular Cell Biology, Lodish et al.)
 that is not part of the RNA chain associates with the active site. The 3 -hydroxyl group of this guanosine participates in a transesterification reaction with the phosphate at the 5 end of the intron; this reaction is analogous to that involving the 2 -hydroxyl groups of the branch-site A in group II introns and pre-mRNA introns spliced in spliceosomes (see Figure 11-16). The subsequent transesterification that links the 5 and 3 exons is similar in all three splicing mechanisms. Note that spliced-out group I introns are linear structures, unlike the branched intron products in the other two cases. [Adapted from P. A. Sharp, 1987, Science 235:769.] (Molecular Cell Biology, Lodish et al.)")
41
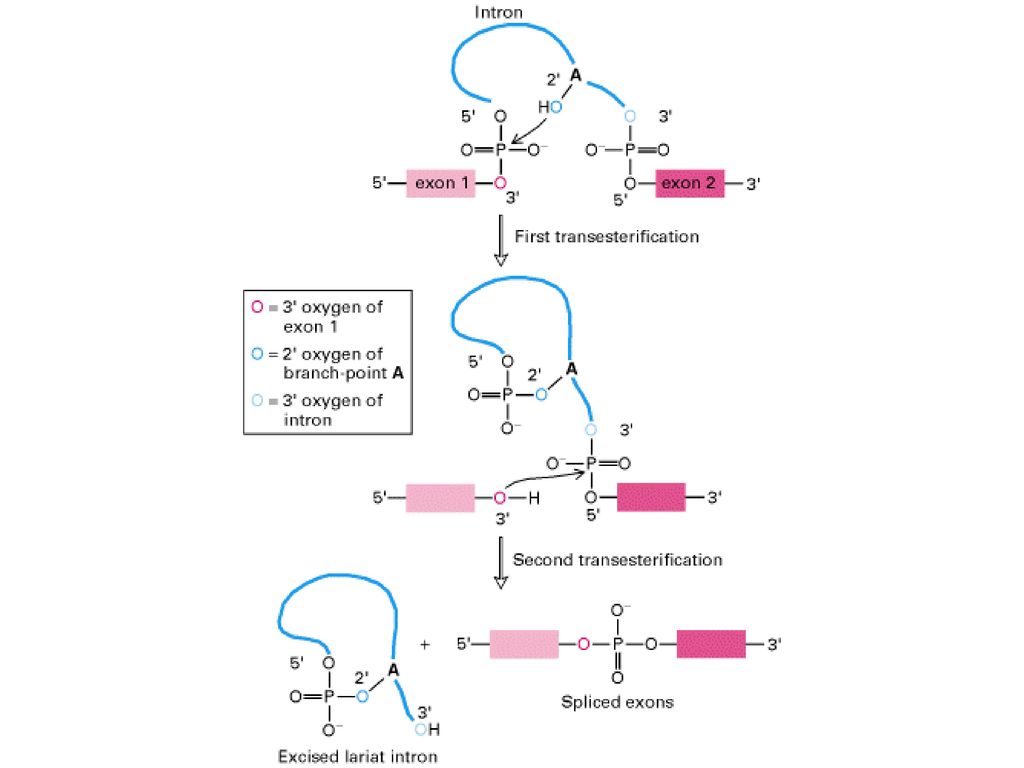
Figure Splicing of exons in pre-mRNA occurs via two transesterification reactions. In the first reaction, the ester bond between the 5 phosphorus of the intron and the 3 oxygen (red) of exon 1 is exchanged for an ester bond with the 2 oxygen (dark blue) of the branch-site A residue. In the second reaction, the ester bond between the 5 phosphorus of exon 2 and the 3 oxygen (light blue) of the intron is exchanged for an ester bond with the 3 oxygen of exon 1, releasing the intron as a lariat structure and joining the two exons. Arrows show where the activated hydroxyl oxygens react with phosphorus atoms. (Molecular Cell Biology, Lodish et al.)
 of exon 1 is exchanged for an ester bond with the 2 oxygen (dark blue) of the branch-site A residue. In the second reaction, the ester bond between the 5 phosphorus of exon 2 and the 3 oxygen (light blue) of the intron is exchanged for an ester bond with the 3 oxygen of exon 1, releasing the intron as a lariat structure and joining the two exons. Arrows show where the activated hydroxyl oxygens react with phosphorus atoms. (Molecular Cell Biology, Lodish et al.).")
44
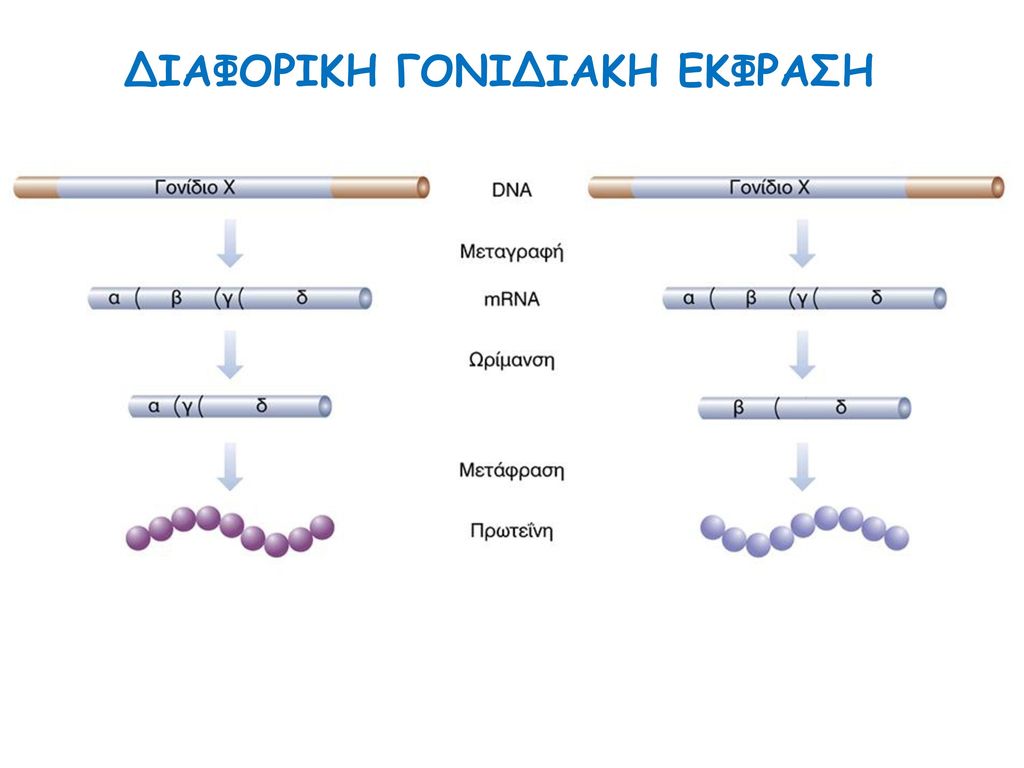
ΔΙΑΦΟΡΙΚΗ ΓΟΝΙΔΙΑΚΗ ΕΚΦΡΑΣΗ

45
ΕΝΑΛΛΑΚΤΙΚΟ SPLICING

46
EDITING Figure RNA editing of apo-B pre-mRNA. The apo-B mRNA produced in the liver has the same sequence as the exons in the primary transcript. This mRNA is translated into Apo-B100, which has two functional domains: a N-terminal domain (green) that associates with lipids and a C-terminal domain (orange) that binds to LDL receptors on cell membranes. In the apo-B mRNA produced in the intestine, the CAA codon in exon 26 is edited to a UAA stop codon. As a result, intestinal cells produce Apo-B48, which corresponds to the N-terminal domain of Apo-B100. [Adapted from P. Hodges and J. Scott, 1992, Trends Biochem. Sci. 17:77 . (Molecular Cell Biology, Lodish et al.)
 that associates with lipids and a C-terminal domain (orange) that binds to LDL receptors on cell membranes. In the apo-B mRNA produced in the intestine, the CAA codon in exon 26 is edited to a UAA stop codon. As a result, intestinal cells produce Apo-B48, which corresponds to the N-terminal domain of Apo-B100. [Adapted from P. Hodges and J. Scott, 1992, Trends Biochem. Sci. 17:77 . (Molecular Cell Biology, Lodish et al.)")
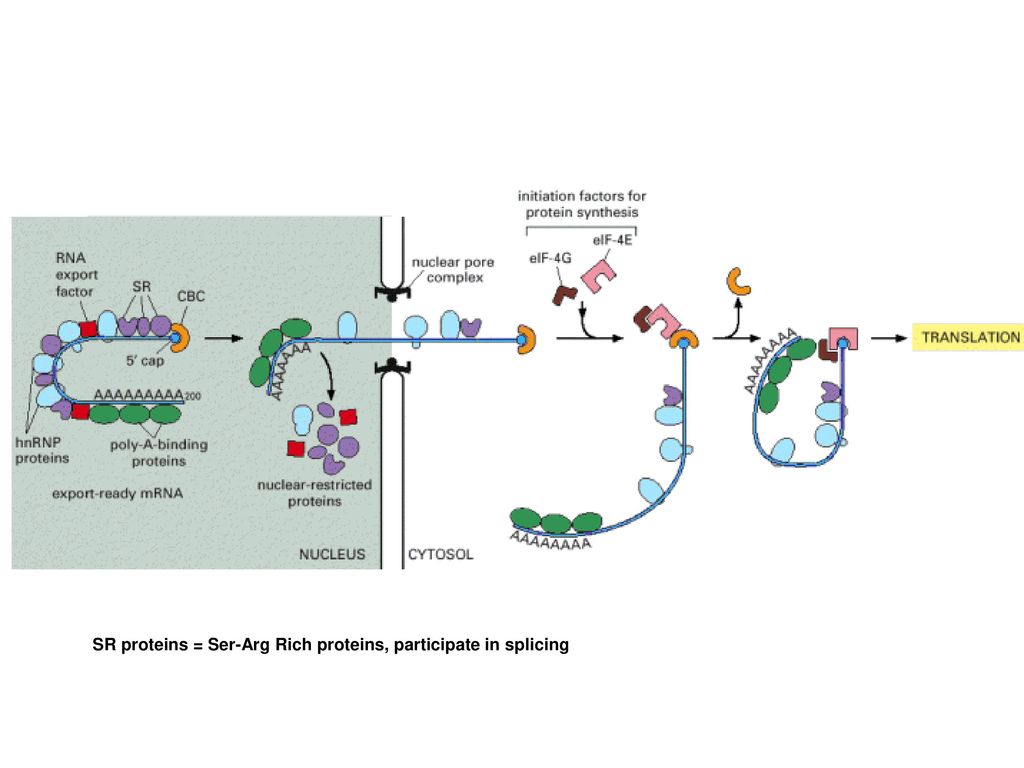
51
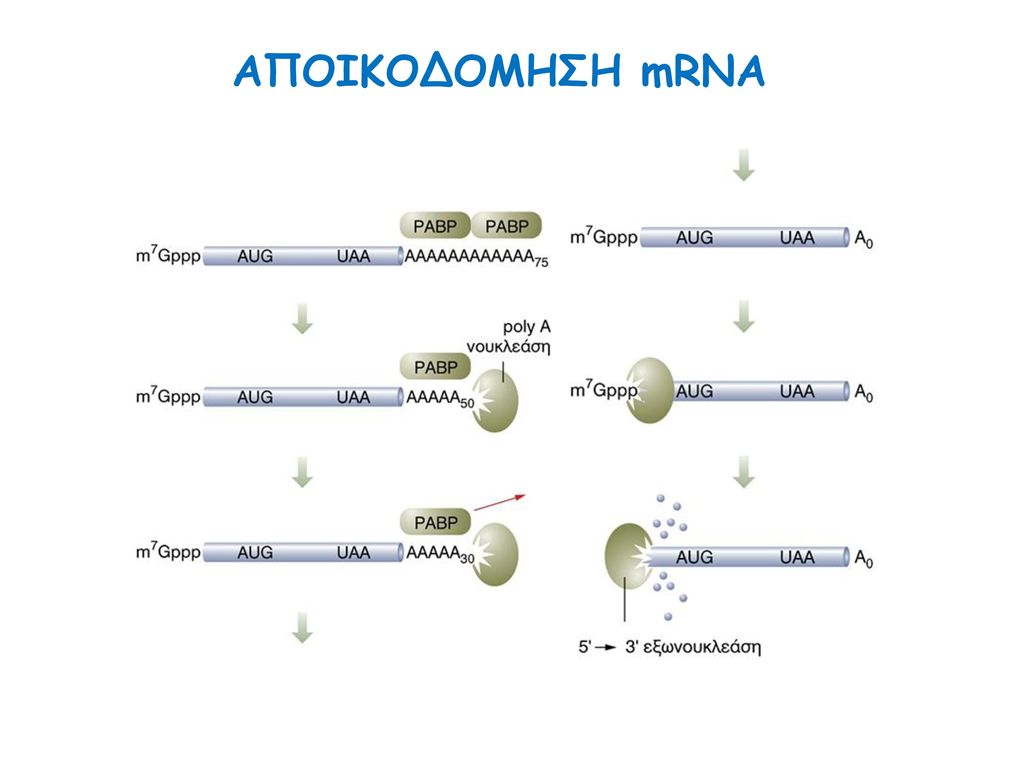
ΑΠΟΙΚΟΔΟΜΗΣΗ mRNA

55
CBC=cap binding complex
SR proteins = Ser-Arg Rich proteins, participate in splicing

56
MicroRNA (miRNA) are small, non-coding RNAs composed of nucleotides. To date, 678 human miRNAs have been identified, and it is estimated that the human genome encodes approximately 1000 miRNAs, leaving at least 300 miRNAs left for discovery[1,2]. MiRNAs are major players in the regulation of genes that are responsible for a myriad of cellular functions including apoptosis, differentiation, development, cellular proliferation, metabolism, and adhesion. In fact, it is estimated that up to 30% of genes in humans are regulated by miRNA, making these molecules one of the largest families of genomic regulators [3, 4]. It has been found that one miRNA can bind up to 200 gene targets, exhibiting the significant duties of just one of these small RNAs. These targets can be diverse in their function, and include transcription factors, secreted factor, receptors, and transporters [4]. At the same time, multiple miRNAs can regulate translation of the same protein, demonstrating the redundancy seen in the functions of these molecules.
![MicroRNA (miRNA) are small, non-coding RNAs composed of nucleotides. To date, 678 human miRNAs have been identified, and it is estimated that the human genome encodes approximately 1000 miRNAs, leaving at least 300 miRNAs left for discovery[1,2]. MiRNAs are major players in the regulation of genes that are responsible for a myriad of cellular functions including apoptosis, differentiation, development, cellular proliferation, metabolism, and adhesion. In fact, it is estimated that up to 30% of genes in humans are regulated by miRNA, making these molecules one of the largest families of genomic regulators [3, 4]. It has been found that one miRNA can bind up to 200 gene targets, exhibiting the significant duties of just one of these small RNAs.](http://slideplayer.gr/slide/11550345/62/images/56/MicroRNA+%28miRNA%29+are+small%2C+non-coding+RNAs+composed+of+nucleotides.+To+date%2C+678+human+miRNAs+have+been+identified%2C+and+it+is+estimated+that+the+human+genome+encodes+approximately+1000+miRNAs%2C+leaving+at+least+300+miRNAs+left+for+discovery%5B1%2C2%5D.+MiRNAs+are+major+players+in+the+regulation+of+genes+that+are+responsible+for+a+myriad+of+cellular+functions+including+apoptosis%2C+differentiation%2C+development%2C+cellular+proliferation%2C+metabolism%2C+and+adhesion.+In+fact%2C+it+is+estimated+that+up+to+30%25+of+genes+in+humans+are+regulated+by+miRNA%2C+making+these+molecules+one+of+the+largest+families+of+genomic+regulators+%5B3%2C+4%5D.+It+has+been+found+that+one+miRNA+can+bind+up+to+200+gene+targets%2C+exhibiting+the+significant+duties+of+just+one+of+these+small+RNAs..jpg "These targets can be diverse in their function, and include transcription factors, secreted factor, receptors, and transporters [4]. At the same time, multiple miRNAs can regulate translation of the same protein, demonstrating the redundancy seen in the functions of these molecules. .")
57
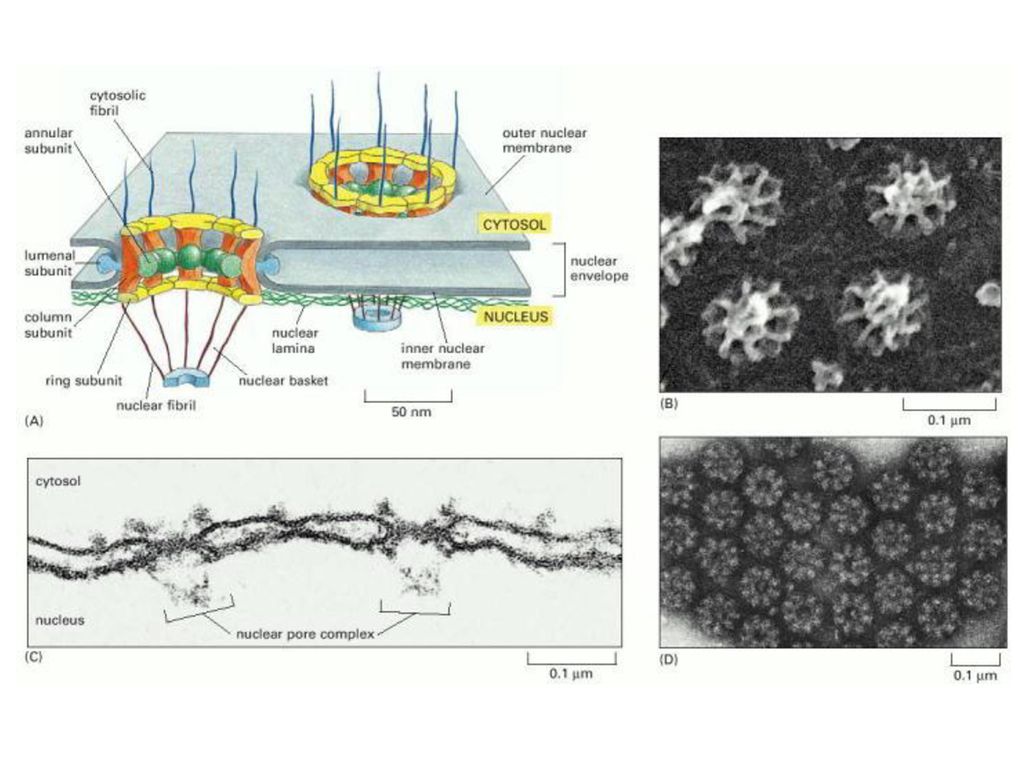
The genes for miRNAs are often found in the intronic regions of genes coding for mRNAs, or they are transcribed as independent units [6]. Transcription of miRNA genes occurs primarily via RNA polymerase II, however, scientists recently uncovered several miRNAs transcribed by RNA pol III. Upon transcription the long primary miRNA (pri-miRNA) is capped with a 7-methylguanosine cap, and given a poly-A tail. It is at this point that the miRNA has a foldback structure consisting of a stem loop with two flanking segments. The foldback structure is further processed within the nucleus during a step known as cropping, by the RNAse III enzyme Drosher, and a double strand RNA-binding domain (dsRBD) protein named DGCR8, to become a nucleotide precursor-miRNA (pre-miRNA). The next step in the maturation of miRNA involves the export of the pre-miRNA from the nucleus to the cytoplasm. This function is performed by exportin-5 (Exp-5), which happens to be dependent on a Ran-GTP acting as a cofactor. The Exp-5 facilitates the movement of the pre-miRNA through a nuclear pore. Once in the cytoplasm, the stem loop of the pre-miRNA is cleaved, and a double stranded RNA duplex of approximately nucleotides in length remains. This cleavage is performed by the RNAse III enzyme Dicer. The fate of the two strands of RNA composing the duplex has been recently debated. Some authors indicate that the cell is capable of using only one of the pieces of RNA within the duplex, while others assert that both can be used in separate roles. Nonetheless, one strand of RNA known as the guide strand of miRNA is loaded onto a complex of proteins called the RNA-induced silencing complex (RISC) to form the miRISC[3]. The miRISC is the effector complex and mediates miRNA function.
![The genes for miRNAs are often found in the intronic regions of genes coding for mRNAs, or they are transcribed as independent units [6]. Transcription of miRNA genes occurs primarily via RNA polymerase II, however, scientists recently uncovered several miRNAs transcribed by RNA pol III.](http://slideplayer.gr/slide/11550345/62/images/57/The+genes+for+miRNAs+are+often+found+in+the+intronic+regions+of+genes+coding+for+mRNAs%2C+or+they+are+transcribed+as+independent+units+%5B6%5D.+Transcription+of+miRNA+genes+occurs+primarily+via+RNA+polymerase+II%2C+however%2C+scientists+recently+uncovered+several+miRNAs+transcribed+by+RNA+pol+III..jpg "Upon transcription the long primary miRNA (pri-miRNA) is capped with a 7-methylguanosine cap, and given a poly-A tail. It is at this point that the miRNA has a foldback structure consisting of a stem loop with two flanking segments. The foldback structure is further processed within the nucleus during a step known as cropping, by the RNAse III enzyme Drosher, and a double strand RNA-binding domain (dsRBD) protein named DGCR8, to become a nucleotide precursor-miRNA (pre-miRNA). The next step in the maturation of miRNA involves the export of the pre-miRNA from the nucleus to the cytoplasm. This function is performed by exportin-5 (Exp-5), which happens to be dependent on a Ran-GTP acting as a cofactor. The Exp-5 facilitates the movement of the pre-miRNA through a nuclear pore. Once in the cytoplasm, the stem loop of the pre-miRNA is cleaved, and a double stranded RNA duplex of approximately nucleotides in length remains. This cleavage is performed by the RNAse III enzyme Dicer. The fate of the two strands of RNA composing the duplex has been recently debated. Some authors indicate that the cell is capable of using only one of the pieces of RNA within the duplex, while others assert that both can be used in separate roles. Nonetheless, one strand of RNA known as the guide strand of miRNA is loaded onto a complex of proteins called the RNA-induced silencing complex (RISC) to form the miRISC[3]. The miRISC is the effector complex and mediates miRNA function. .")
58
Επεξεργασία των μικρο-RNAs
Overview of miRNA processing. MicroRNAs (miRNAs) comprise a growing class of endogenous molecules that regulate gene expression post-transcriptionally. By binding to partially complementary regions at the 3' end of messenger RNAs, these approximately 22-nucleotide single-stranded molecules direct target site recognition by the macromolecular RNA-induced silencing complex (RISC). RISC induces cleavage or translational repression of targeted transcripts. Mammalian miRNA biosynthesis begins with RNA polymerase II-dependent transcription, which yields long primary miRNA (pri-miRNA) transcripts. The pri-miRNA is processed in the nucleus to a pre-miRNA by the Microprocesser complex, which consists of Drosha, a member of ribonuclease III family (RNaseIII), and DGCR8 (also known as Pasha), which contains two double-stranded RNA binding domains and is responsible for pri-miRNA recognition. The pre-miRNA is then exported to the cytoplasm by Exportin-5 and further processed into a miRNA duplex by Dicer, another RNase III. One strand of the miRNA duplex is then incorporated into the RISC.
 comprise a growing class of endogenous molecules that regulate gene expression post-transcriptionally. By binding to partially complementary regions at the 3 end of messenger RNAs, these approximately 22-nucleotide single-stranded molecules direct target site recognition by the macromolecular RNA-induced silencing complex (RISC). RISC induces cleavage or translational repression of targeted transcripts. Mammalian miRNA biosynthesis begins with RNA polymerase II-dependent transcription, which yields long primary miRNA (pri-miRNA) transcripts. The pri-miRNA is processed in the nucleus to a pre-miRNA by the Microprocesser complex, which consists of Drosha, a member of ribonuclease III family (RNaseIII), and DGCR8 (also known as Pasha), which contains two double-stranded RNA binding domains and is responsible for pri-miRNA recognition. The pre-miRNA is then exported to the cytoplasm by Exportin-5 and further processed into a miRNA duplex by Dicer, another RNase III. One strand of the miRNA duplex is then incorporated into the RISC.")
59
Ποια είναι τα χαρακτηριστικά της RNA πολυμεράσης;
Τι είναι ο υποκινητής και τι ο ενισχυτής ενός γονιδίου; Πώς η συσπείρωση της χρωματίνης επηρεάζει τη γονιδιακή έκφραση; Πώς ρυθμίζονται τα οπερόνια της λακτόζης ή της τρυπτοφάνης; Ποιες είναι οι μετα-μεταγραφικές τροποποιήσεις του mRNA; Τι είναι το εναλλακτικό splicing; Τι είναι τα microRNAs;

Παρόμοιες παρουσιάσεις














 Όραση Μαρία Κουτρομάνου. Structure of the Eye: Iris The iris is similar to the diaphragm in a camera Your iris widens in dim light and.>")





